
Tart Cherry (Fruit of Prunus cerasus) Concentrated Powder (TCcp) Ameliorates Glucocorticoid-Induced Muscular Atrophy in Mice

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample
2.2. Animal Husbandry and Experimental Design
2.3. Measurement of Body Weights
2.4. Measurement of Gastrocnemius Muscle and Calf Thickness
2.5. Measurement of Calf Muscle Strength
2.6. Measurement of Gastrocnemius Muscle Weight
2.7. Serum Biochemical Analysis
2.8. Antioxidant Analysis
2.9. qRT-PCR
2.10. Histopathology
2.11. Immunohistochemistry
2.12. Statistical Analyses
3. Results
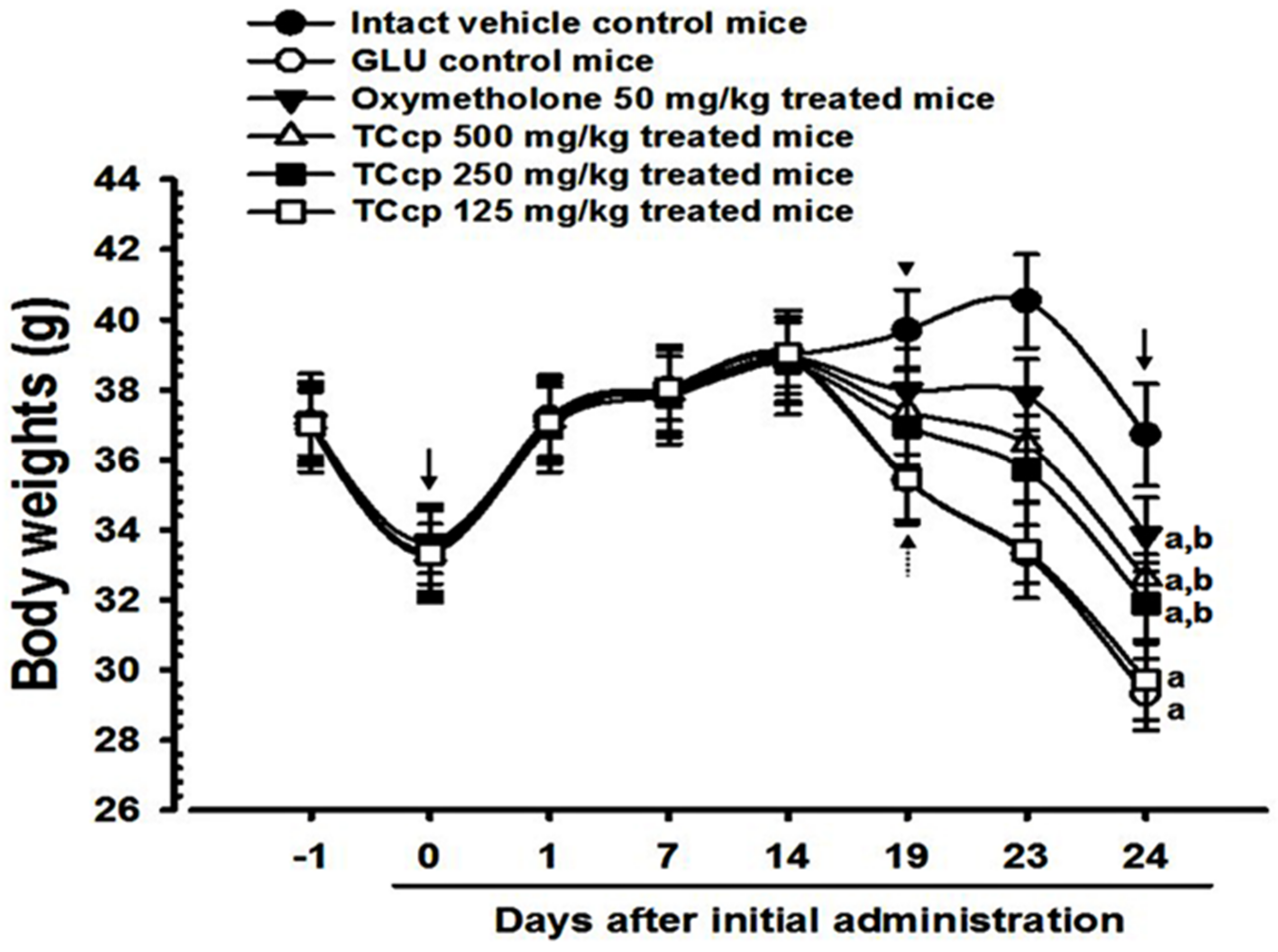
3.1. Body Weight Changes
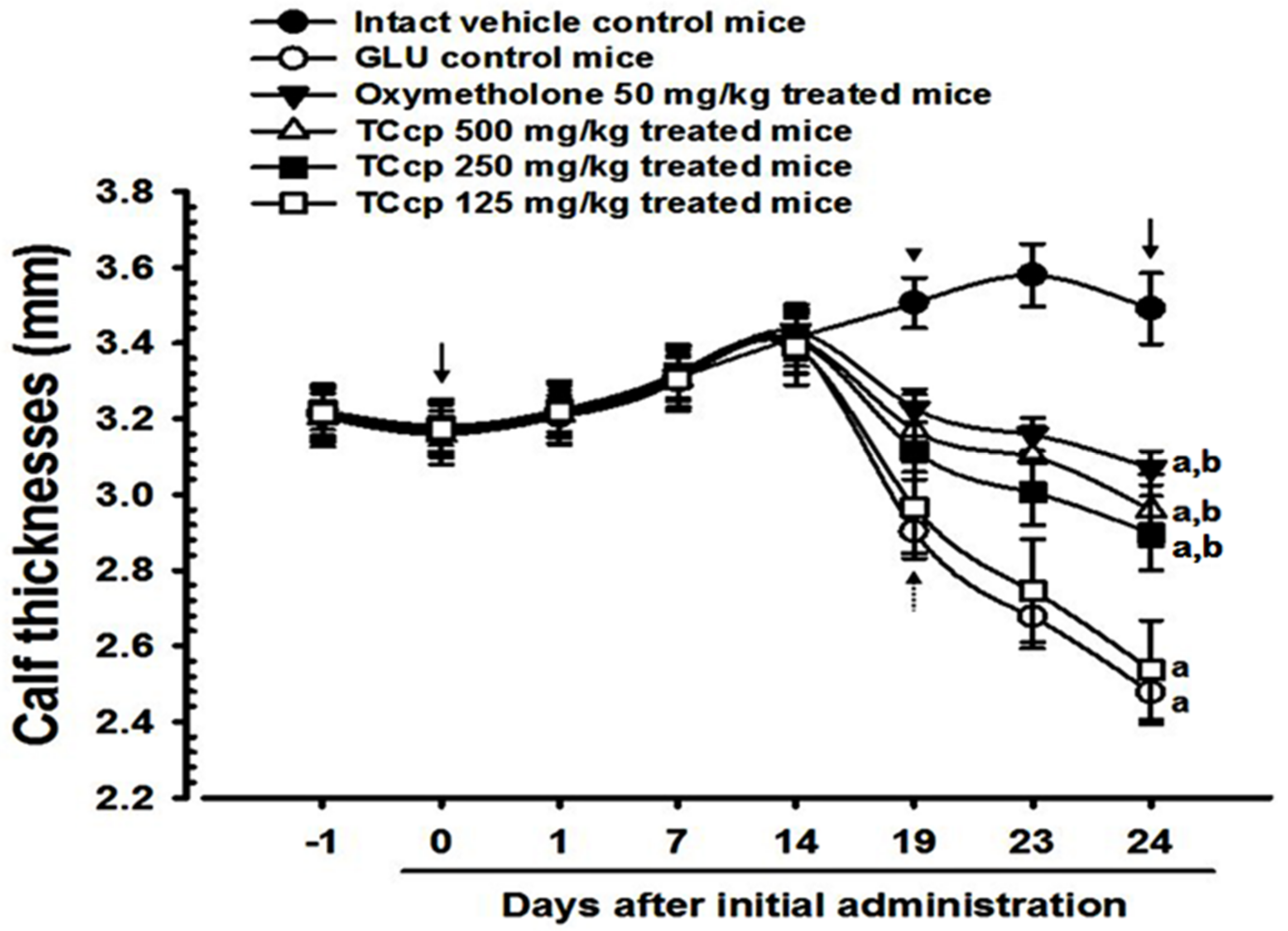
3.2. Changes in Calf Thickness
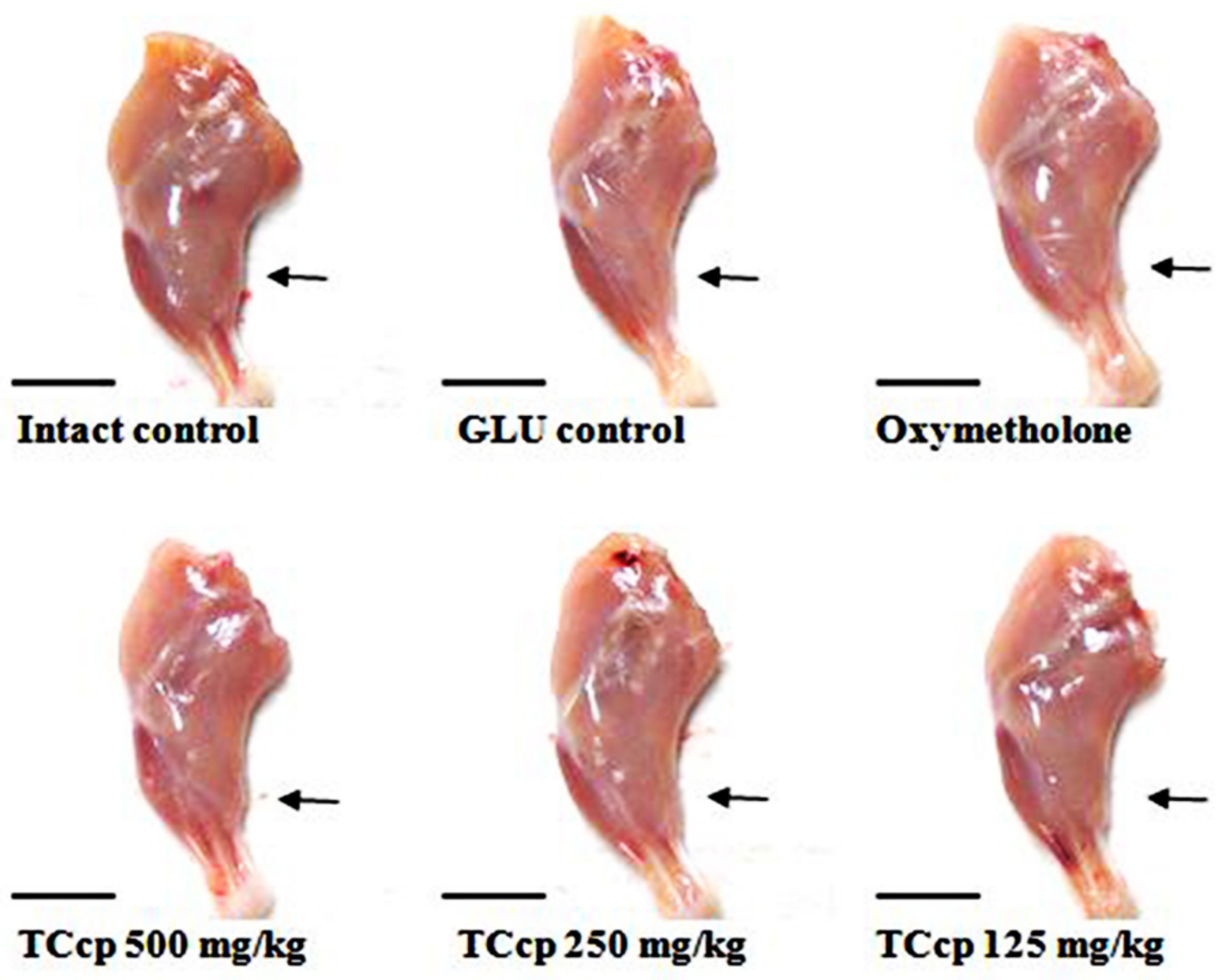
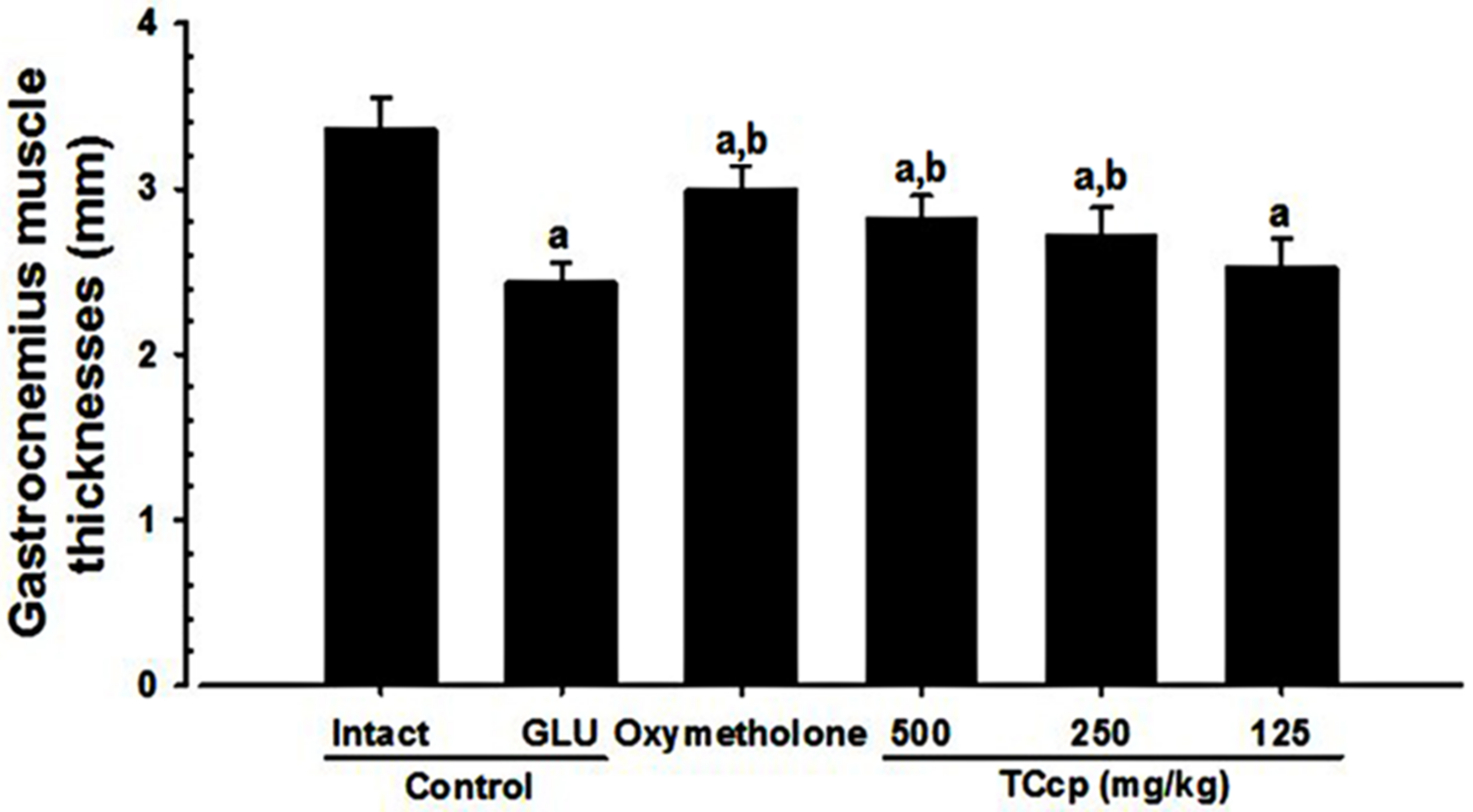
3.3. Effects on Gastrocnemius Muscle Thickness
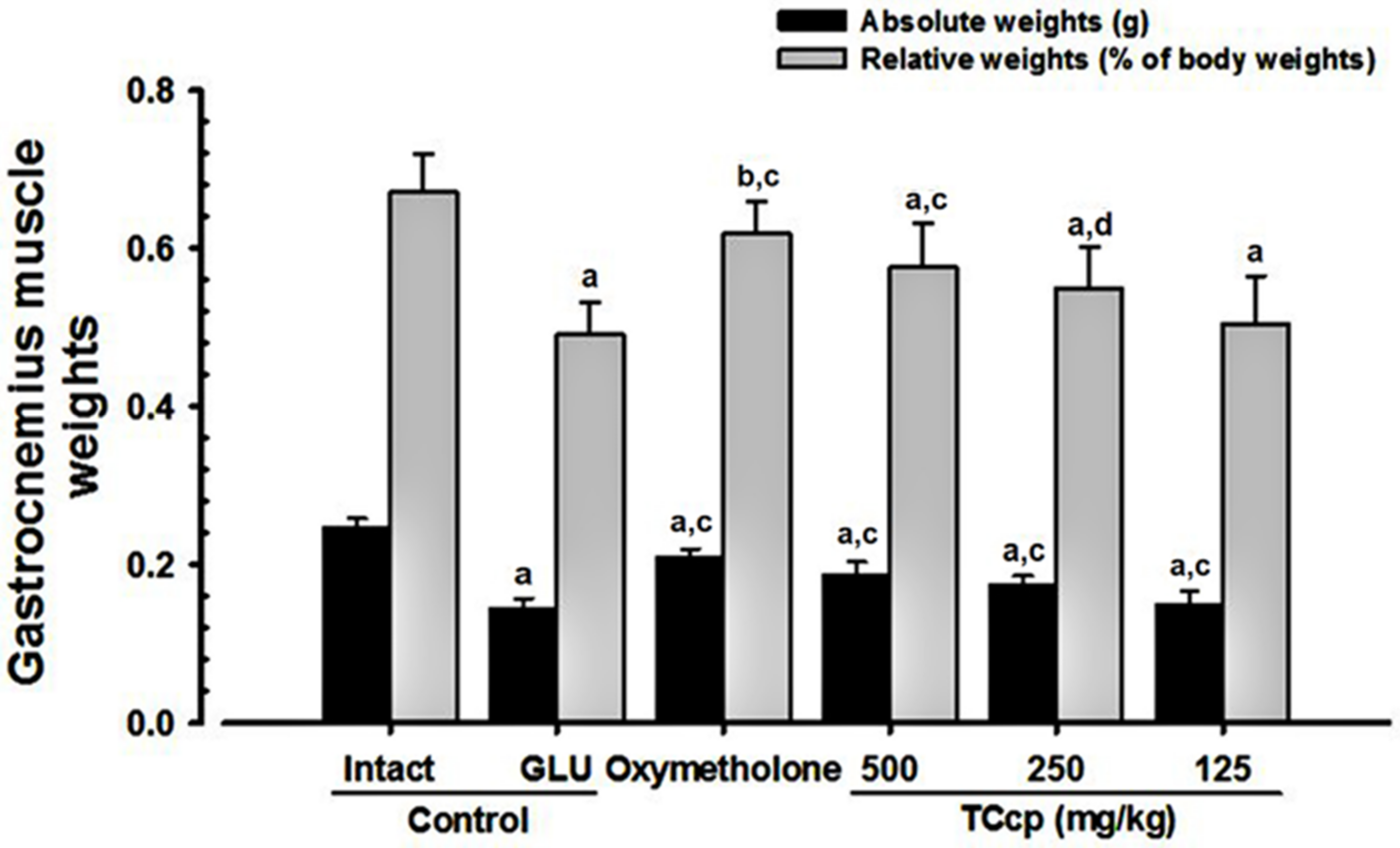
3.4. Changes in Weight of Gastrocnemius Muscle Mass
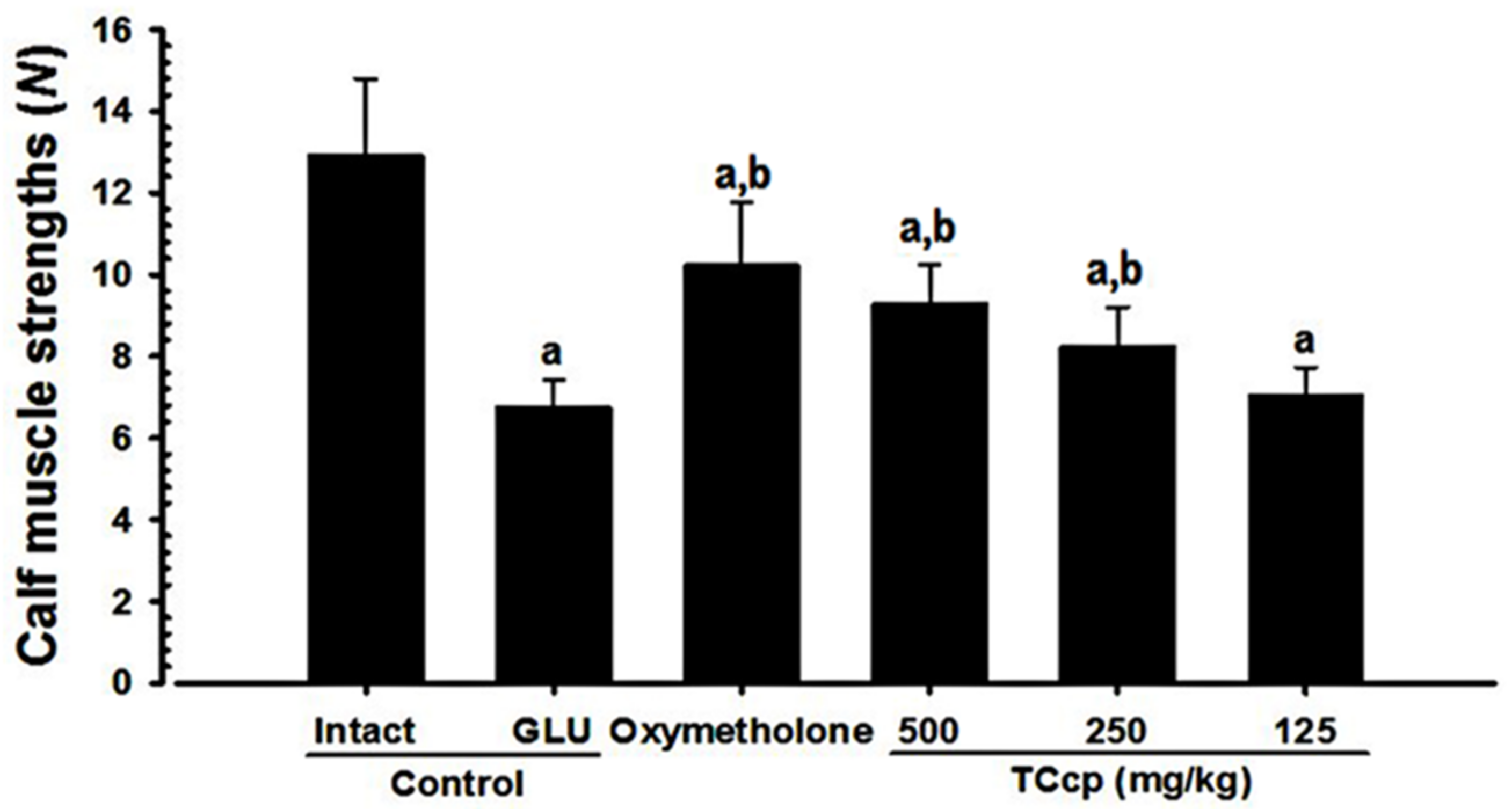
3.5. Changes in Calf Muscle Strength
3.6. Changes in Serum Biochemistry
3.7. Changes in Antioxidant Properties
3.7.1. Changes in Muscle MDA Levels and ROS Content
3.7.2. Changes in Muscle GSH Content and Enzyme Activities
3.8. Changes in Gastrocnemius Muscle Protein mRNA Expressions (qRT-PCR Analysis)
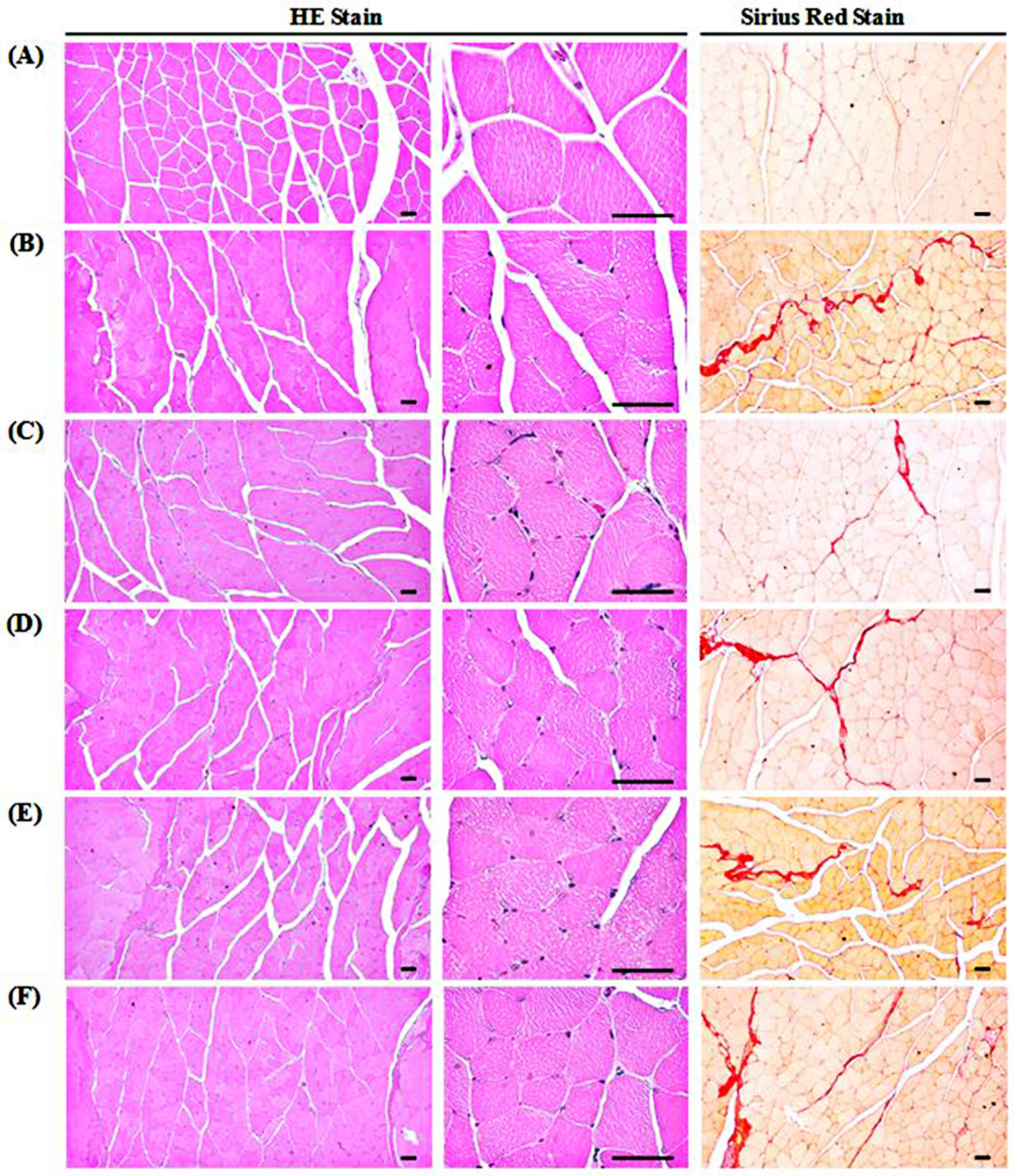
3.9. Changes in Gastrocnemius Muscle Histopathology
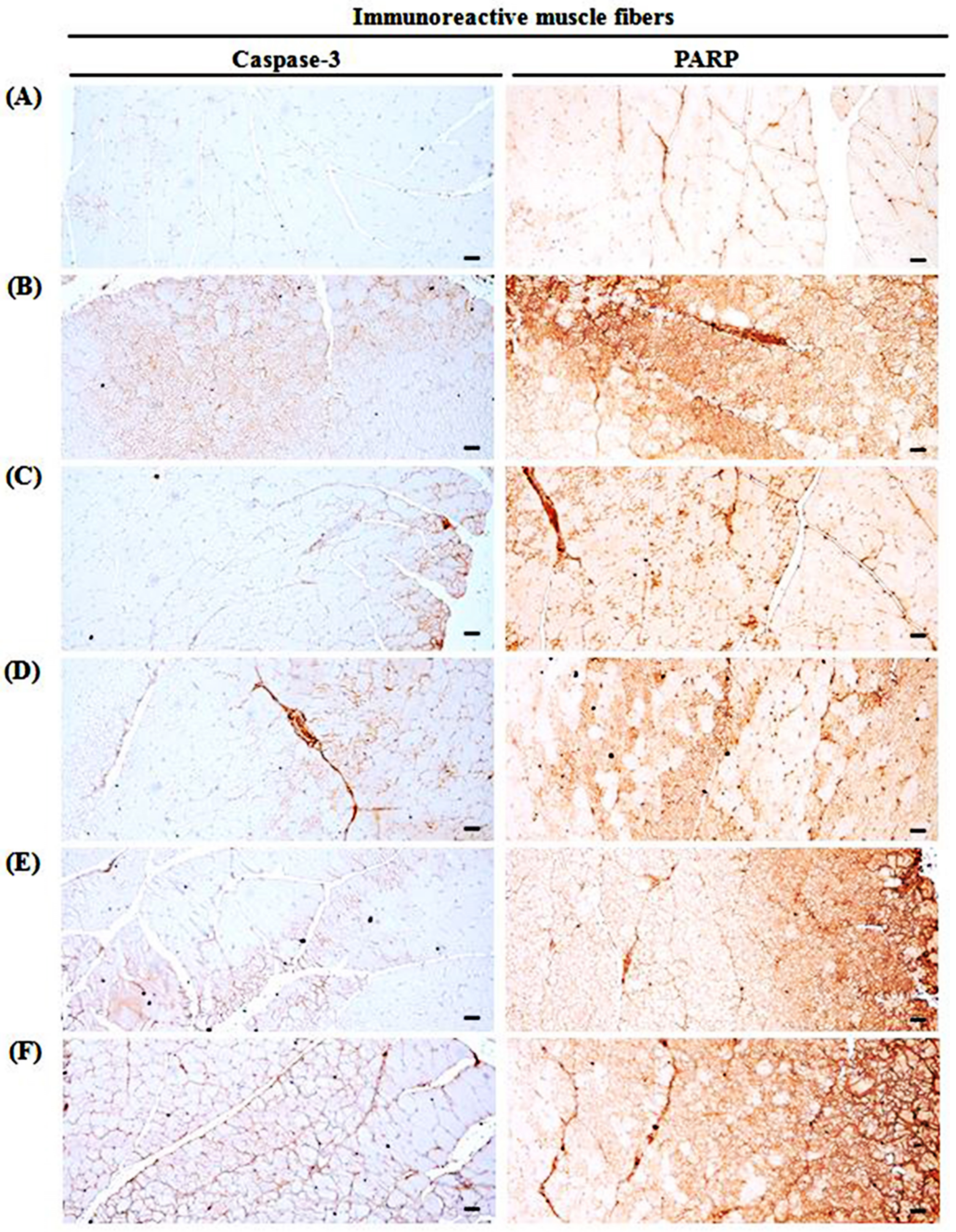
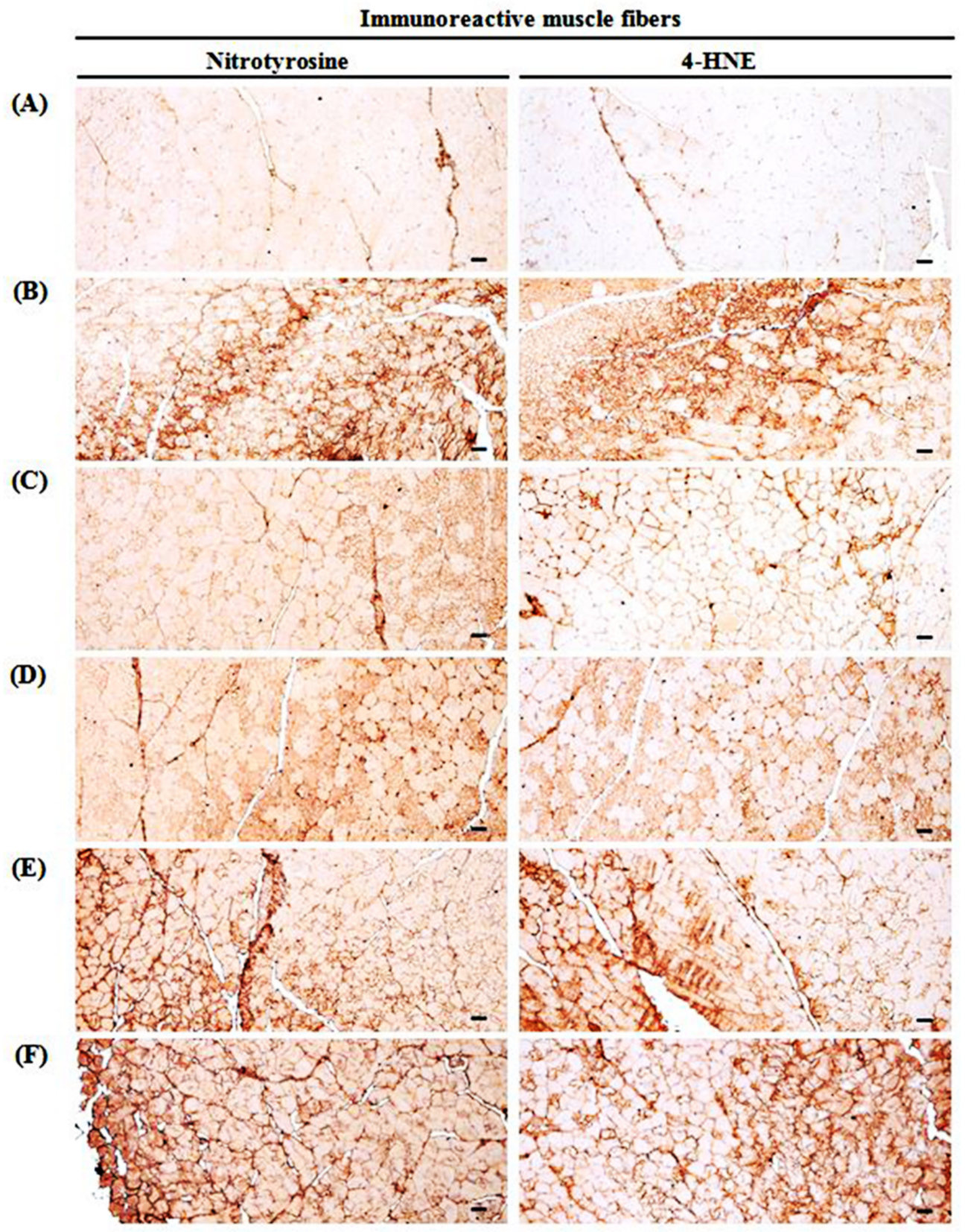
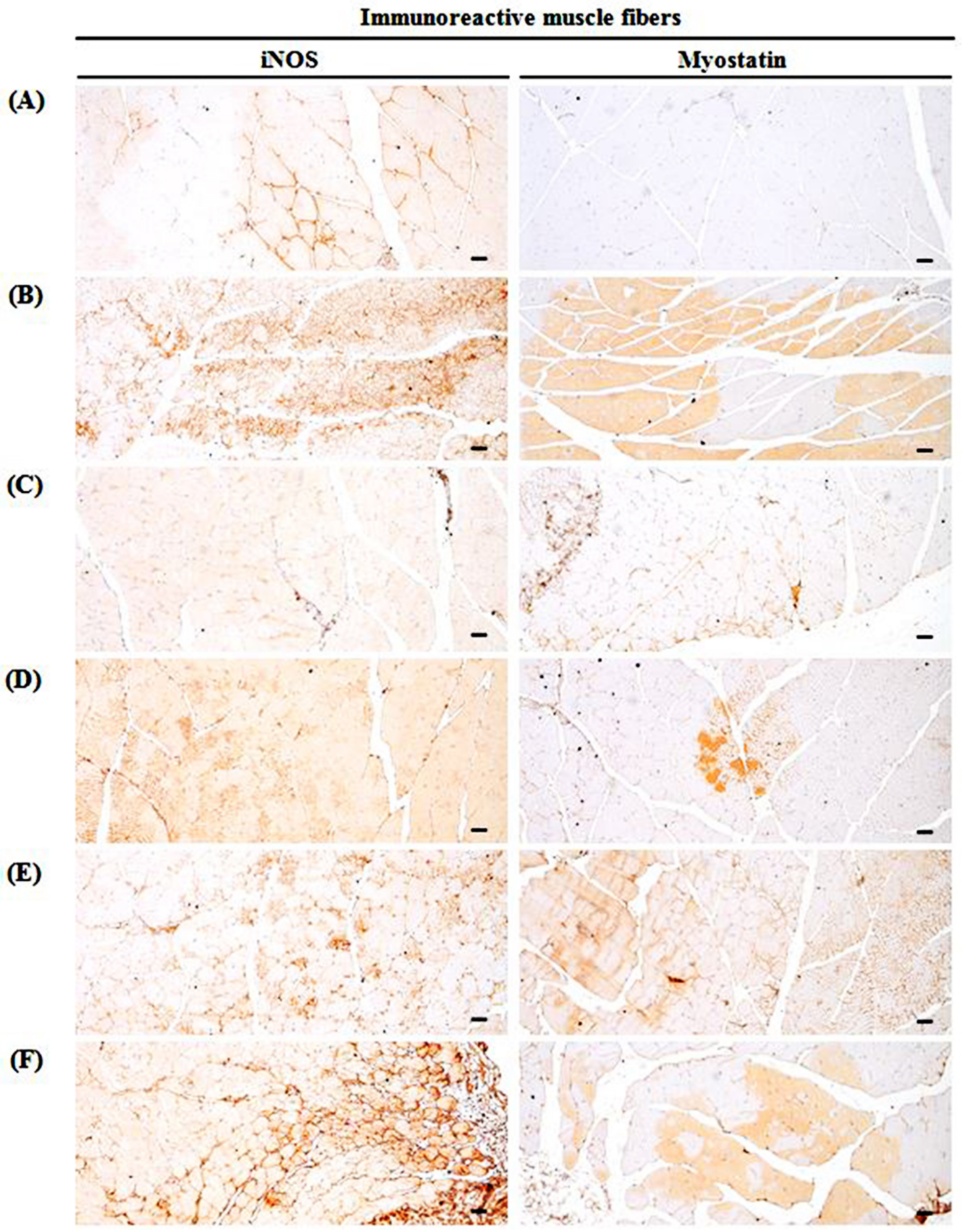
3.10. Changes in Immunohistochemistry of Gastrocnemius Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goodpaster, B.H.; Park, S.W.; Harris, T.B.; Kritchevsky, S.B.; Nevitt, M.; Schwartz, A.V.; Simonsick, E.M.; Tylavsky, F.A.; Visser, M.; Newman, A.B. The loss of skeletal muscle strength, mass, and quality in older adults: The health, aging and body composition study. J. Gerontol. A Biol. Sci. Med. 2006, 61, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Deschenes, M.R. Effects of Aging on Muscle Fibre Type and Size. Sports Med. 2004, 34, 809–824. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.B.; Kupelian, V.; Visser, M.; Simonsick, E.M.; Goodpaster, B.H.; Kritchevsky, S.B.; Tylavsky, F.A.; Rubin, S.M.; Harris, T.B. Strength, but not muscle mass, is associated with mortality in the health, aging and body composition study cohort. J. Gerontol. A Biol. Sci. Med. 2006, 61, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Solomon, A.M.; Bouloux, P.M.G. Modifying muscle mass—The endocrine perspective. J. Endocrinol. 2006, 191, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardozo, T.; Pagano, M. The SCF ubiquitin ligase: Insights into a molecular machine. Nat. Rev. Mol. Cell Biol. 2004, 5, 739–751. [Google Scholar] [CrossRef]
