
Glossogyne tenuifolia Extract Increases Nitric Oxide Production in Human Umbilical Vein Endothelial Cells

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Cell Viability
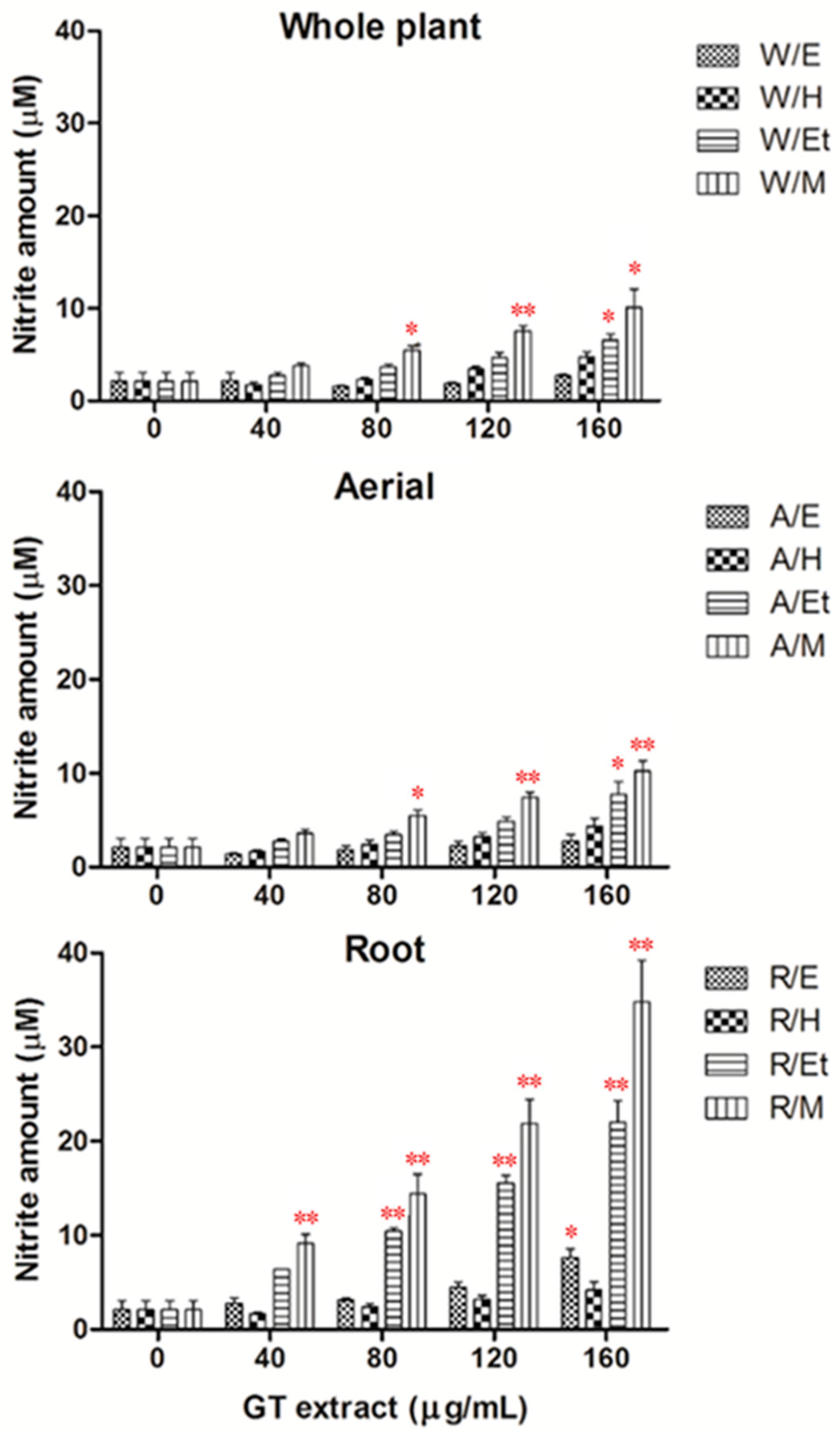
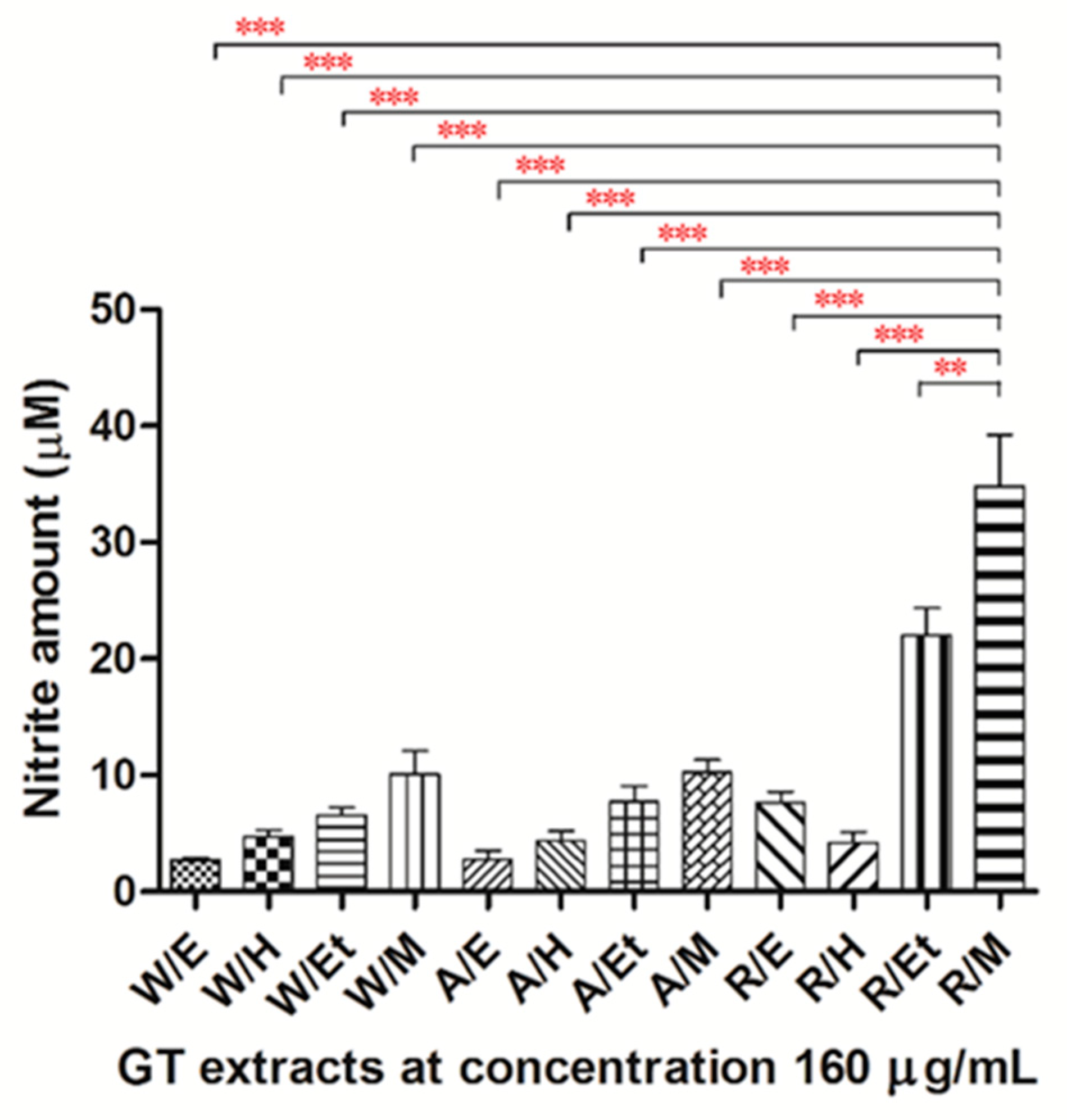
2.2. Effects of Extracts of GT on NO Production in HUVECs
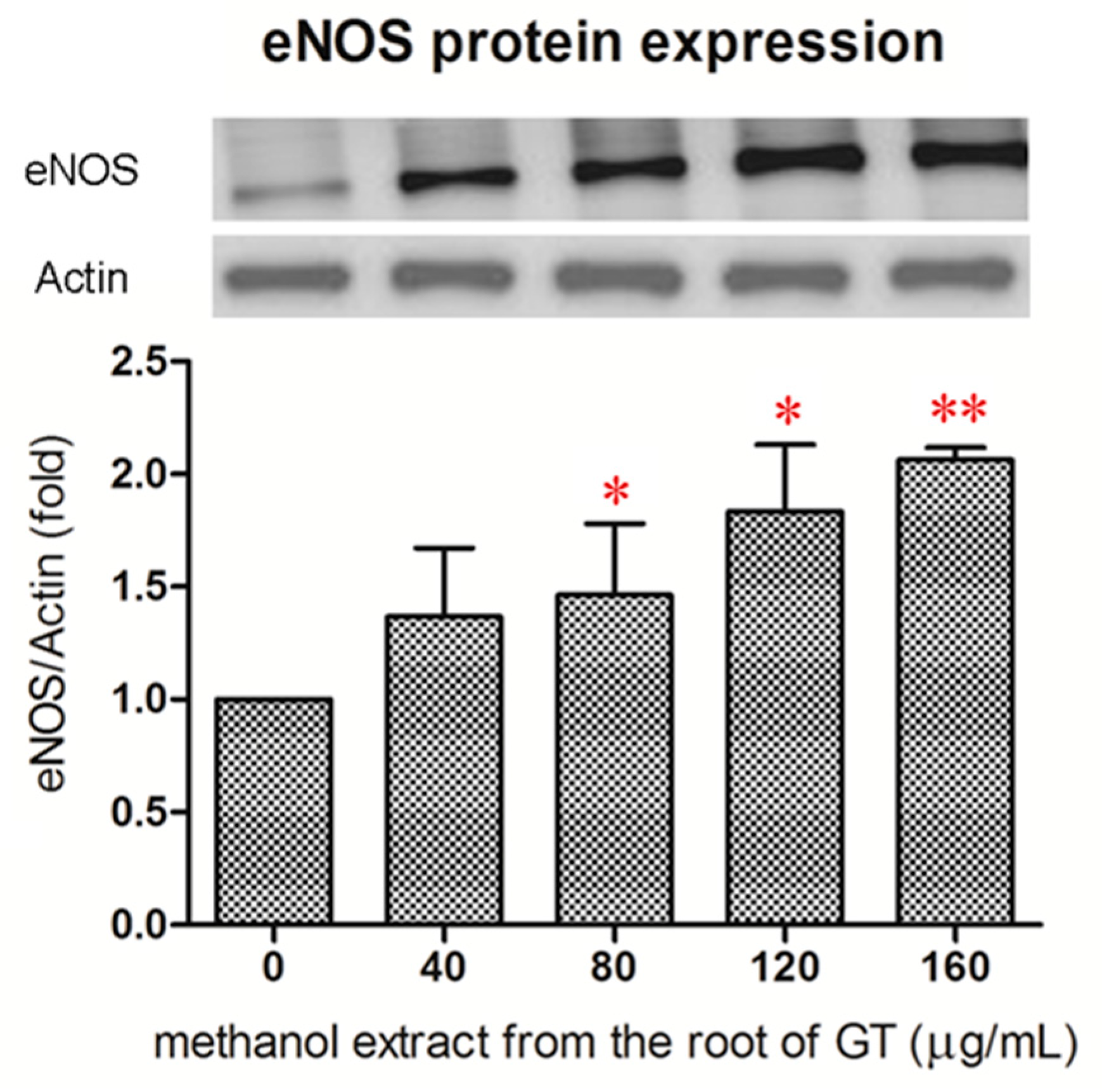
2.3. Effect of the R/M GTE on eNOS Protein Expression
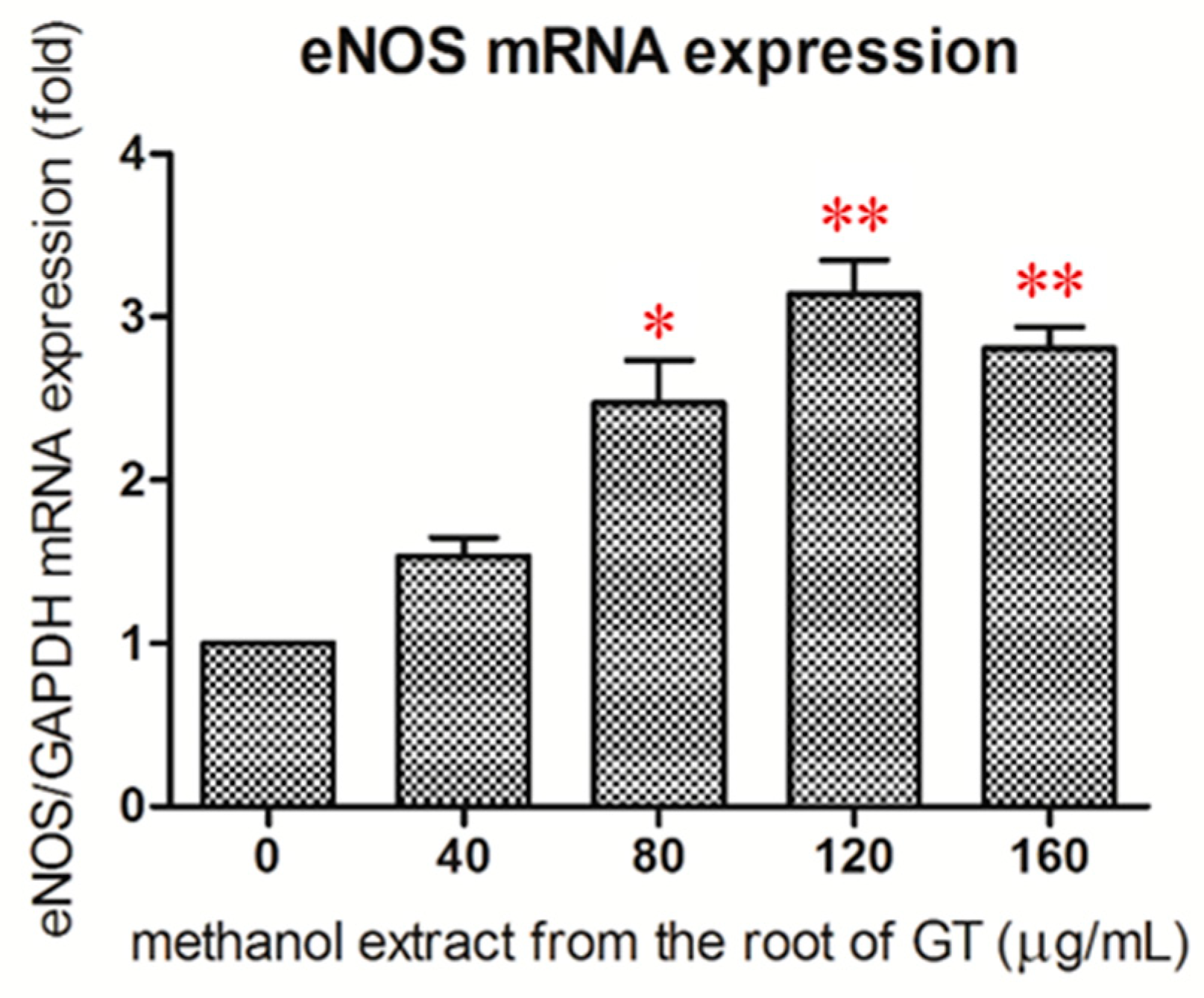
2.4. Effect of the R/M GTE on eNOS mRNA Expression
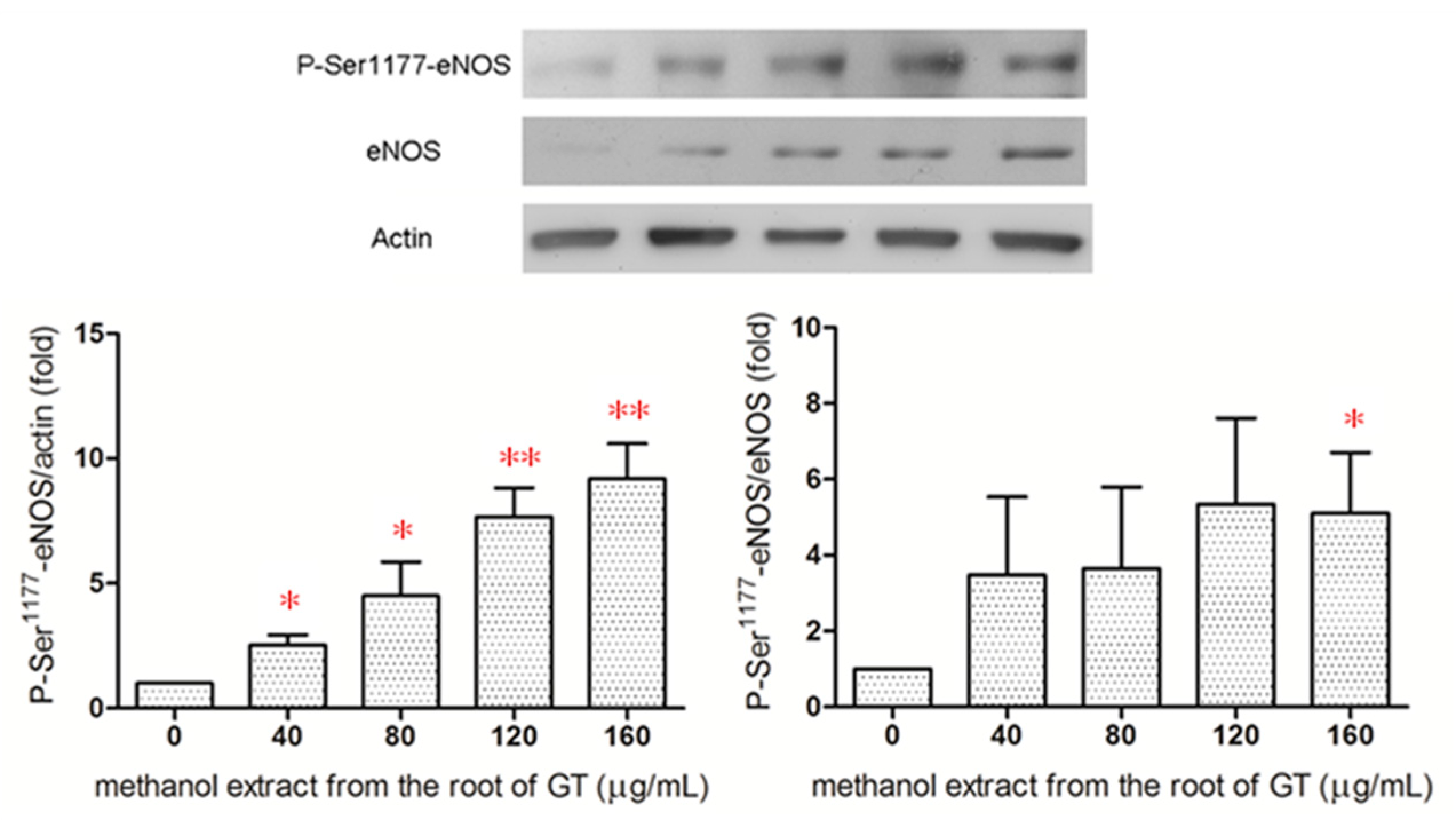
2.5. Effect of the R/M GTE on eNOS Phosphorylation at Ser1177
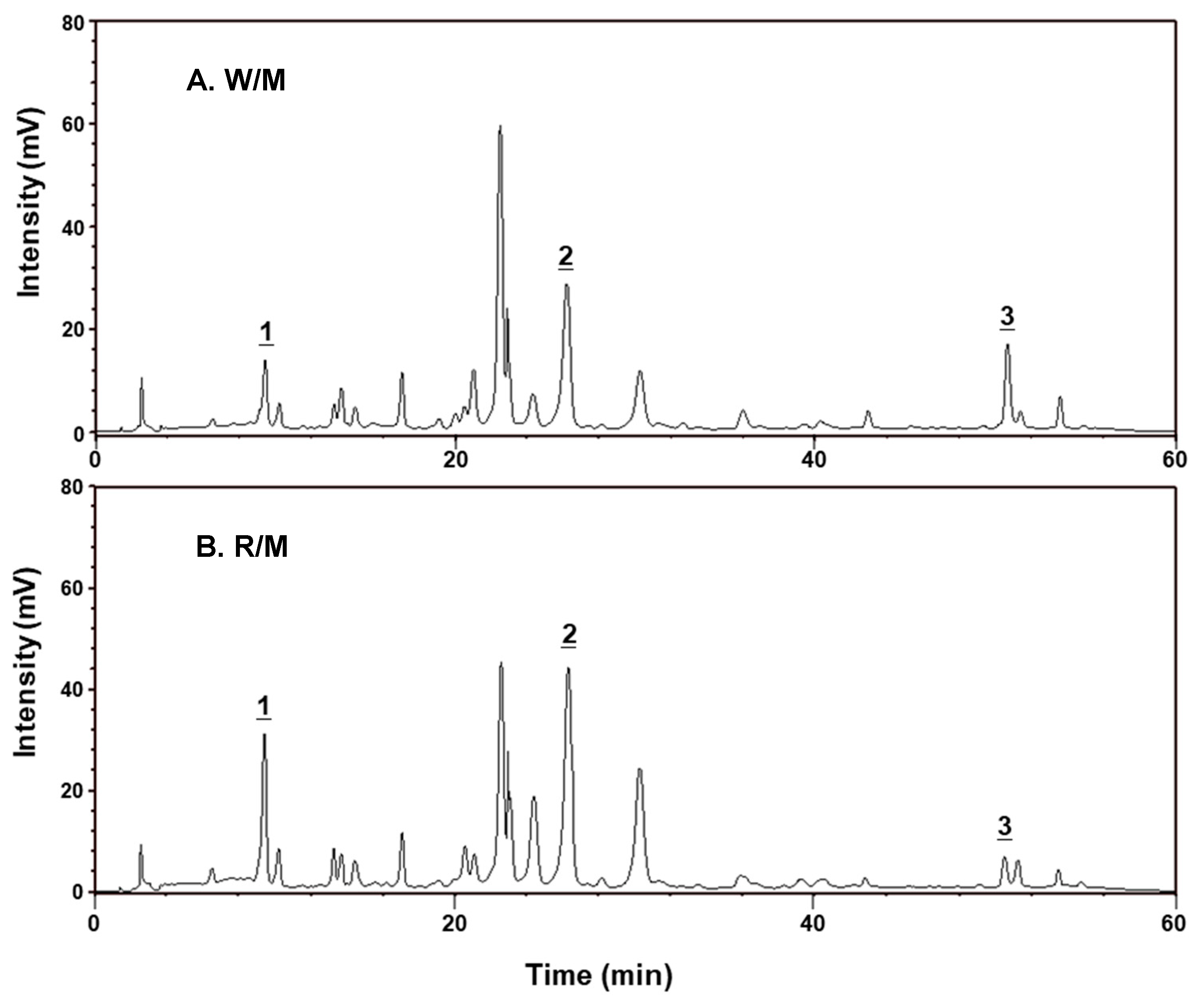
2.6. Chemical Composition Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures
4.3. MTT Assay for Cell Viability
4.4. Determination of NO Synthesis
4.5. Western Blot Analysis
4.6. Real-Time RT-PCR Analysis
4.7. Chemical Composition Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rosenblit, P.D. Extreme atherosclerotic cardiovascular disease (ASCVD) risk recognition. Curr. Diab. Rep. 2019, 19, 61. [Google Scholar] [CrossRef] [PubMed]
- Davignon, J.; Ganz, P. Role of endothelial dysfunction in atherosclerosis. Circulation 2004, 109, III27–III32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassi, D.; Desideri, G.; Di Giosia, P.; De Feo, M.; Fellini, E.; Cheli, P.; Ferri, L.; Ferri, C. Tea, flavonoids, and cardiovascular health: Endothelial protection. Am. J. Clin. Nutr. 2013, 98, 1660S–1666S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Förstermann, U. Prevention of atherosclerosis by interference with the vascular nitric oxide system. Curr. Pharm. Des. 2009, 15, 3133–3145. