
Effect of Ducrosia flabellifolia and Savignya parviflora Extracts on Inhibition of Human Colon and Prostate Cancer Cell Lines

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Design and Statistical Analysis
2.2. Plant Materials
2.3. Preparation of Methanolic Extract
2.4. Analysis of the Total Content of Phenolics, Flavonoids and Antioxidant Effect of Plant Extracts
2.5. Cell Lines and Cell Cultures
2.6. In Vitro Cytotoxicity Assay
2.7. Apoptosis Detection
2.8. Cell Cycle Analysis
2.9. Detection of Expression Levels of Bax and Bcl-2 Genes
2.10. Real-Time Quantitative PCR (qPCR)
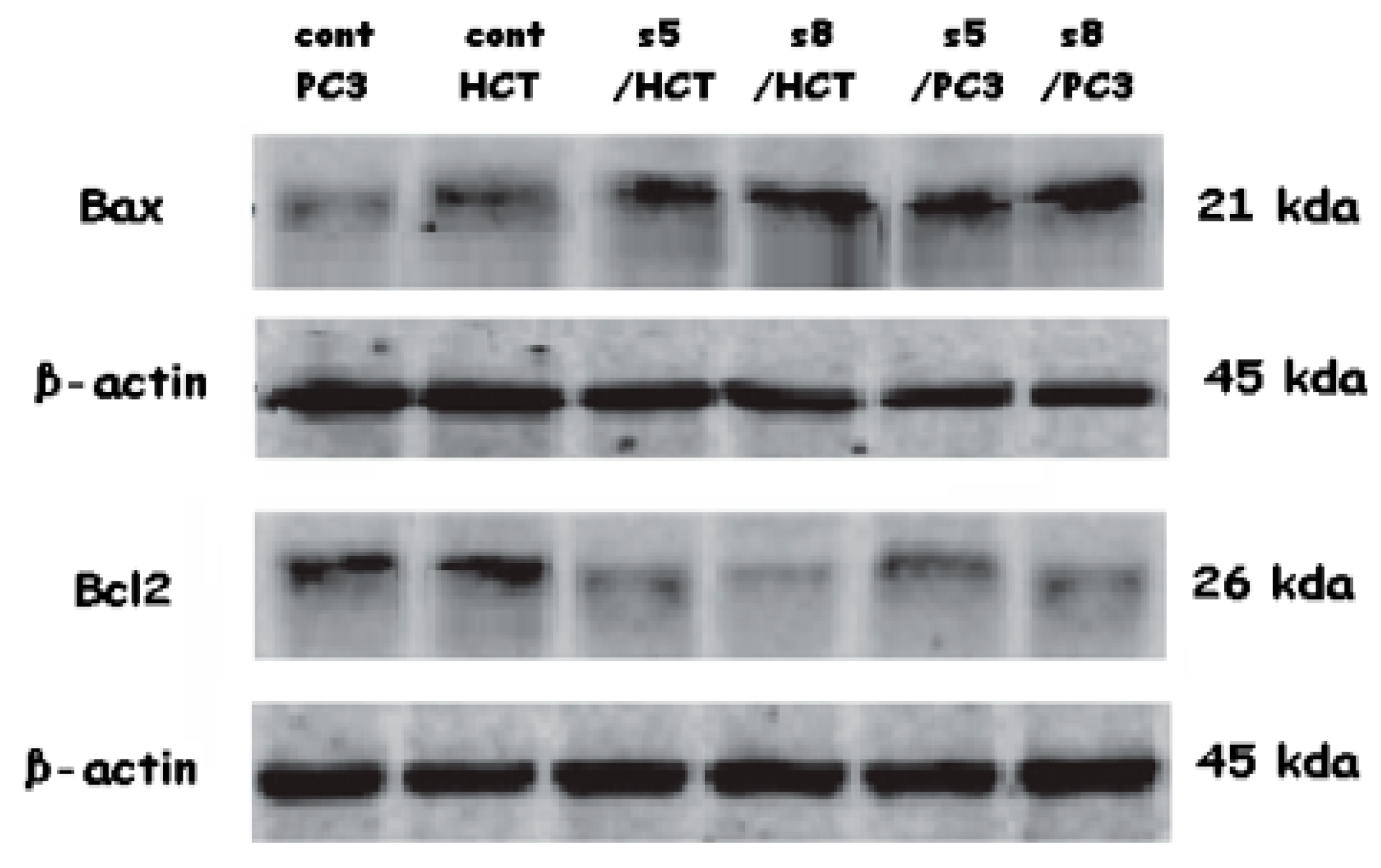
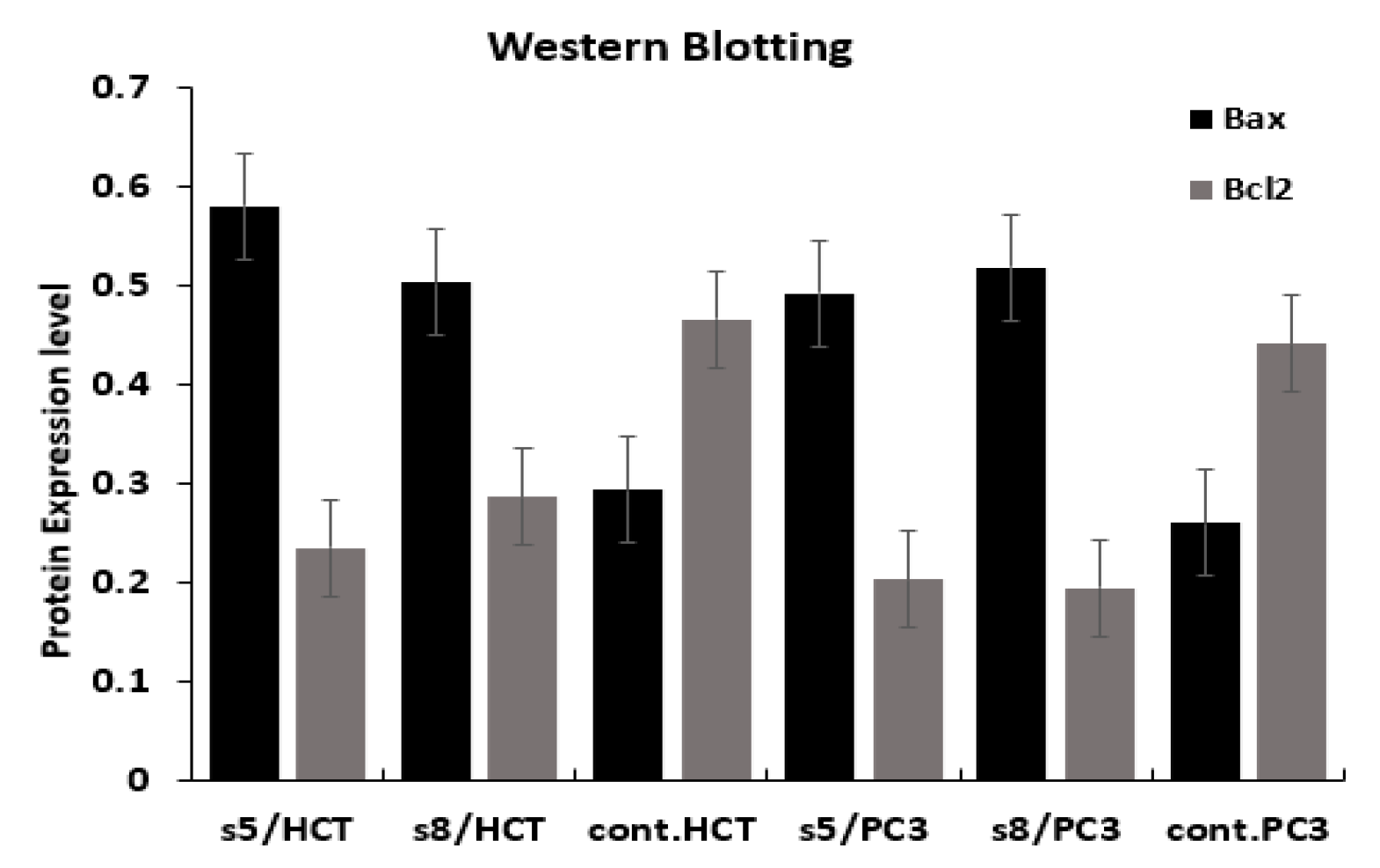
2.11. Western Blotting Analysis
3. Results
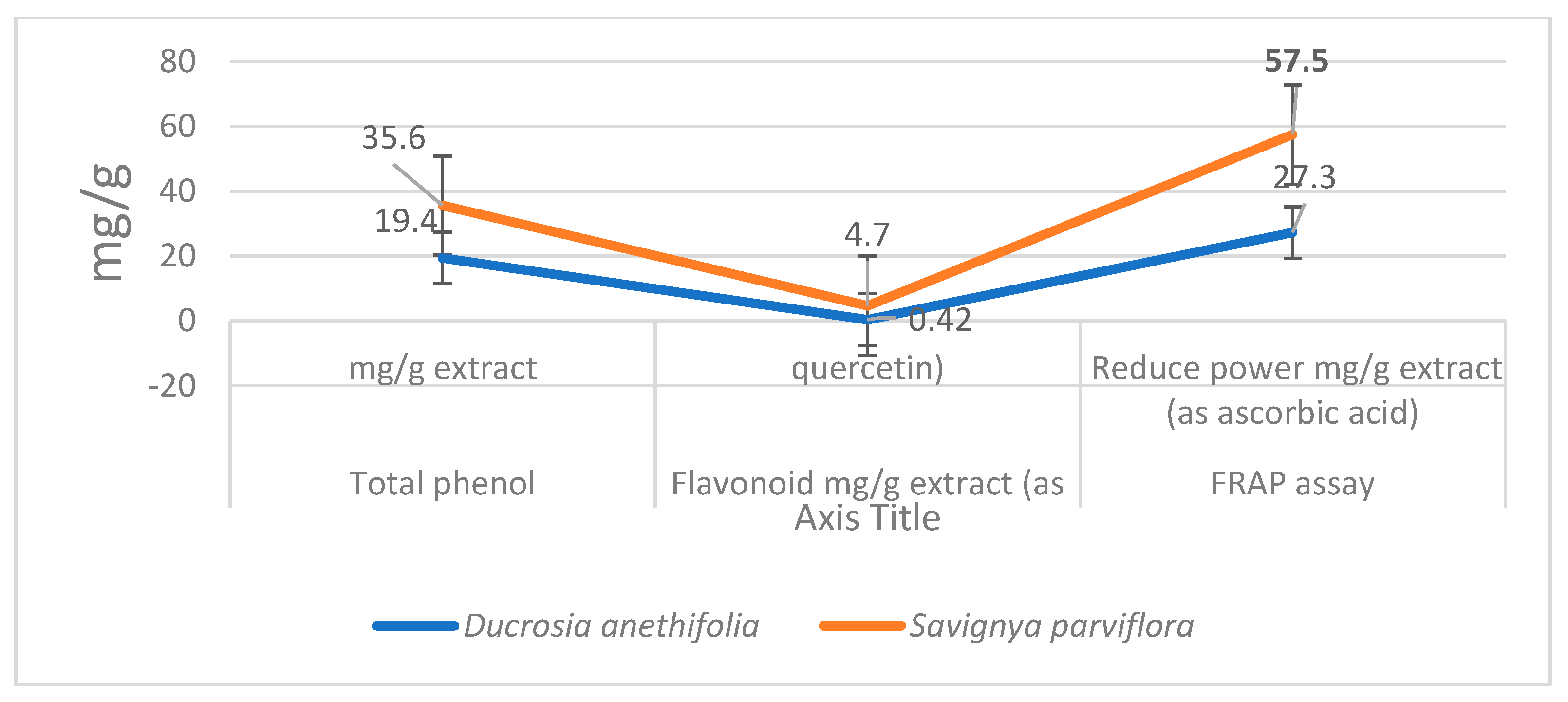
3.1. Analysis of the Total Content of Phenolics, Flavonoids and Antioxidant Effect of Plant Extracts
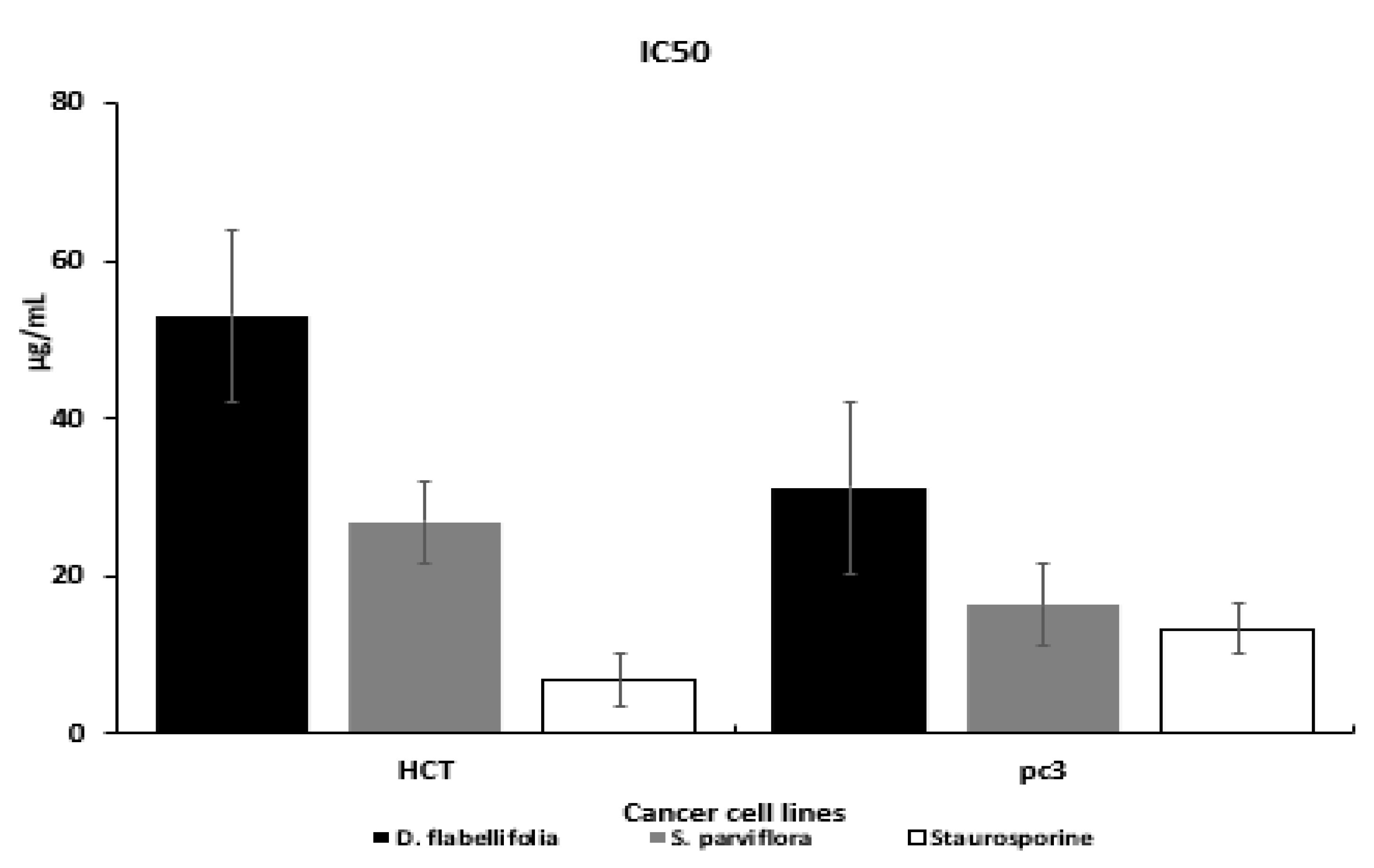
3.2. Cytotoxic Effects
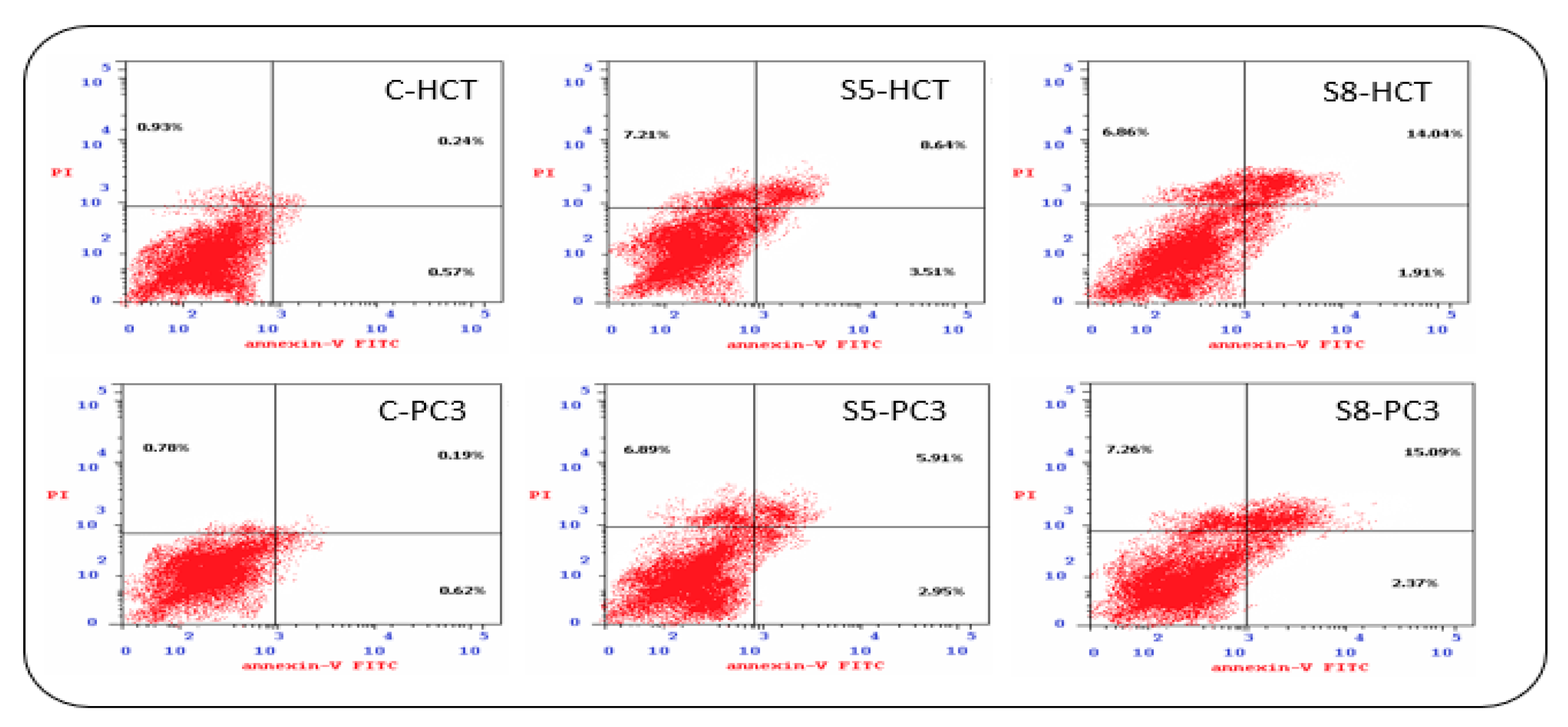
3.3. Apoptosis Detection
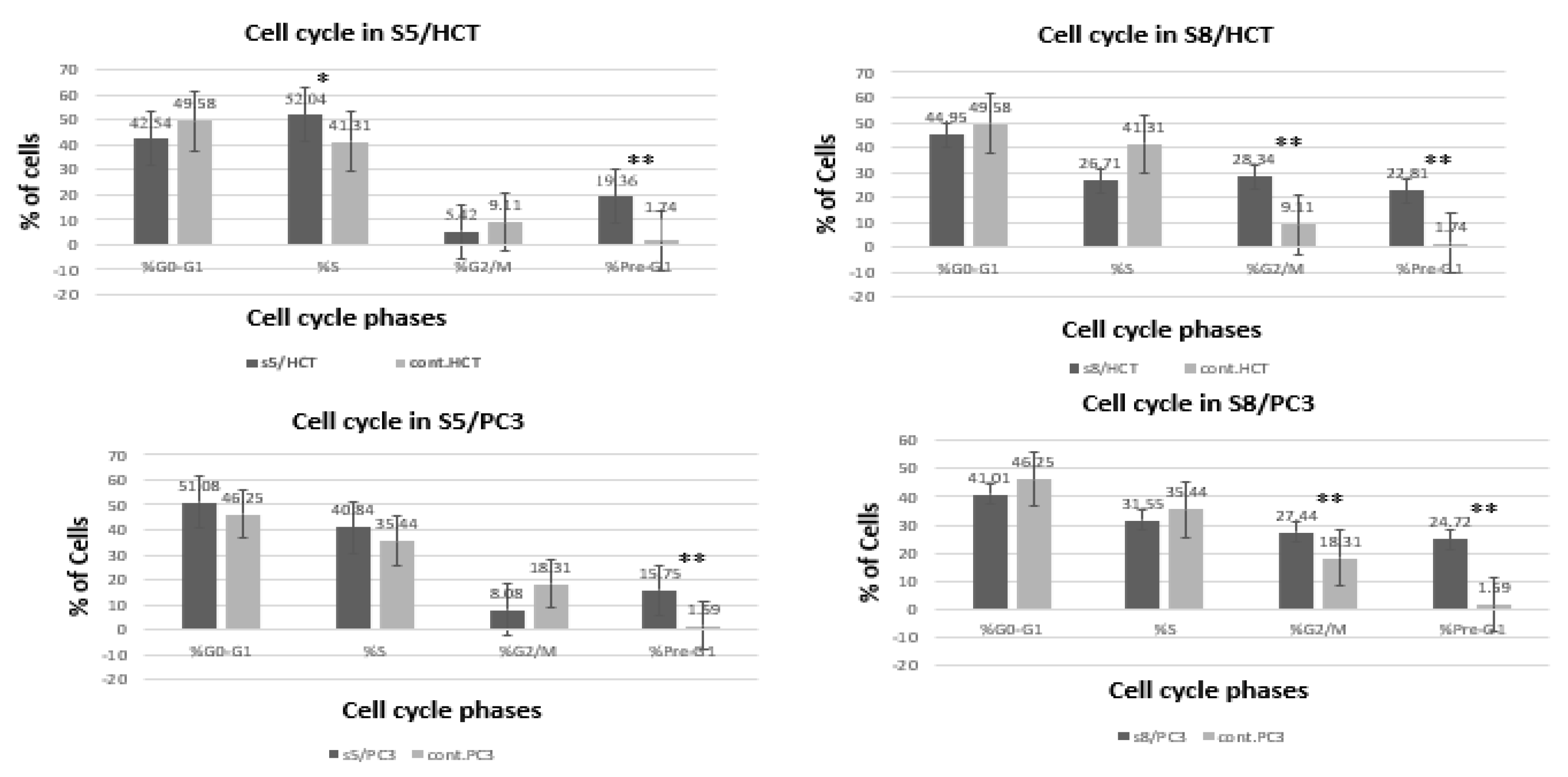
3.4. Cell Cycle Analysis
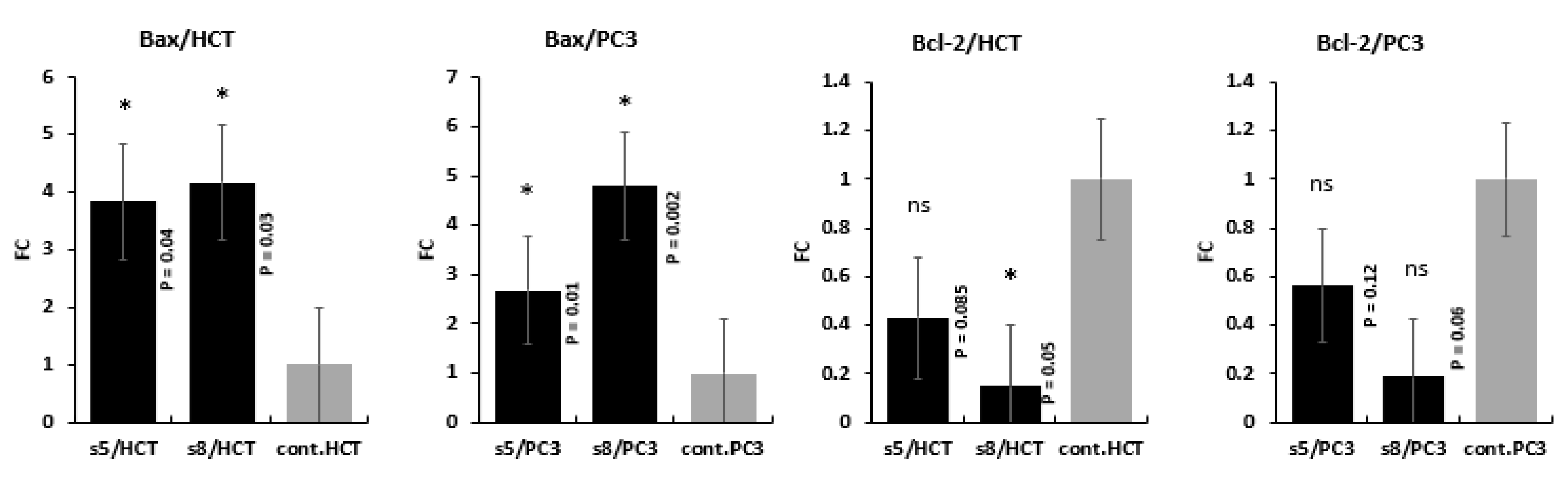
3.5. Gene Expression Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Twilley, D.; Rademan, S.; Lall, N. A review on traditionally used South African medicinal plants, their secondary metabolites and their potential development into anticancer agents. J. Ethnopharmacol. 2020, 261, 113101. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moot, A.; Polglase, A.; Giles, G.; Garson, M.; Thursfield, V.; Gunter, D. Men with colorectal cancer are predisposed to prostate cancer. ANZ J. Surg. 2003, 73, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Hugh, D.; Hetzel, J.T.; Roy, H.; Rubin, D.T. Association of colorectal cancer and prostate cancer and impact of radiation therapy. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1979–1985. [Google Scholar]