- Lecker, S.H.; Jagoe, R.T.; Gilbert, A.; Gomes, M.; Baracos, V.; Bailey, J.; Price, S.R.; Mitch, W.E.; Goldberg, A.L. Multiple types of skeletal muscle atrophy involve a common program of changes in gene expression. FASEB J. 2003, 18, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Egerman, M.A.; Glass, D.J. Signaling pathways controlling skeletal muscle mass. Crit. Rev. Biochem. Mol. Biol. 2013, 49, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Hortobágyi, T.; Dempsey, G.L.; Fraser, D.D.; Zheng, D.; Hamilton, G.; Lambert, J.; Dohm, G.L. Changes in muscle strength, muscle fibre size and myofibrillar gene expression after immobilization and retraining in humans. J. Physiol. 2000, 524, 293–304. [Google Scholar] [CrossRef] [Green Version]
- Reardon, T.F.; Allen, D.G. Iron injections in mice increase skeletal muscle iron content, induce oxidative stress and reduce exercise performance. Exp. Physiol. 2009, 94, 720–730. [Google Scholar] [CrossRef]
- Su, Z.; Robinson, A.; Hu, L.; Klein, J.D.; Hassounah, F.; Li, M.; Wang, H.; Cai, H.; Wang, X.H. Acupuncture plus Low-Frequency Electrical Stimulation (Acu-LFES) Attenuates Diabetic Myopathy by Enhancing Muscle Regeneration. PLoS ONE 2015, 10, e0134511. [Google Scholar] [CrossRef]
- Russell, A.P. Molecular regulation of skeletal muscle mass. Clin. Exp. Pharmacol. Physiol. 2010, 37, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.R. Loss of skeletal muscle mass in aging: Examining the relationship of starvation, sarcopenia and cachexia. Clin. Nutr. 2007, 26, 389–399. [Google Scholar] [CrossRef]
- Kawano, F.; Tanihata, J.; Sato, S.; Nomura, S.; Shiraishi, A.; Tachiyashiki, K.; Imaizumi, K. Effects of dexamethasone on the expression of beta1–, beta2– and beta3–adrenoceptor mRNAs in skeletal and left ventricle muscles in rats. J. Physiol. 2009, 59, 383–390. [Google Scholar]
- Schakman, O.; Kalista, S.; Barbé, C.; Loumaye, A.; Thissen, J. Glucocorticoid-induced skeletal muscle atrophy. Int. J. Biochem. Cell Biol. 2013, 45, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Singleton, J.; Feldman, E.L. Insulin-like Growth Factor-I in Muscle Metabolism and Myotherapies. Neurobiol. Dis. 2001, 8, 541–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.; Vernus, B.; Chelh, I.; Cassar-Malek, I.; Gabillard, J.C.; Sassi, A.H.; Seiliez, I.; Picard, B.; Bonnieu, A. Myostatin and the skeletal muscle atrophy and hypertrophy signaling pathways. Cell. Mol. Life Sci. 2014, 71, 4361–4371. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Mallidis, C.; Bhasin, S.; Mahabadi, V.; Artaza, J.; Gonzalez-Cadavid, N.; Arias, J.; Salehian, B. Glucocorticoid-induced skeletal muscle atrophy is associated with upregulation of myostatin gene expression. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E363–E371. [Google Scholar] [CrossRef] [PubMed]
- Komamura, K.; Shirotani-Ikejima, H.; Tatsumi, R.; Tsujita-Kuroda, Y.; Kitakaze, M.; Miyatake, K.; Sunagawa, K.; Miyata, T. Differential gene expression in the rat skeletal and heart muscle in glucocorticoid-induced myopathy: Analysis by microarray. Cardiovasc. Drugs Ther. 2003, 17, 303–310. [Google Scholar] [CrossRef]