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Horke, S.; Förstermann, U. Vascular oxidative stress, nitric oxide and atherosclerosis. Atherosclerosis 2014, 237, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Gliozzi, M.; Scicchitano, M.; Bosco, F.; Musolino, V.; Carresi, C.; Scarano, F.; Maiuolo, J.; Nucera, S.; Maretta, A.; Paone, S.; et al. Modulation of nitric oxide synthases by oxidized LDLs: Role in vascular inflammation and atherosclerosis development. Int. J. Mol. Sci. 2019, 20, 3294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeiher, A.M.; Krause, T.; Schächinger, V.; Minners, J.; Moser, E. Impaired endothelium-dependent vasodilation of coronary resistance vessels is associated with exercise-induced myocardial ischemia. Circulation 1995, 91, 2345–2352. [Google Scholar] [CrossRef]
- Zhong, Q.; Nong, Q.; Mao, B.; Pan, X.; Meng, L. Association of impaired vascular endothelial function with increased cardiovascular risk in asymptomatic adults. Biomed. Res. Int. 2018, 2018, 3104945. [Google Scholar] [CrossRef] [Green Version]
- Halcox, J.P.; Schenke, W.H.; Zalos, G.; Mincemoyer, R.; Prasad, A.; Waclawiw, M.A.; Nour, K.R.; Quyyumi, A.A. Prognostic value of coronary vascular endothelial dysfunction. Circulation 2002, 106, 653–658. [Google Scholar] [CrossRef]
- Malekmohammad, K.; Sewell, R.D.E.; Rafieian-Kopaei, M. Mechanisms of medicinal plant activity on nitric oxide (NO) bioavailability as prospective treatments for atherosclerosis. Curr. Pharm. Des. 2020, 26, 2591–2601. [Google Scholar] [CrossRef]
