Causality of Biodiversity Loss: Climate, Vegetation, and Urbanization in China and America

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Preprocessing
2.2. Analyses
3. Results and Discussion
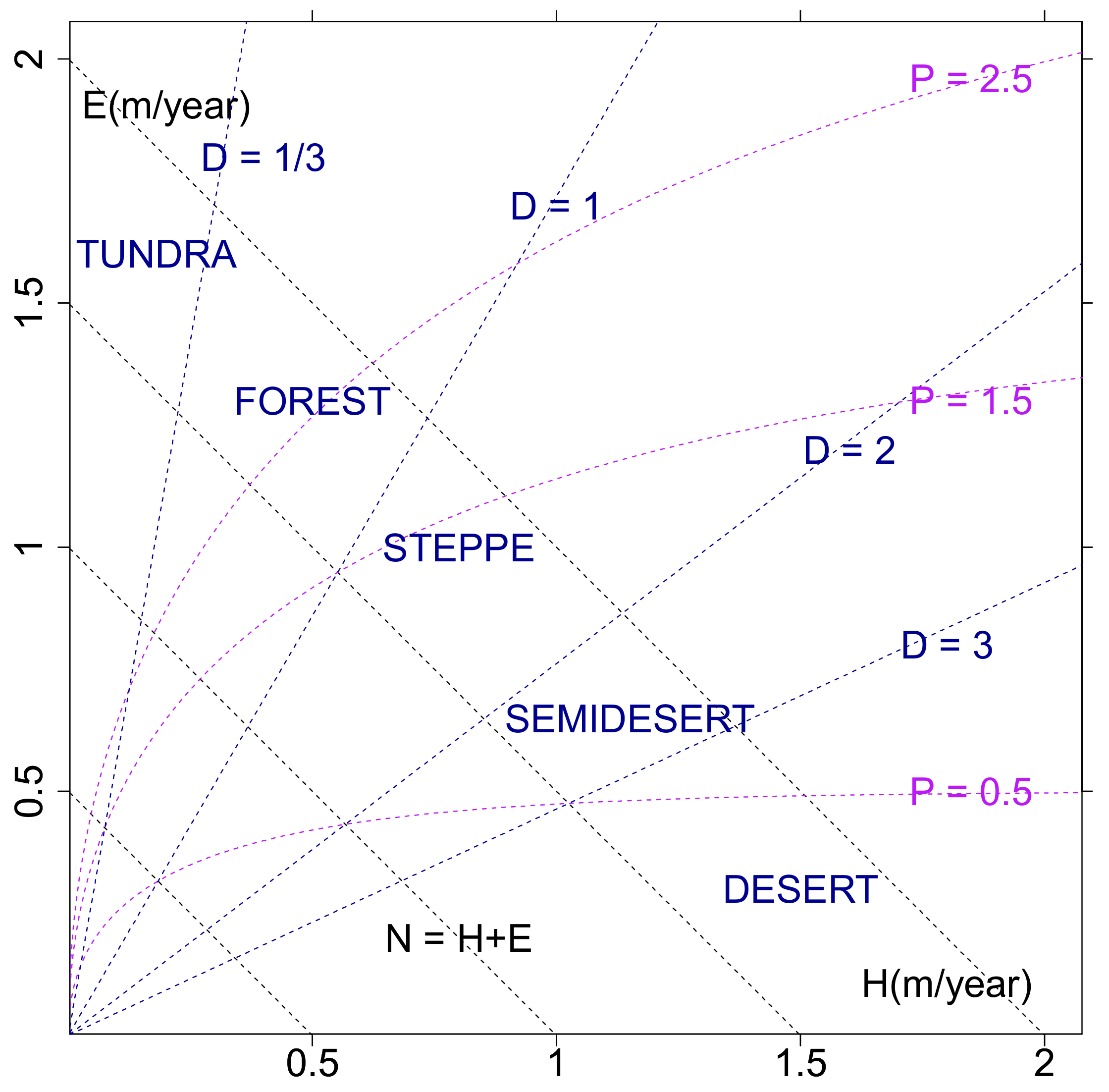
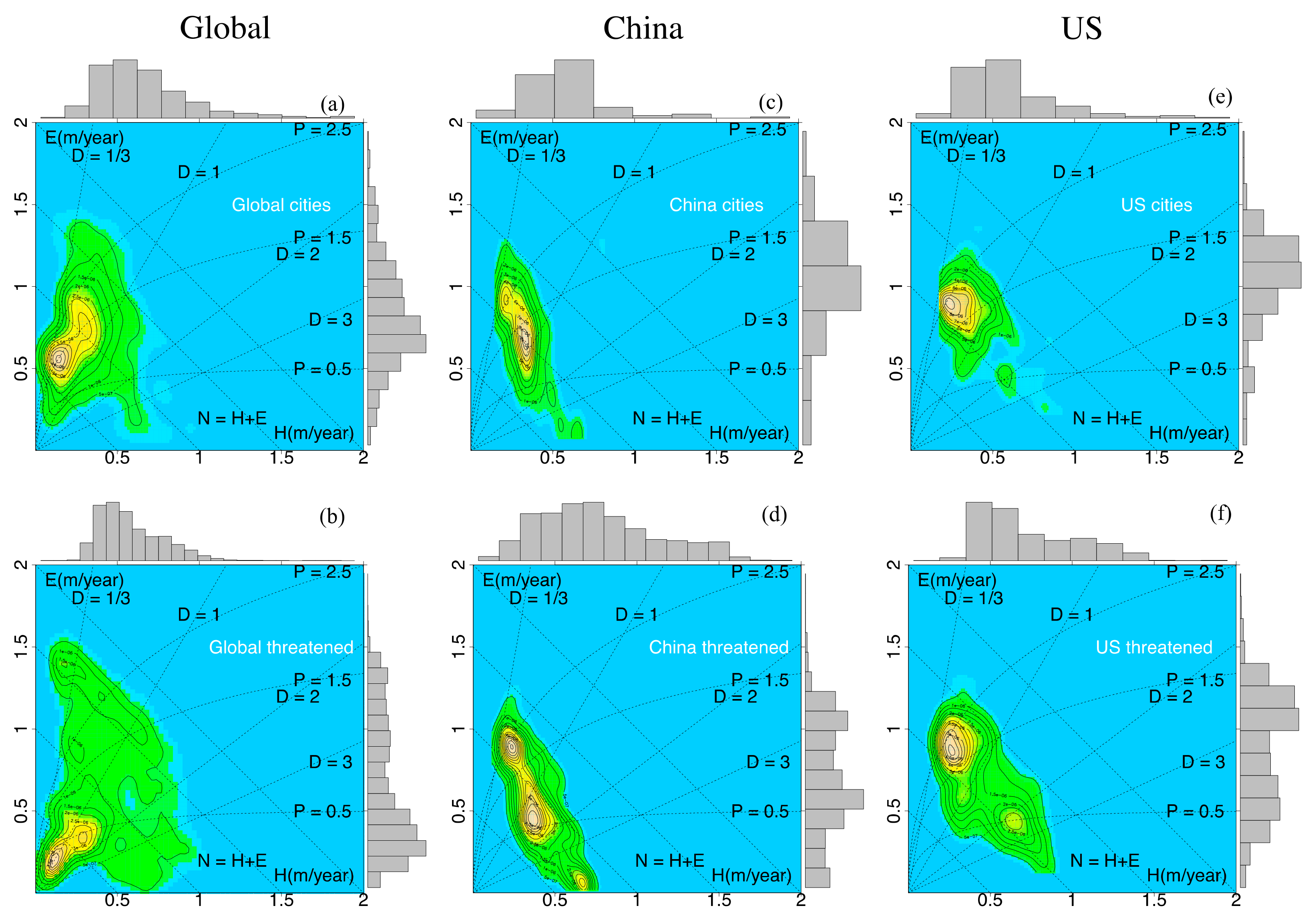
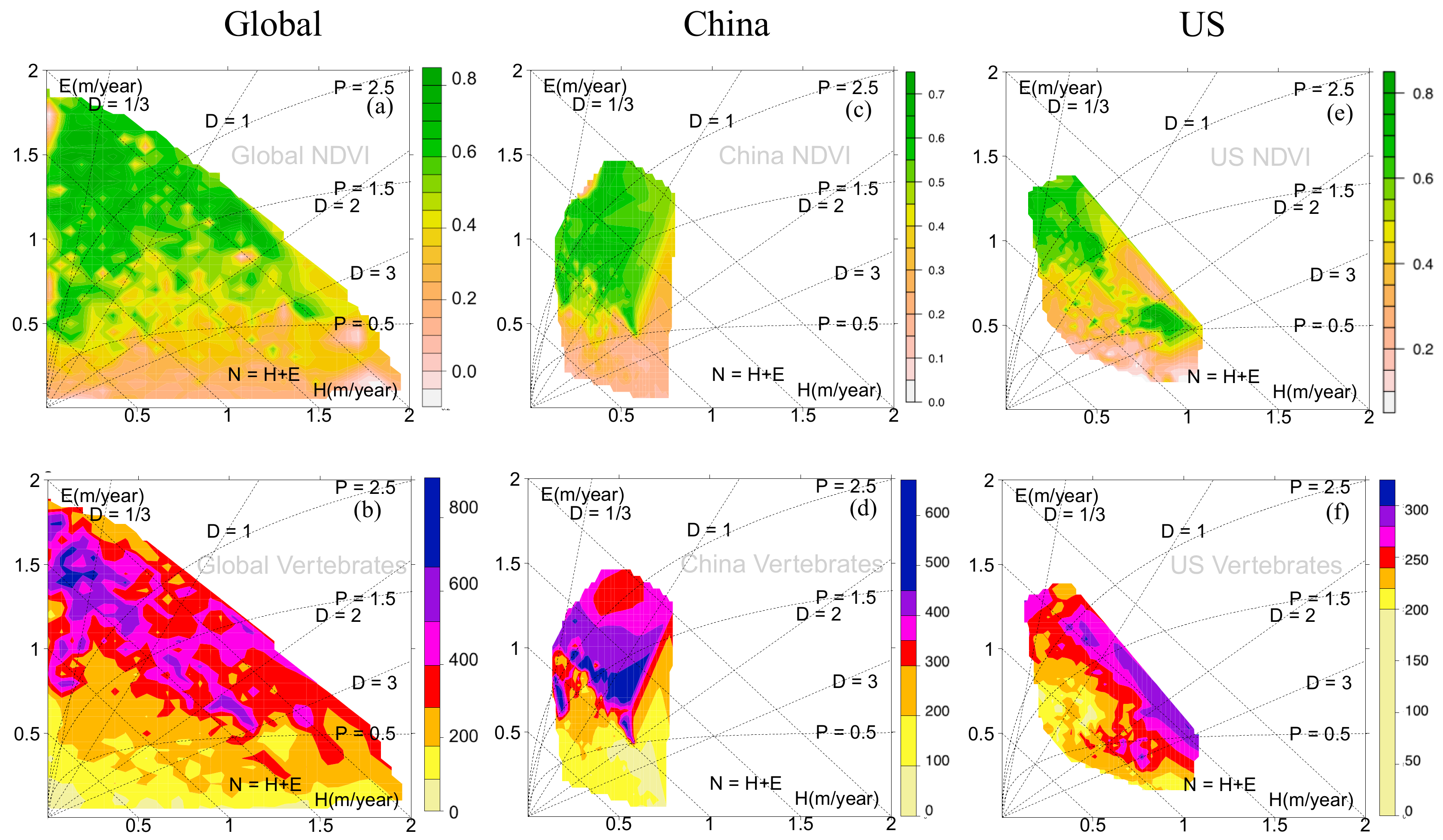
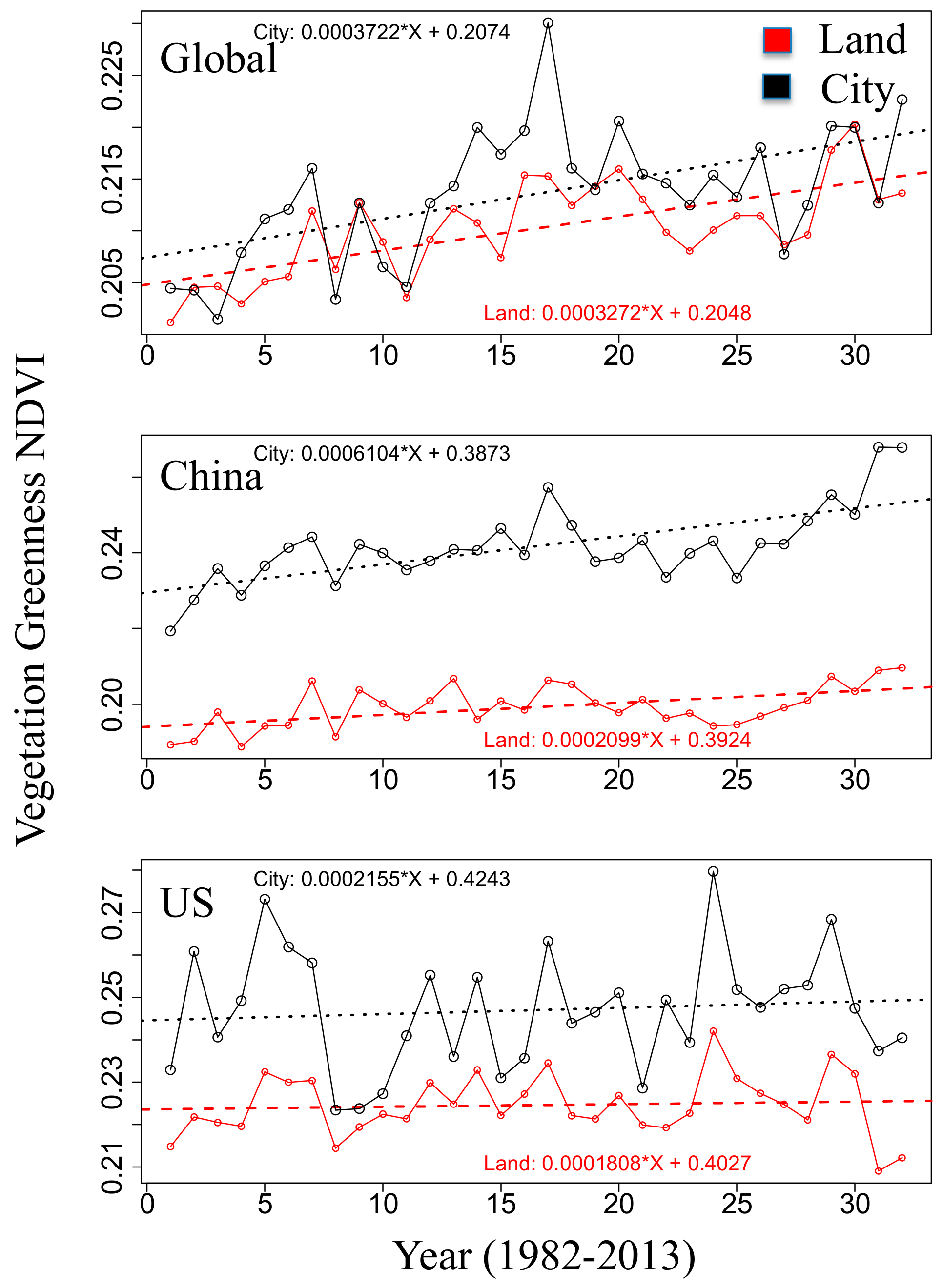
3.1. Eco-Hydrological Embedding
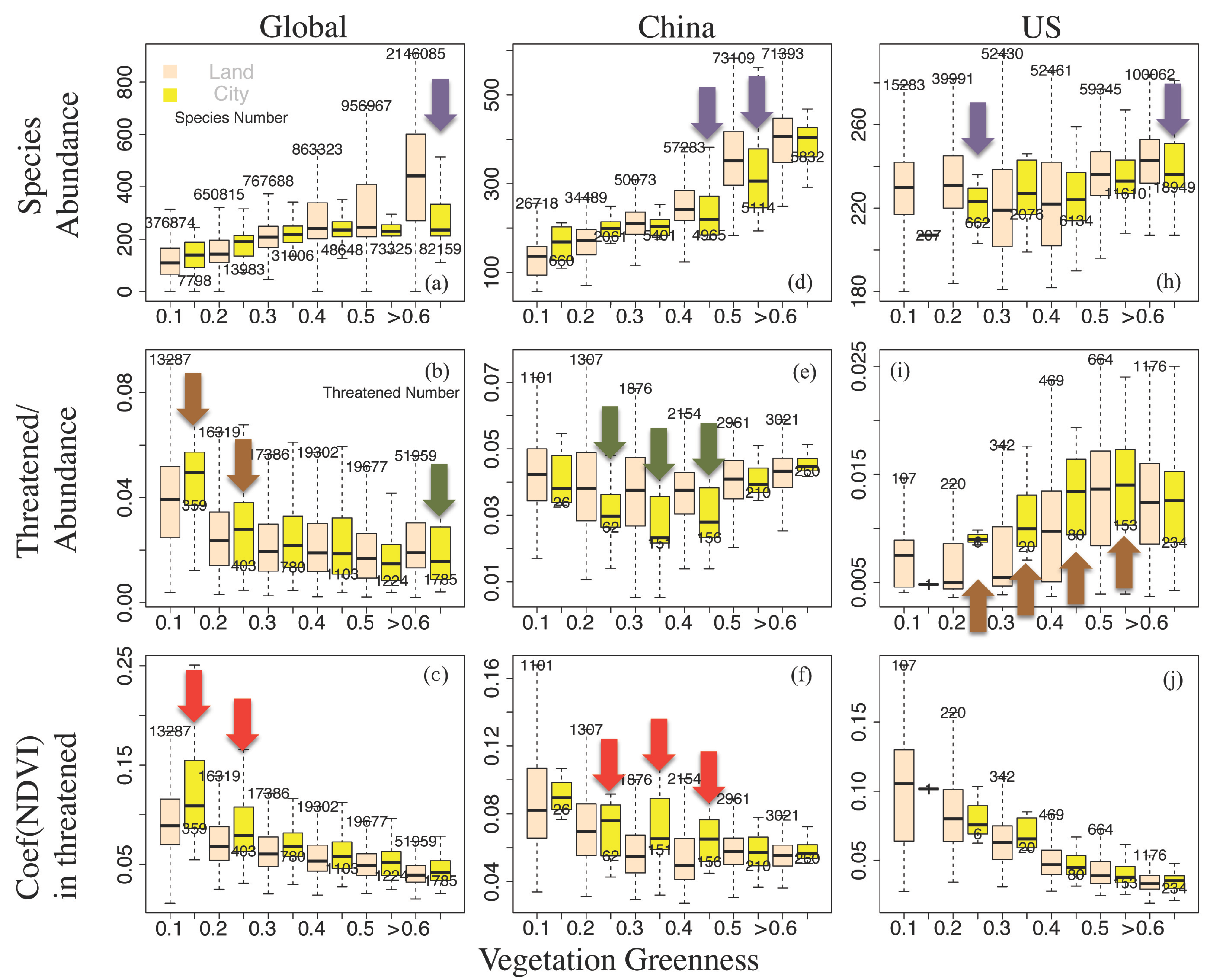
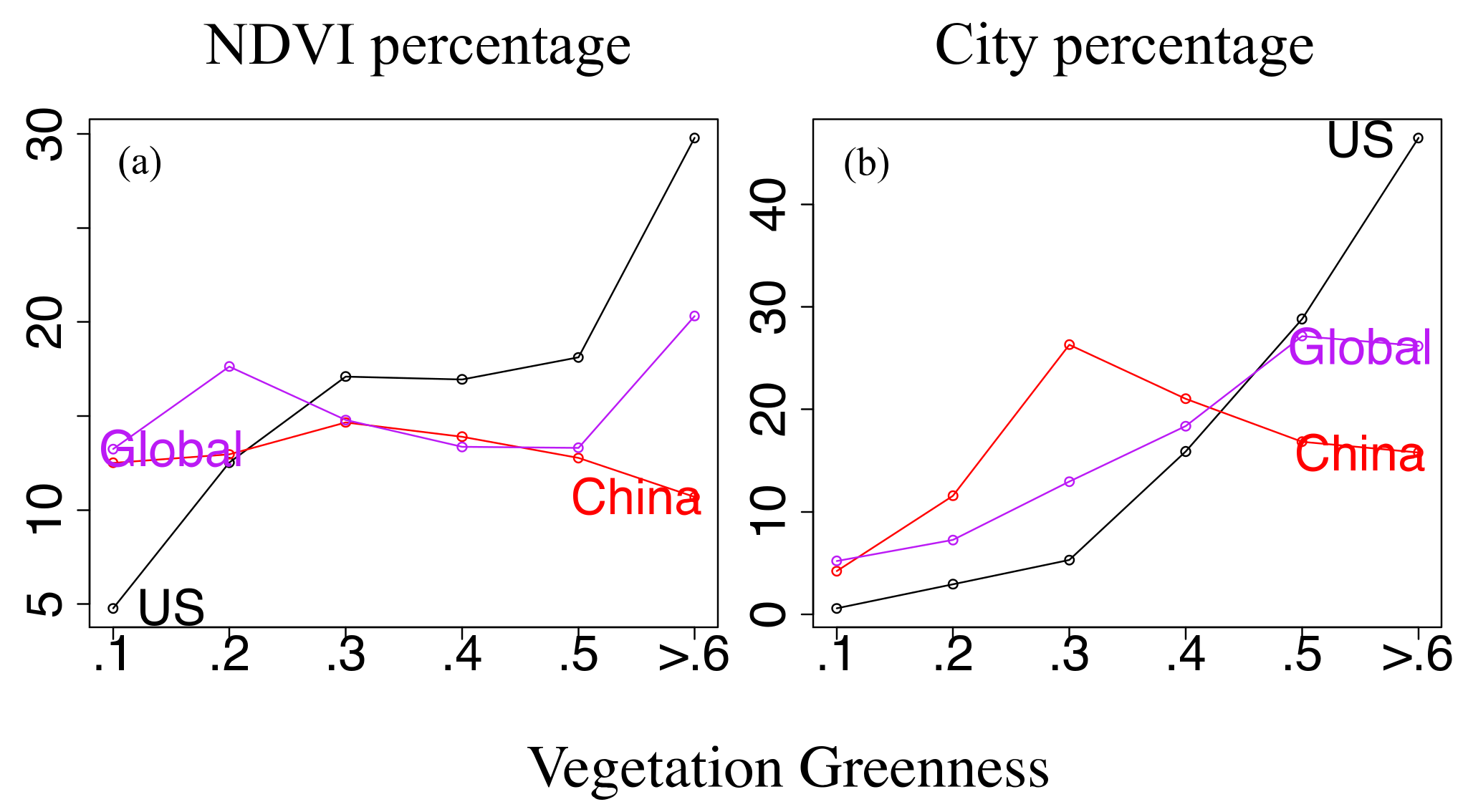
3.2. Urbanization, Vegetation, and Vertebrates
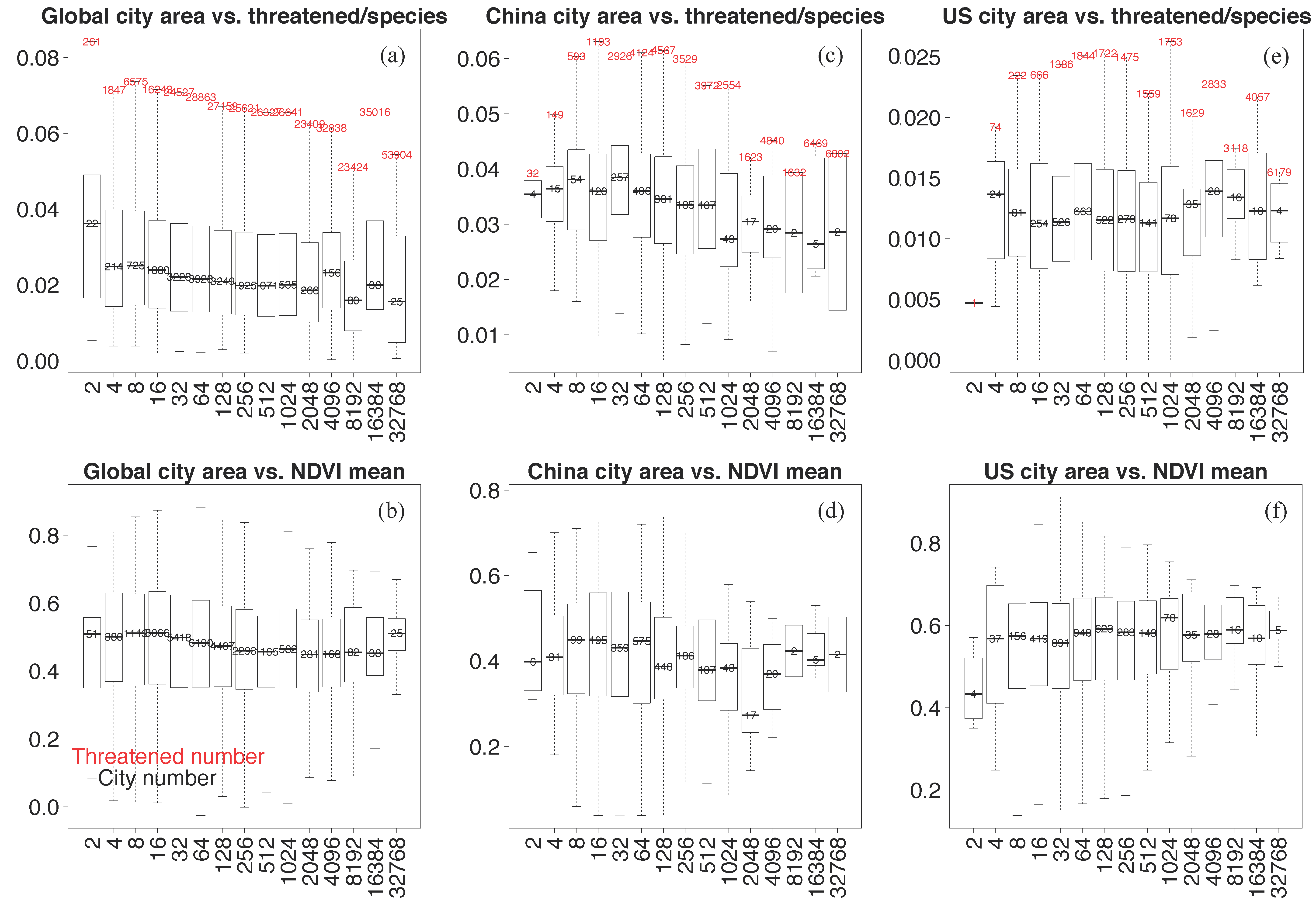
3.3. City Size Relation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- He, S.; Zhong, Y.; Sun, Y.; Su, Z.; Jia, X.; Hu, Y.; Zhou, Q. Topography-associated thermal gradient predicts warming effects on woody plant structural diversity in a subtropical forest. Sci. Rep. 2017, 7, 40387. [Google Scholar] [CrossRef] [PubMed]