- Schakman, O.; Gilson, H.; De Coninck, V.; Lause, P.; Verniers, J.; Havaux, X.; Ketelslegers, J.M.; Thissen, J.P. Insulin-Like Growth Factor-I Gene Transfer by Electroporation Prevents Skeletal Muscle Atrophy in Glucocorticoid-Treated Rats. Endocrinology 2005, 146, 1789–1797. [Google Scholar] [CrossRef] [Green Version]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF–β superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Syverud, B.C.; VanDusen, K.W.; Larkin, L.M. Effects of Dexamethasone on Satellite Cells and Tissue Engineered Skeletal Muscle Units. Tissue Eng. Part A 2016, 22, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Sheng, L.; Wan, B.; Feng, P.; Sun, J.; Rigo, F.; Bennett, C.F.; Akerman, M.; Krainer, A.R.; Hua, Y. Downregulation of Survivin contributes to cell-cycle arrest during postnatal cardiac development in a severe spinal muscular atrophy mouse model. Hum. Mol. Genet. 2018, 27, 486–498. [Google Scholar] [CrossRef]
- Hoppstädter, J.; Perez, J.V.V.; Linnenberger, R.; Dahlem, C.; Legroux, T.M.; Hecksteden, A.; Tse, W.K.F.; Flamini, S.; Andreas, A.; Herrmann, J.; et al. The glucocorticoid-induced leucine zipper mediates statin-induced muscle damage. FASEB J. 2020, 34, 4684–4701. [Google Scholar] [CrossRef] [Green Version]
- Kweon, M.; Lee, H.; Park, C.; Choi, Y.H.; Ryu, J.H. A chalcone from Ashitaba (Angelica keiskei) stimulates myoblast differen-tiation and inhibits dexamethasone–induced muscle atrophy. Nutrients 2019, 11, 2419. [Google Scholar] [CrossRef] [Green Version]
- Pavlatos, A.M.; Fultz, O.; Monberg, M.J.; Vootkur, A. Review of oxymetholone: A 17α–alkylated anabolic–androgenic steroid. Clin. Ther. 2001, 23, 789–801. [Google Scholar] [CrossRef]
- Dorfman, R.I.; Kincl, F.A. Relative Potency of Various Steroids in an Anabolic-Androgenic Assay Using the Castrated Rat. Endocrinology 1963, 72, 259–266. [Google Scholar] [CrossRef]
- Aramwit, P.; Palapinyo, S.; Wiwatniwong, S.; Supasyndh, O. The efficacy of oxymetholone in combination with erythropoietin on hematologic parameters and muscle mass in CAPD patients. Int. J. Clin. Pharmacol. Ther. 2010, 48, 803–813. [Google Scholar] [CrossRef]
- Young, G.P.; Bhathal, P.S.; Sullivan, J.R.; Wall, J.A.J.; Fone, D.J.; Hurley, T.H. Fatal Hepatic Coma Complicating Oxymetholone Therapy in Multiple Myeloma. Aust. N. Z. J. Med. 1977, 7, 47–51. [Google Scholar] [CrossRef]
- Wood, P.; Liu, Y.J.A. Oxymetholone hepatotoxicity enhanced by concomitant use of cyclosporin A in a bone marrow transplant patient. Clin. Lab. Haematol. 1994, 16, 201–204. [Google Scholar] [CrossRef]
- Chang, S.; Münster, A.M.B.; Gram, J.; Sidelmann, J.J. Anabolic Androgenic Steroid Abuse: The Effects on Thrombosis Risk, Coagulation, and Fibrinolysis. In Seminars in Thrombosis and Hemostasis; Thieme Medical Publishers: New York, NY, USA, 2018; pp. 734–746. [Google Scholar]