- Azumi, H.; Inoue, N.; Ohashi, Y.; Terashima, M.; Mori, T.; Fujita, H.; Awano, K.; Kobayashi, K.; Maeda, K.; Hata, K.; et al. Superoxide generation in directional coronary atherectomy specimens of patients with angina pectoris: Important role of NAD(P)H oxidase. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Heistad, D.D.; Wakisaka, Y.; Miller, J.; Chu, Y.; Pena-Silva, R. Novel aspects of oxidative stress in cardiovascular diseases. Circ. J. 2009, 73, 201–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, R.; Hecker, M.; Stahmann, N.; Thiele, J.J.; Werner-Felmayer, G.; Werner, E.R. α-Tocopherol amplifies phosphorylation of endothelial nitric oxide synthase at serine 1177 and its short-chain derivative trolox stabilizes tetrahydrobiopterin. Free Radic. Biol. Med. 2004, 37, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Ladurner, A.; Schmitt, C.A.; Schachner, D.; Atanasov, A.G.; Werner, E.R.; Dirsch, V.M.; Heiss, E.H. Ascorbate stimulates endothelial nitric oxide synthase enzyme activity by rapid modulation of its phosphorylation status. Free Radic. Biol. Med. 2012, 52, 2082–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biegańska-Hensoldt, S.; Rosołowska-Huszcz, D. Polyphenols in preventing endothelial dysfunction. Postepy Hig. Med. Dosw. 2017, 71, 227–235. [Google Scholar] [CrossRef]
- Kim, D.H.; Meza, C.A.; Clarke, H.; Kim, J.S.; Hickner, R.C. Vitamin D and endothelial function. Nutrients 2020, 12, 575. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, W.C.; Hsieh, M.T.; Chiu, N.Y.; Lin, J.K. Investigation and study on the resources of Chinese crude drugs in Taiwan (VI): Investigation and study on the medicinal plants in Penghu County. Annu. Rep. Chin. Med. 1999, 17, 245–407. [Google Scholar]
- Wu, M.J.; Wang, L.; Ding, H.Y.; Weng, C.Y.; Yen, J.H. Glossogyne tenuifolia acts to inhibit inflammatory mediator production in a macrophage cell line by downregulating LPS-induced NF-κB. J. Biomed. Sci. 2004, 11, 186–199. [Google Scholar] [CrossRef]