- Dhanasekaran, S. Phytochemical characteristic s of aerial part of Cissus quadrangularis (L.) and it’s in vitro inhibitory activity against leukemic cells and antioxidant properties. Saudi J. Biol. Sci. 2020, 27, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.F.; Saleem, A.; Rasul, A.; Baig, M.M.F.A.; Bin-Jumah, M.; AbdelDaim, M.M. Anticancer natural medicines: An overview of cell signaling and other targets of anticancer phytochemicals. Eur. J. Pharmacol. 2020, 888, 173488. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural Products as Sources of New Drugs over the Period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef]
- Al-Harbi, N.A.; Awad, N.S.; Alsberi, H.M.; Abdein, M.A. Apoptosis Induction, Cell Cycle Arrest and in vitro Anticancer Potentiality of Convolvulus spicatus and Astragalus vogelii. World J. Environ. Biosci. 2014, 8, 69–75. [Google Scholar]
- Hajhashemi, V.; Rabbani, M.; Ghanadi, A.; Davari, E. Evaluation of anti-anxiaty and sedative effects of essential oil of Ducrossia anethifolia in mice. Clinics 2010, 10, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Karimi, M.; Ebrahimi, A.; Sahraroo, A.; Moosavi, S.A.; Moosavi, F.; Bihamta, M.R. Callus formation and shoot organogenesis in Moshgak (Ducrosia flabellifolia Boiss.) from cotyledon. J. Food. Agric. Environ. 2009, 7, 441–445. [Google Scholar]
- Talib, W.H.; Issa, R.A.; Kherissat, F.; Mahasneh, A.M. Jordanian Ducrosia flabellifolia Inhibits Proliferation of Breast Cancer Cells by Inducing Apoptosis. Br. J. Med. Med. Res. 2013, 3, 771–783. [Google Scholar] [CrossRef]