- Wang, H.; Nair, M.G.; Strasburg, G.M.; Chang, Y.-C.; Booren, A.M.; Gray, J.I.; DeWitt, D.L. Antioxidant and Antiinflammatory Activities of Anthocyanins and Their Aglycon, Cyanidin, from Tart Cherries. J. Nat. Prod. 1999, 62, 294–296. [Google Scholar] [CrossRef]
- Seeram, N.P.; Momin, R.A.; Nair, M.G.; Bourquin, L.D. Cyclooxygenase inhibitory and antioxidant cyanidin glycosides in cherries and berries. Phytomedicine 2001, 8, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-O.; Heo, H.J.; Kim, Y.J.; Yang, H.S.; Lee, C.Y. Sweet and Sour Cherry Phenolics and Their Protective Effects on Neuronal Cells. J. Agric. Food Chem. 2005, 53, 9921–9927. [Google Scholar] [CrossRef] [PubMed]
- Kirakosyan, A.; Seymour, E.; Llanes, D.E.U.; Kaufman, P.B.; Bolling, S.F. Chemical profile and antioxidant capacities of tart cherry products. Food Chem. 2009, 115, 20–25. [Google Scholar] [CrossRef]
- Bell, P.G.; Gaze, D.C.; Davison, G.W.; George, T.W.; Scotter, M.J.; Howatson, G. Tart Montmorency cherry (Prunus cerasus L.) concentrate lowers uric acid, independent of plasma cyanidin–3–O–glucosiderutinoside. J. Funct. Foods 2014, 11, 82–90. [Google Scholar] [CrossRef]
- Seymour, E.M.; Warber, S.M.; Kirakosyan, A.; Noon, K.R.; Gillespie, B.; Uhley, V.E.; Wunder, J.; Urcuyo, D.E.; Kaufman, P.B.; Bolling, S.F. Anthocyanin pharmacokinetics and dose-dependent plasma antioxidant pharmacodynamics following whole tart cherry intake in healthy humans. J. Funct. Foods 2014, 11, 509–516. [Google Scholar] [CrossRef]
- Keane, K.M.; Bell, P.G.; Lodge, J.K.; Constantinou, C.L.; Jenkinson, S.E.; Bass, R.; Howatson, G. Phytochemical uptake following human consumption of Montmorency tart cherry (L. Prunus cerasus) and influence of phenolic acids on vascular smooth muscle cells in vitro. Eur. J. Nutr. 2015, 55, 1695–1705. [Google Scholar] [CrossRef] [Green Version]
- Howatson, G.; McHugh, M.P.; Hill, J.A.; Brouner, J.; Jewell, A.P.; Van Someren, K.A.; Shave, R.E.; Howatson, S.A. Influence of tart cherry juice on indices of recovery following marathon running. Scand. J. Med. Sci. Sports 2009, 20, 843–852. [Google Scholar] [CrossRef]
- Bell, P.G.; Walshe, I.H.; Davison, G.W.; Stevenson, E.; Howatson, G. Montmorency Cherries Reduce the Oxidative Stress and Inflammatory Responses to Repeated Days High-Intensity Stochastic Cycling. Nutrients 2014, 6, 829–843. [Google Scholar] [CrossRef] [Green Version]
- Keane, K.M.; George, T.W.; Constantinou, C.L.; Brown, M.A.; Clifford, T.; Howatson, G. Effects of Montmorency tart cherry (Prunus Cerasus L.) consumption on vascular function in men with early hypertension. Am. J. Clin. Nutr. 2016, 103, 1531–1539. [Google Scholar] [CrossRef] [Green Version]