- Wu, M.J.; Weng, C.Y.; Ding, H.Y.; Wu, P.J. Anti-inflammatory and antiviral effects of Glossogyne tenuifolia. Life Sci. 2005, 76, 1135–1146. [Google Scholar] [CrossRef]
- Ha, C.L.; Weng, C.Y.; Wang, L.; Lian, T.W.; Wu, M.J. Immunomodulatory effect of Glossogyne tenuifolia in murine peritoneal macrophages and splenocytes. J. Ethnopharmacol. 2006, 107, 116–125. [Google Scholar] [CrossRef]
- Hsu, H.F.; Houng, J.Y.; Chang, C.L.; Wu, C.C.; Chang, F.R.; Wu, Y.C. Antioxidant activity, cytotoxicity, and DNA information of Glossogyne tenuifolia. J. Agric. Food Chem. 2005, 53, 6117–6125. [Google Scholar] [CrossRef]
- Wu, M.J.; Huang, C.L.; Lian, T.W.; Kou, M.C.; Wang, L. Antioxidant activity of Glossogyne tenuifolia. J. Agric. Food Chem. 2005, 53, 6305–6312. [Google Scholar] [CrossRef]
- Yang, J.H.; Tsai, S.Y.; Han, C.M.; Shih, C.C.; Mau, J.L. Antioxidant properties of Glossogyne tenuifolia. Am. J. Chin. Med. 2006, 34, 707–720. [Google Scholar] [CrossRef]
- Wang, S.W.; Kuo, H.C.; Hsu, H.F.; Tu, Y.K.; Cheng, T.T.; Houng, J.Y. Inhibitory activity on RANKL-mediated osteoclastogenesis of Glossogyne tenuifolia extract. J. Funct. Foods 2014, 6, 215–223. [Google Scholar] [CrossRef]
- Wang, C.P.; Houng, J.Y.; Hsu, H.F.; Chen, H.J.; Huang, B.; Hung, W.C.; Yu, T.H.; Chiu, C.A.; Lu, L.F.; Hsu, C.C. Glossogyne tenuifolia enhances posttranslational S-nitrosylation of proteins in vascular endothelial cells. Taiwania 2011, 56, 97–104. [Google Scholar]