- Queslati, M.H.; Bouajila, J.; Belkacem, M.A.; Harrath, A.H.; Alwasel, S.H.; Ben Jannet, H. Cytotoxicity of new secondary metabolites, fatty acids and tocols composition of seeds of Ducrosia anethifolia (DC.) Boiss. Nat. Prod. Res. 2017, 6419, 1–7. [Google Scholar]
- Luo, K.W.; Sun, J.G.; Chan, J.Y.W.; Yang, L.; Wu, S.H.; Fung, K.P.; Liu, F.Y. Anticancer effects of imperatorin isolated from Angelica dahurica: Induction of apoptosis in HepG2 cells through both death-receptor- and mitochondria-mediated pathways. Chemotherapy 2011, 57, 449–459. [Google Scholar] [CrossRef]
- Nermien, Z.; Ahmed, A.; Farid, O.A. Assessment of In vitro and In vivo Bioactivities Potential of Some Egyptian Wild Plants as Anti-Ulcerogenic Effect on male albino rats. Pharm. Chem. J. 2017, 4, 68–86. [Google Scholar]
- Manshood, M.A.; Al-Halbosiy, M.F.M.; Radhwan, M.M. In Vitro Cytotoxic Activity of Some Wild Plants Extracts Against Raw264.7 Cell Line. Plant Arch. 2019, 19, 3983–3986. [Google Scholar]
- Potikanond, S.; Sookkhee, S.; Na Takuathung, M.; Mungkomasawakul, P.; Wikan, N.; Smith, D.R.; Nimlamool, W. Kaempferia parviflora Extract Exhibits Anti-Cancer Activity against HeLa Cervical Cancer Cells. Front. Pharmacol. 2017, 8, 630. [Google Scholar] [CrossRef] [Green Version]
- Paramee, S.; Sookkhee, S.; Sakonwasun, C.; Na Takuathung, M.; Mungkomasawakul, P.; Nimlamool, W.; Potikanond, S. Anti-cancer effects of Kaempferia parviflora on ovarian cancer SKOV3 cells. BMC Complement. Altern. Med. 2018, 18, 178–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koldas, S.; Demirtas, I.; Ozen, T.; Demircia, M.; Behçet, L. Phytochemical screening, anticancer and antioxidant activities of Origanum vulgare L. ssp. Viride (Boiss.) Hayek, a plant of traditional usage. J. Sci. Food Agric. 2015, 95, 786–798. [Google Scholar] [CrossRef]