- Matchynski, J.J.; Lowrance, S.A.; Pappas, C.; Rossignol, J.; Puckett, N.; Sandstrom, M.; Dunbar, G.L. Combinatorial treatment of tart cherry extract and essential fatty acids reduces cognitive impairments and inflammation in the mu–p75 saporin–induced mouse model of Alzheimer’s disease. J. Med. Food 2013, 16, 288–295. [Google Scholar] [CrossRef]
- Ducharme, N.G.; Fortier, L.A.; Kraus, M.S.; Hobo, S.; Mohammed, H.O.; McHugh, M.P.; Hackett, R.P.; Soderholm, L.V.; Mitchell, L.M. Effect of a tart cherry juice blend on exercise–induced muscle damage in horses. Am. J. Vet. Res. 2009, 70, 758–763. [Google Scholar] [CrossRef]
- Kim, J.W.; Ku, S.K.; Han, M.H.; Kim, K.Y.; Kim, S.G.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, C.M.; Choi, Y.H. The admin-istration of Fructus Schisandrae attenuates dexamethasone-induced muscle atrophy in mice. Int. J. Mol. Med. 2015, 36, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Jamall, I.S.; Smith, J.C. Effects of cadmium on glutathione peroxidase, superoxidase dismutase and lipid peroxidation in the rat heart: A possible mechanism of cadmium cardiotoxicity. Toxicol. Appl. Pharmacol. 1985, 80, 33–42. [Google Scholar] [CrossRef]
- He, H.-J.; Wang, G.-Y.; Gao, Y.; Ling, W.-H.; Guo-Yu, W.; Jin, T.-R. Curcumin attenuates Nrf2 signaling defect, oxidative stress in muscle and glucose intolerance in high fat diet-fed mice. World J. Diabetes 2012, 3, 94–104. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods in Enzymatic Analysis; Bergmeyer, H.U., Ed.; Academic Press: New York, NY, USA, 1974; pp. 673–686. [Google Scholar]
- Kandarian, S. The molecular basis of skeletal muscle atrophy—Parallels with osteoporotic signaling. J. Musculoskelet. Neuronal Interact. 2009, 8, 340–341. [Google Scholar]
- Powers, S.K.; Smuder, A.; Judge, A. Oxidative stress and disuse muscle atrophy: Cause or consequence? Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 240. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, T.; Katada, A.; Shigyo, H.; Kishibe, K.; Adachi, M.; Nonaka, S.; Harabuchi, Y. Electrical stimulation prevents apoptosis in denervated skeletal muscle. NeuroRehabilitation 2010, 27, 147–154. [Google Scholar] [CrossRef]
- Klopstock, T. Drug–induced myopathies. Curr. Opin. Neurol. 2008, 21, 590–595. [Google Scholar] [CrossRef]
- Le Quintrec, J.S.; Le Quintrec, J.L. Drug–induced myopathies. Baillières Clin. Rheumatol. 1991, 5, 21–38. [Google Scholar] [CrossRef]
- Strack, A.M.; Sebastian, R.J.; Schwartz, M.W.; Dallman, M.F. Glucocorticoids and insulin: Reciprocal signals for energy balance. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1995, 268, R142–R149. [Google Scholar] [CrossRef]
- Cooper, D.S.; Rivkees, S.A. Putting Propylthiouracil in Perspective. J. Clin. Endocrinol. Metab. 2009, 94, 1881–1882. [Google Scholar] [CrossRef] [Green Version]