- Hsuan, C.F.; Hsu, H.F.; Tseng, W.K.; Lee, T.L.; Wei, Y.F.; Hsu, K.L.; Wu, C.C.; Houng, J.Y. Glossogyne tenuifolia extract inhibits TNF-α-induced expression of adhesion molecules in human umbilical vein endothelial cells via blocking the NF-κB signaling pathway. Molecules 2015, 20, 16908–16923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsuan, C.F.; Lu, Y.C.; Tsai, I.T.; Houng, J.Y.; Wang, S.W.; Chang, T.H.; Chen, Y.L.; Chang, C.C. Glossogyne tenuifolia attenuates proliferation and migration of vascular smooth muscle cells. Molecules 2020, 25, 5832. [Google Scholar] [CrossRef] [PubMed]
- Mount, P.E.; Kemp, B.E.; Power, D.A. Regulation of endothelial and myocardial NO synthesis by multi-site eNOS phosphorylation. J. Mol. Cell. Cardiol. 2007, 42, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.D.; Hsu, H.F.; Chen, Z.H.; Wang, Y.T.; Huang, S.H.; Chen, H.J.; Wang, C.P.; Wang, S.W.; Chang, C.C.; Houng, J.Y. Antioxidant, anti-inflammatory, and anti-proliferative activities of extracts from different parts of farmed and wild Glossogyne tenuifolia. Ind. Crop Prod. 2014, 57, 98–105. [Google Scholar] [CrossRef]
- Li, H.; Förstermann, U. Nitric oxide in the pathogenesis of vascular disease. J. Pathol. 2000, 190, 244–254. [Google Scholar] [CrossRef]
- Hong, F.F.; Liang, X.Y.; Liu, W.; Lv, S.; He, S.J.; Kuang, H.B.; Yang, S.L. Roles of eNOS in atherosclerosis treatment. Inflamm. Res. 2019, 68, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, K.; Kawashima, S.; Mikami, S.; Miwa, Y.; Hirata, K.; Suematsu, M.; Hayashi, Y.; Itoh, H.; Yokoyama, M. Endothelial constitutive nitric oxide synthase protein and mRNA increased in rabbit atherosclerotic aorta despite impaired endothelium-dependent vascular relaxation. Am. J. Pathol. 1996, 148, 1949–1956. [Google Scholar]