- Hurlbert, A.H.; Jetz, W. Species richness, hotspots, and the scale dependence of range maps in ecology and conservation. Proc. Natl. Acad. Sci. USA 2007, 104, 13384–13389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, E2602–E2610. [Google Scholar] [CrossRef] [PubMed]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.; Romdal, T.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–219. [Google Scholar] [CrossRef] [Green Version]
- United Nations (UN). World Urbanization Prospects, 2009 Revision; United Nations: New York, NY, USA, 2010. [Google Scholar]
- Cai, D.; Fraedrich, K.; Guan, Y.; Guo, S.; Zhang, C. Urbanization and the thermal environment of Chinese and American cities. Sci. Total Environ. 2017, 589, 200–211. [Google Scholar] [CrossRef]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 2016, 7, 12558. [Google Scholar] [CrossRef] [Green Version]
- Von Humboldt, A. Ansichten der Natur, Mit Wissenschaflichen Erläuterungen; JG Cotta: Stuttgart, Germany, 1826. [Google Scholar]
- Currie, D.J. Energy and large-scale patterns of animal-and plant-species richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Porter, E.E. Does herbivore diversity depend on plant diversity? The case of California butterflies. Am. Nat. 2002, 161, 40–49. [Google Scholar] [CrossRef]
- Field, R.; Hawkins, B.A.; Cornell, H.V.; Currie, D.J.; Diniz-Filho, J.A.F.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T. Spatial species-richness gradients across scales: A meta-analysis. J. Biogeogr. 2009, 36, 132–147. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, D.; de Laender, F. The combined effects of biotic and abiotic stress on species richness and connectance. PLoS ONE 2017, 12, e0172828. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Cai, D.; Fraedrich, K.; Sielmann, F.; Zhang, L.; Zhu, X.; Guo, S.; Guan, Y. Vegetation dynamics on the Tibetan Plateau (1982–2006): An attribution by eco-hydrological diagnostics. J. Clim. 2015, 28, 4576–4584. [Google Scholar] [CrossRef]