- Hengge, U.R.; Stocks, K.; Wiehler, H.; Faulkner, S.; Esser, S.; Lorenz, C.; Jentzen, W.; Hengge, D.; Goos, M.; Dudley, R.E.; et al. Double-blind, randomized, placebo-controlled phase III trial of oxymetholone for the treatment of HIV wasting. AIDS 2003, 17, 699–710. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. Creatine: Endogenous Metabolite, Dietary, and Therapeutic Supplement. Annu. Rev. Nutr. 2007, 27, 241–261. [Google Scholar] [CrossRef] [Green Version]
- Cohen, I.; Bogin, E.; Chechick, A.; Rzetelny, V. Biochemical alterations secondary to disuse atrophy in the rat’s serum and limb tissues. Arch. Orthop. Trauma Surg. 1999, 119, 410–417. [Google Scholar] [CrossRef]
- Orzechowski, A.; Ostaszewski, P.; Wilczak, J.; Jank, M.; Bałasińska, B.; Wareski, P.; Fuller, J., Jr. Rats with a glucocorti-coid-induced catabolic state show symptoms of oxidative stress and spleen atrophy: The effects of age and recovery. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2002, 49, 256–263. [Google Scholar] [CrossRef]
- Pellegrino, M.A.; D’Antona, G.; Bortolotto, S.; Boschi, F.; Pastoris, O.; Bottinelli, R.; Polla, B.; Reggiani, C. Clenbuterol antago-nizes glucocorticoid-induced atrophy and fibre type transformation in mice. Exp. Physiol. 2004, 89, 89–100. [Google Scholar] [CrossRef]
- Bray, T.M.; Taylor, C.G. Tissue glutathione, nutrition, and oxidative stress. Can. J. Physiol. Pharmacol. 1993, 71, 746–751. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.L. Antioxidant signaling in skeletal muscle: A brief review. Exp. Gerontol. 2007, 42, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Zarkovic, N. 4-Hydroxynonenal as a bioactive marker of pathophysiological processes. Mol. Asp. Med. 2003, 24, 281–291. [Google Scholar] [CrossRef]
- Kanski, J.; Hong, S.J.; Schöneich, C. Proteomic analysis of protein nitration in aging skeletal muscle and identification of ni-trotyrosine–containing sequences in vivo by nanoelectrospray ionization tandem mass spectrometry. J. Biol. Chem. 2005, 280, 24261–24266. [Google Scholar] [CrossRef] [Green Version]
- Saborido, A.; Naudí, A.; Portero–Otín, M.; Pamplona, R.; Megías, A. Stanozolol treatment decreases the mitochondrial ROS generation and oxidative stress induced by acute exercise in rat skeletal muscle. J. Appl. Physiol. 2011, 110, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Li, J.S.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [Green Version]
- Dam, A.D.; Mitchell, A.S.; Rush, J.W.E.; Quadrilatero, J. Elevated skeletal muscle apoptotic signaling following glutathione depletion. Apoptosis 2011, 17, 48–60. [Google Scholar] [CrossRef]
- Glass, D.J. Signalling pathways that mediate skeletal muscle hypertrophy and atrophy. Nat. Cell Biol. 2003, 5, 87–90. [Google Scholar] [CrossRef]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo Transcription Factors Induce the Atrophy-Related Ubiquitin Ligase Atrogin-1 and Cause Skeletal Muscle Atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Egan, B.; Zierath, J.R. Exercise Metabolism and the Molecular Regulation of Skeletal Muscle Adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [Green Version]