- d’Uscio, L.V.; Smith, L.A.; Katusic, Z.S. Hypercholesterolemia impairs endothelium-dependent relaxations in common carotid arteries of apolipoprotein e-deficient mice. Stroke 2001, 32, 2658–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambardella, J.; Khondkar, W.; Morelli, M.B.; Wang, X.; Santulli, G.; Trimarco, V. Arginine and endothelial function. Biomedicines 2020, 8, 277. [Google Scholar] [CrossRef] [PubMed]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction methods, antioxidative action, bioavailability and anticarcinogenic effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H.B. Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.Y.; Meng, X.; Gan, R.Y.; Zhao, C.N.; Liu, Q.; Feng, Y.B.; Li, S.; Wei, X.L.; Atanasov, A.G.; Corke, H.; et al. Health functions and related molecular mechanisms of tea components: An update review. Int. J. Mol. Sci. 2019, 20, 6196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.; Hodgson, J.M.; Mas, E.; Croft, K.D.; Ward, N.C. Chlorogenic acid improves ex vivo vessel function and protects endothelial cells against HOCl-induced oxidative damage, via increased production of nitric oxide and induction of Hmox-1. J. Nutr. Biochem. 2016, 27, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hada, Y.; Uchida, H.A.; Otaka, N.; Onishi, Y.; Okamoto, S.; Nishiwaki, M.; Takemoto, R.; Takeuchi, H.; Wada, J. The protective effect of chlorogenic acid on vascular senescence via the Nrf2/HO-1 pathway. Int. J. Mol. Sci. 2020, 21, 4527. [Google Scholar] [CrossRef]
- Qin, L.; Zang, M.; Xu, Y.; Zhao, R.; Wang, Y.; Mi, Y.; Mei, Y. Chlorogenic acid alleviates hyperglycemia-induced cardiac fibrosis through activation of the NO/cGMP/PKG pathway in cardiac fibroblasts. Mol. Nutr. Food Res. 2021, 65, e2000810. [Google Scholar] [CrossRef]