- Cai, D.; Fraedrich, K.; Sielmann, F.; Guan, Y.; Guo, S. Land-cover characterization and aridity changes of South America (1982–2006): An attribution by ecohydrological diagnostics. J. Clim. 2016, 29, 8175–8189. [Google Scholar] [CrossRef]
- Stephenson, N.L. Climatic control of vegetation distribution: The role of the water balance. Am. Nat. 1990, 135, 649–670. [Google Scholar] [CrossRef]
- Frolking, S.; Milliman, T.; Seto, K.C.; Friedl, M.A. A global fingerprint of macro-scale changes in urban structure from 1999 to 2009. Environ. Res. Lett. 2013, 8, 024004. [Google Scholar] [CrossRef]
- Šímová, I.; Storch, D. The enigma of terrestrial primary productivity: Measurements, models, scales and the diversity–productivity relationship. Ecography 2017, 40, 239–252. [Google Scholar] [CrossRef]
- Oindo, B.O.; Skidmore, A.K. Interannual variability of NDVI and species richness in Kenya. Int. J. Remote Sens. 2002, 23, 285–298. [Google Scholar] [CrossRef] [Green Version]
- Balsamo, G.; Albergel, C.; Beljaars, A.; Boussetta, S.; Brun, E.; Cloke, H.; Dee, D.; Dutra, E.; Muñoz-Sabater, J.; Pappenberger, F. ERA-Interim/Land: A global land surface reanalysis data set. Hydrol. Earth Syst. Sci. 2015, 19, 389–407. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; Saleous, N.E. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Elvidge, C.D.; Baugh, K.E.; Kihn, E.A.; Kroehl, H.W.; Davis, E.R. Mapping city lights with nighttime data from the DMSP Operational Linescan System. Photogramm. Eng. Remote Sens. 1997, 63, 727–734. [Google Scholar]
- Small, C.; Elvidge, C.D.; Balk, D.; Montgomery, M. Spatial scaling of stable night lights. Remote Sens. Environ. 2011, 115, 269–280. [Google Scholar] [CrossRef]
- Budyko, M. Climate and Life; Academic Press: New York, NY, USA, 1974; Volume 508. [Google Scholar]