- Singleton, L.; Rossi, A. Colorimetry of total phenolics with phosphomolybdic–phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Chang, C.; Yang, H.; Wen, M.; Chern, C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Oyaizu, M. Antioxidative Activities of Products of Browning Reaction Prepared from Glucose amine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Puranik, S.; Murigendra, B.; Hiremath, H.; Nerli, R.; Ghagane, S. Evaluation of in vitro Antioxidant and Anticancer Activity of Tabernaemontana divaricata Leaf Extracts Against T-24 Human Bladder Cancer Cell Lines. Int. J. Cancer Res. 2018, 14, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Mielanczyk, A.; Mrowiecb, K.; Kupczaka, M.; Mielanczykc, Ł.; Scieglinskab, D.; Gogler-Piglowskab, A.; Michalskic, M.; Gabrielc, A.; Neugebauera, D.; Skonieczna, M. Synthesis and in vitro cytotoxicity evaluation of star-shaped polymethacrylic conjugates with methotrexate or acitretin as potential antipsoriatic prodrugs. Eur. J. Pharmacol. 2020, 866, 172804. [Google Scholar] [CrossRef]
- Suzuki, K.; Kazui, T.; Yoshida, M.; Uno, T.; Kobayashi, T. Drug-induced apoptosis and p53, bcl2 and bax expression in breast cancer tissues in vivo and in fibroblast cells in vitro. Jpn. J. Clin. Oncol. 1999, 29, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Burnette, W.N. Western Blotting: Electrophoretic Transfer of Proteins from Sodium Dodecyl Sulfate Polyacrylamide Gels to Unmodified Nitrocellulose and Radiographic Detection with Antibody and Radioiodinated Protein A. Annal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1989. [Google Scholar]