- Sparks, H.V.; Gorman, M.W. Adenosine in the Local Regulation of Blood Flow: Current Controversies. In Topics and Perspectives in Adenosine Research; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 1987; pp. 406–415. [Google Scholar]
- Hellsten, Y.; Maclean, D.; Rådegran, G.; Saltin, B.; Bangsbo, J. Adenosine Concentrations in the Interstitium of Resting and Contracting Human Skeletal Muscle. Circulation 1998, 98, 6–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.G.; Hafen, P.S.; Brault, J.J. Increased Adenine Nucleotide Degradation in Skeletal Muscle Atrophy. Int. J. Mol. Sci. 2019, 21, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynge, J.; Hellsten, Y. Distribution of adenosine A1, A2A and A2B receptors in human skeletal muscle. Acta Physiol. Scand. 2000, 169, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Borea, P.A.; Varani, K.; Vincenzi, F.; Baraldi, P.G.; Tabrizi, M.A.; Merighi, S.; Gessi, S. The A3Adenosine Receptor: History and Perspectives. Pharmacol. Rev. 2015, 67, 74–102. [Google Scholar] [CrossRef] [Green Version]
- Faravelli, I.; Corti, S.P. Addressing Cell Therapy for Spinal Muscular Atrophy: Open Issues and Future Perspectives. In Spinal Muscular Atrophy; Academic Press: Cambridge, MA, USA, 2017; pp. 357–367. [Google Scholar]
- Elkina, Y.; Von Haehling, S.; Anker, S.D.; Springer, J. The role of myostatin in muscle wasting: An overview. J. Cachex-Sarcopenia Muscle 2011, 2, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Gautam, V.; Costantini, S.; Paladino, A.; Colonna, G. Interactomic and Pharmacological Insights on Human Sirt-1. Front. Pharmacol. 2012, 3, 40. [Google Scholar] [CrossRef] [Green Version]
- Haigis, M.C.; Guarente, L.P. Mammalian sirtuins—Emerging roles in physiology, aging, and calorie restriction. Genes Dev. 2006, 20, 2913–2921. [Google Scholar] [CrossRef] [Green Version]
- De Souza, G.L.; Hallak, J. Anabolic steroids and male infertility: A comprehensive review. BJU Int. 2011, 108, 1860–1865. [Google Scholar] [CrossRef]
- Akbari Bazm, M.; Goodarzi, N.; Shahrokhi, S.R.; Khazaei, M. The effects of hydroalcoholic extract of Vaccinium arctostaphylos L. on sperm parameters, oxidative injury and apoptotic changes in oxymetholone-induced testicular toxicity in mouse. Andrologia 2020, 52, e13522. [Google Scholar] [CrossRef]
- Akbari Bazm, M.; Khazaei, M.; Khazaei, F.; Naseri, L. Nasturtium Officinale L. hydroalcoholic extract improved oxymetholone-induced oxidative injury in mouse testis and sperm parameters. Andrologia 2019, 51, e13294. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | 5′–3′ | Sequence | Size (bp) | Gene ID |
|---|---|---|---|---|
| Atrogin-1 | Forward Reverse | CAGCTTCGTGAGCGACCT CGGCAGTCGAGAAGTCCAGTC | 244 | 67731 |