- Schreiber, P. Über die Beziehungen zwischen dem Niederschlag und der Wasserführung der Flüsse in Mitteleuropa. Z. Meteorol. 1904, 21, 441–452. [Google Scholar]
- Fraedrich, K. A parsimonious stochastic water reservoir: Schreiber’s 1904 equation. J. Hydrometeorol. 2010, 11, 575–578. [Google Scholar] [CrossRef]
- Sposito, G. Understanding the budyko equation. Water 2017, 9, 236. [Google Scholar] [CrossRef]
- Adamic, L.A.; Bernardo, A.H. Zipf’s law and the Internet. Glottometrics 2002, 3, 143–150. [Google Scholar]
- Cai, D.; Fraedrich, K.; Sielmann, F.; Guan, Y.; Guo, S.; Zhang, L.; Zhu, X. Climate and vegetation: An ERA-interim and GIMMS NDVI analysis. J. Clim. 2014, 27, 5111–5118. [Google Scholar] [CrossRef]
- Madonsela, S.; Cho, M.A.; Ramoelo, A.; Mutanga, O. Remote sensing of species diversity using Landsat 8 spectral variables. ISPRS J. Photogramm. Remote Sens. 2017, 133, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Kraas, F.; Leggewie, C.; Lemke, P.; Matthies, E.; Messner, D.; Nakicenovic, N.; Schellnhuber, H.J.; Schlacke, S.; Schneidewind, U.; Brandi, C. Humanity on the Move: Unlocking the Transformative Power of Cities; WBGU-German Advisory Council on Global Change: Berlin, Germany, 2016. [Google Scholar]
- Pimm, S.L.; Jenkins, C. Sustaining the variety of life. Sci. Am. 2005, 293, 66–73. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| China (%) | USA (%) | Global | |
|---|---|---|---|
| a) Total Land Area | 6.3 | 6.1 | 148,940,000 km2 |
| b) Vertebrates Abundance | 5.9 | 5.4 | 5,930,794 number |
| Amphibian Birds Mammal | 4.4 5.9 6.1 | 7.3 5.2 5.8 | 283,256 4,376,655 1,270,883 |
| c) Threatened Vertebrates | 9.6 | 2.1 | 145,187 number |
| Amphibian Birds Mammal | 29.5 7.5 12.2 | 5.3 2.7 0.8 | 2,892 92,513 49,782 |
| d) Cities*/Non-Cities in Continental/Global Land | 6.0/94.0 | 12.3/87.7 | 4.5/95.5% |