- Liu, S.; Khan, A.; Yang, X.; Dong, B.; Ji, J.; Zhai, G. The reversal of chemotherapy-induced multidrug resistance by nanomedicine for cancer therapy. J. Control. Release 2021, 335, 1–2. [Google Scholar] [CrossRef]
- Khan, T.; Ali, M.; Khan, A.; Nisar, P.; Jan, S.; Afridi, S.; Khan Shinwari, Z. Anticancer Plants: A Review of the Active Phytochemicals, Applications in Animal Models, and Regulatory Aspects. Biomolecules 2020, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Silva, A.; Tavares-Carreón, F.; Figueroa, M.; De la Torre-Zavala, S.; Gastelum-Arellanez, A.; Rodríguez-García, A.; Galán-Wong, L.; Avilés-Arnaut, H. Anticancer potential of the vetiaperuviana fruit methanolic extract. BMC Complement. Altern. Med. 2017, 17, 241. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- Bendary, E.; Francis, R.R.; Ali, H.M.G.; Sarwat, M.I.; El Hady, S. Antioxidant and structure–activity relationships (SARs) of some phenolic and anilines compounds. Ann. Agric. Sci. 2013, 58, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Côté, J.; Caillet, S.; Doyon, G.; Sylvain, J.F.; Lacroix, M. Bioactive compounds in cranberries and their biological properties. Crit. Rev. Food Sci. Nutr. 2010, 50, 666–679. [Google Scholar] [CrossRef]
- Oberoi, H.S.; Sandhu, S.K. Therapeutic and Nutraceutical Potential of Bioactive Compounds Extracted from Fruit Residues AU—Babbar, Neha. Crit. Rev. Food Sci. Nutr. 2015, 55, 319–337. [Google Scholar]
- Chikara, S.; Nagaprashantha, L.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef]