- Li, H.; Xia, N.; Brausch, I.; Yao, Y.; Förstermann, U. Flavonoids from artichoke (Cynara scolymus L.) up-regulate endothelial-type nitric-oxide synthase gene expression in human endothelial cells. J. Pharmacol. Exp. Ther. 2004, 310, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.T.; Qian, L.B.; Zhang, F.J.; Wang, J.; Ai, H.; Tang, L.H.; Wang, H.P. Cardioprotective effects of luteolin on ischemia/reperfusion injury in diabetic rats are modulated by eNOS and the mitochondrial permeability transition pathway. J. Cardiovasc. Pharmacol. 2015, 65, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Xia, M.L.; Wang, J.; Zhou, X.R.; Lou, Y.Y.; Tang, L.H.; Zhang, F.J.; Yang, J.T.; Qian, L.B. Luteolin attenuates cardiac ischemia/reperfusion injury in diabetic rats by modulating Nrf2 antioxidative function. Oxid. Med. Cell. Longev. 2019, 2019, 2719252. [Google Scholar] [CrossRef] [PubMed]
- Si, H.; Wyeth, R.P.; Liu, D. The flavonoid luteolin induces nitric oxide production and arterial relaxation. Eur. J. Nutr. 2014, 53, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamkaew, N.; Paracha, T.U.; Ingkaninan, K.; Waranuch, N.; Chootip, K. Vasodilatory effects and mechanisms of action of Bacopa monnieri active compounds on rat mesenteric arteries. Molecules 2019, 24, 2243. [Google Scholar] [CrossRef] [Green Version]
- Gentile, D.; Fornai, M.; Pellegrini, C.; Colucci, R.; Benvenuti, L.; Duranti, E.; Masi, S.; Carpi, S.; Nieri, P.; Nericcio, A.; et al. Luteolin prevents cardiometabolic alterations and vascular dysfunction in mice with HFD-induced obesity. Front Pharmacol. 