| Threatened Amphibian in/Out Cities Threatened Birds in/Out Cities Threatened Mammal in/Out Cities | 7.0/93.0 8.0/92.0 4.4/95.6 | 9.8/90.2 16.1/83.9 22.3/77.7 | 7.8/92.2 4.3/95.7 3.3/96.7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, D.; Fraedrich, K.; Guan, Y.; Guo, S.; Zhang, C.; Carvalho, L.M.V.; Zhu, X. Causality of Biodiversity Loss: Climate, Vegetation, and Urbanization in China and America. Sensors 2019, 19, 4499. https://0-doi-org.brum.beds.ac.uk/10.3390/s19204499
Cai D, Fraedrich K, Guan Y, Guo S, Zhang C, Carvalho LMV, Zhu X. Causality of Biodiversity Loss: Climate, Vegetation, and Urbanization in China and America. Sensors. 2019; 19(20):4499. https://0-doi-org.brum.beds.ac.uk/10.3390/s19204499
Chicago/Turabian StyleCai, Danlu, Klaus Fraedrich, Yanning Guan, Shan Guo, Chunyan Zhang, Leila M.V. Carvalho, and Xiuhua Zhu. 2019. "Causality of Biodiversity Loss: Climate, Vegetation, and Urbanization in China and America" Sensors 19, no. 20: 4499. https://0-doi-org.brum.beds.ac.uk/10.3390/s19204499