- Auyeung, B.; Taylor, K.; Hackett, G.; Baron-Cohen, S. Foetal testosterone and autistic traits in 18 to 24-month-old children. Mol. Autism 2010, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Priyadarsini, R.V.; Murugan, R.S.; Maitreyi, S.; Ramalingam, K.; Karunagaran, D.; Nagini, S. The flavonoid quercetin induces cell cycle arrest and mitochondria-mediated apoptosis in human cervical cancer (HeLa) cells through p53 induction and NF-κB inhibition. Eur. J. Pharmacol. 2010, 649, 84–91. [Google Scholar] [CrossRef]
- Anwar, A.; Uddin, N.; Siddiqui, B.S.; Siddiqui, R.A.; Begum, S.; Choudhary, M.I. A natural flavonoid lawsonaringenin induces cell cycle arrest and apoptosis in HT-29 colorectal cancer cells by targeting multiple signalling pathways. Mol. Biol. Rep. 2018, 45, 1339–1348. [Google Scholar] [CrossRef]
- Bishayee, A.; Darvesh, S. Pomegranate-derived constituents as inducers of cell death: Implications in cancer prevention and therapy. In Natural Compounds as Inducers of Cell Death; Diederich, M., Noworyta, K., Eds.; Springer: Berlin/Heidelberg, Germany; Amsterdam, The Netherlands, 2012; Volume 1, pp. 33–47. [Google Scholar]
- Mukhtar, E.; Adhami, V.M.; Khan, N.; Mukhtar, H. Apoptosis, and autophagy induction as mechanism of cancer prevention by naturally occurring dietary agents. Curr. Drug Targets 2012, 13, 1831–1841. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, D.; Mandal, A.; Nevo, E.; Bishayee, A. Apoptosis-inducing effects of extracts from desert plants in HepG2 human hepatocarcinoma cells. Asian Pac. J. Trop. Biomed. 