2018, 9, 1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.I.; Hu, W.S.; Hung, M.Y.; Ou, H.C.; Huang, S.H.; Hsu, P.T.; Day, C.H.; Lin, K.H.; Viswanadha, V.P.; Kuo, W.W.; et al. Protective effects of luteolin against oxidative stress and mitochondrial dysfunction in endothelial cells. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 1032–1043. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Concentration (mg/g Extract) | |
|---|---|---|
| W/M GTE | R/M GTE | |
| Chlorogenic Acid (1) | 5.32 ± 0.13 | 8.59 ± 0.31 |
| Luteolin-7-glucoside (2) | 44.51 ± 0.27 | 63.02 ± 0.69 |
| Luteolin (3) | 4.23 ± 0.28 | 2.10 ± 0.11 |
| Total Polyphenols Content | 61.78 ± 0.84 | 76.72 ± 1.27 |
| Total Flavonoids Content | 58.56 ± 1.50 | 66.44 ± 1.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsuan, C.-F.; Lee, T.-L.; Tseng, W.-K.; Wu, C.-C.; Chang, C.-C.; Ko, T.-L.; Chen, Y.-L.; Houng, J.-Y. Glossogyne tenuifolia Extract Increases Nitric Oxide Production in Human Umbilical Vein Endothelial Cells. Pharmaceuticals 2021, 14, 577. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060577
Hsuan C-F, Lee T-L, Tseng W-K, Wu C-C, Chang C-C, Ko T-L, Chen Y-L, Houng J-Y. Glossogyne tenuifolia Extract Increases Nitric Oxide Production in Human Umbilical Vein Endothelial Cells. Pharmaceuticals. 2021; 14(6):577. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060577
Chicago/Turabian StyleHsuan, Chin-Feng, Thung-Lip Lee, Wei-Kung Tseng, Chau-Chung Wu, Chi-Chang Chang, Tsui-Ling Ko, Ya-Ling Chen, and Jer-Yiing Houng. 2021. "Glossogyne tenuifolia Extract Increases Nitric Oxide Production in Human Umbilical Vein Endothelial Cells" Pharmaceuticals 14, no. 6: 577. https://0-doi-org.brum.beds.ac.uk/10.3390/ph14060577