2015, 5, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, N.; Nakamura, Y.; Ueda, Y.; Abe, M.; Ozawa, Y.; Uchida, K.; Osawa, T. Dietary Ginger constituents galangals A and B, are potent apoptosis inducers in human T lymphoma Jurkat T cells. Cancer Lett. 2003, 199, 113–119. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M.; Dikilitas, M. Role of antioxidant phytochemicals in prevention, formation, and treatment of cancer in Reactive Oxygen Species (ROS) in Living Cells. In Reactive Oxygen Species (ROS) in Living Cells; Cristiana, F., Elena, A., Eds.; InTech Open: Rijeka, Croatia, 2018. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.; Liebes, L.; Ng, B.; Buckley, M.; Elliott, J.; Adams, J.; Jiang, D.; Muggia, M.; Perez-Soler, R. PS-341, a novel proteasome inhibitor, induces Bcl-2 phosphorylation and cleavage in association with G2-M phase arrest and apoptosis. Mol. Cancer Ther. 2002, 1, 841–849. [Google Scholar] [PubMed]
- Thomas, A.; Giesler, T.; White, E. p53 mediates Bcl-2 phosphorylation and apoptosis via activation of the Cdc42/JNK1 pathway. Oncogene 2000, 19, 5259–5269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crescenzi, E.; Palumbo, G. Bcl-2 exerts a pRb-mediated cell cycle inhibitory function in HEC1B endometrial carcinoma cells. Gynecol. Oncol. 2001, 81, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Crescenzi, E.; Palumbo, G.; Brady, H.J. Bcl-2 activates a program of premature senescence in human carcinoma cells. Biochem. J. 2003, 15, 263–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Annealing Temperature | ||
|---|---|---|---|---|
| Forward | Reverse | |||
| 1 | Bcl-2 | 5′-CCTGTGGATGACTGAGTACC-3′ | 5′-GAGACAGCCAGGAGAAATCA-3′ | 55 °C |
| 2 | Bax | 5′-TTCCGAGTGGCAGCTGAGATGTTT-3′ | 5′-TGCTGGCAAAGTAGAAGAGGGCAA-3′ | 55 °C |
| 3 | β-actin | 5′-GTGACATCCACACCCAGAGG-3′ | 5′-ACAGGATGTCAAAACTGCCC-3′ | 58 °C |
| Concentration (µg/mL) | % of Viability | |||
|---|---|---|---|---|
| D. flabellifolia | S. parviflora | |||
| HCT-116 | PC3 | HCT-116 | PC3 | |
| 100 | 45 | 43 | 40 | 40 |
| 25 | 57 | 52 | 52 | 45 |
| 6.3 | 64 | 60 | 59 | 56 |
| 1.6 | 73 | 69 | 71 | 68 |
| 0.4 | 84 | 78 | 77 | 71 |
| No. | Sample | Cytotoxicity | |
|---|---|---|---|
| Code | IC50 µg/mL | ||
| Code | HCT | PC3 | |
| 1 | D. flabellifolia | 53.1 ± 2.98 | 31.3 ± 1.7 |
| 2 | S. parviflora | 26.9 ± 1.51 | 16.4 ± 0.89 |
| 3 | Staurosporine | 6.9 ± 0.39 | 13.4 ± 0.73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alghamdi, Y.S.; Saleh, O.M.; Alqadri, N.; Mashraqi, M.M.; Bahattab, O.; Awad, N.S. Effect of Ducrosia flabellifolia and Savignya parviflora Extracts on Inhibition of Human Colon and Prostate Cancer Cell Lines. Curr. Issues Mol. Biol. 2021, 43, 1518-1528. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030107
Alghamdi YS, Saleh OM, Alqadri N, Mashraqi MM, Bahattab O, Awad NS. Effect of Ducrosia flabellifolia and Savignya parviflora Extracts on Inhibition of Human Colon and Prostate Cancer Cell Lines. Current Issues in Molecular Biology. 2021; 43(3):1518-1528. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030107
Chicago/Turabian StyleAlghamdi, Youssef Saeed, Osama Moseilhy Saleh, Nada Alqadri, Mutaib Mosaued Mashraqi, Omar Bahattab, and Nabil Saied Awad. 2021. "Effect of Ducrosia flabellifolia and Savignya parviflora Extracts on Inhibition of Human Colon and Prostate Cancer Cell Lines" Current Issues in Molecular Biology 43, no. 3: 1518-1528. https://0-doi-org.brum.beds.ac.uk/10.3390/cimb43030107