| MuRF 1 | Forward Reverse | GACAGTCGCATTTCAAAGCA GCCTAGCACTGACCTGGAAG | 194 | 433766 |
| PI3K p85α | Forward Reverse | GCCAGTGGTCATTTGTGTTG ACACAACCAGGGAAGTCCAG | 236 | 18708 |
| Akt 1 | Forward Reverse | ATGAACGACGTAGCCATTGTG TTGTAGCCAATAAAGGTGCCAT | 116 | 11651 |
| Adenosine A1R | Forward Reverse | TGTTCCCAGGGCCTTTCAC TAATGGACTGAGACTAGCTTGACTGGTA | 155 | 11539 |
| TRPV4 | Forward Reverse | CAGGACCTCTGGAAGAGTGC AAGAGCTAGCCTGGACACCA | 165 | 63873 |
| Myostatin | Forward Reverse | CCTCCACTCCGGGAACTGA AAGAGCCATCACTGCTGTCATC | 185 | 17700 |
| SIRT1 | Forward Reverse | TTCACATTGCATGTGTGTGG TGAGGCCCAGTGCTCTAACT | 175 | 93759 |
| 18s Ribosomal RNA | Forward Reverse | AGCCTGAGAAACGGCTACC TCCCAAGATCCAACTACGAG | 252 | 19791 |
| Antisera or Detection Kits | Code | Source | Dilution |
|---|---|---|---|
| Primary Antisera | |||
| Anti-cleaved caspase-3 (Asp175) polyclonal antibody | 9661 | Cell Signaling Technology Inc. 1 | 1:400 |
| Anti-cleaved PARP (Asp214) specific antibody | 9545 | Cell Signaling Technology Inc. 1 | 1:100 |
| Anti-4-Hydroxynonenal polyclonal antibody | Ab46545 | Abcam 2 | 1:100 |
| Anti-Nitrotyrosine polyclonal antibody | 06-284 | Millipore Corporation 3 | 1:200 |
| Anti-nitric oxide synthase2 (N-20) polyclonal antibody | sc-651 | Santa Cruz Biotechnology 4 | 1:100 |
| Anti-GDF8/Myoststin antibody | Ab71808 | Abcam 2 | 1:50 |
| Detection kits | |||
| Vectastain Elite ABC Kit | PK-6200 | Vector Lab. Inc. 4 | 1:50 |
| Peroxidae substrate kit | SK-4100 | Vector Lab. Inc. 4 | 1:50 |
| Items (Unit) Groups | Serum Levels | ||
|---|---|---|---|
| Creatinine (mg/dL) | Creatine Kinase (IU/L) | LDH (IU/L) | |
| Controls | |||
| Intact | 0.34 ± 0.04 | 83.75 ± 20.86 | 665.50 ± 128.94 |
| GLU | 0.86 ± 0.13 a | 281.63 ± 55.15 d | 168.25 ± 50.55 d |
| Reference | |||
| Oxymetholone | 0.44 ± 0.06 b,c | 148.00 ± 17.78 d,e | 318.50 ± 79.79 d,e |
| TCcp treated | |||
| 500 mg/kg | 0.55 ± 0.10 a,c | 186.00 ± 15.94 d,e | 281.50 ± 25.53 d,e |
| 250 mg/kg | 0.64 ± 0.09 a,c | 208.88 ± 20.95 d,f | 241.13 ± 38.72 d,e |
| 125 mg/kg | 0.83 ± 0.10 a | 270.25 ± 40.21 d | 178.88 ± 61.66 d |
| Items (Unit) Groups | Fundus Antioxidant Defense Systems | ||||
|---|---|---|---|---|---|
| Malondialdehyde (nM/mg Protein) | Reactive Oxygen Species (RFU/μg Protein) | Glutathione (nM/mg Protein) | Superoxide Dismutase (nM/mim/mg Protein) | Catalase (U/mg Protein) | |
| Controls | |||||
| Intact | 1.87 ± 0.75 | 22.57 ± 10.53 | 0.66 ± 0.16 | 35.39 ± 13.08 | 7.25 ± 2.02 |
| GLU | 8.54 ± 1.22 a | 70.01 ± 15.85 d | 0.17 ± 0.07 a | 11.97 ± 2.11 d | 2.07 ± 0.56 d |
| Reference | |||||
| Oxymetholone | 4.53 ± 0.93 a,b | 32.57 ± 11.84 f | 0.41 ± 0.12 a,b | 22.79 ± 4.32 e,f | 3.88 ± 0.63 d,f |
| TCcp treated | |||||
| 500 mg/kg | 5.19 ± 0.89 a,b | 39.40 ± 11.09 e,f | 0.31 ± 0.10 a,c | 19.36 ± 2.79 d,f | 3.56 ± 0.99 d,f |
| 250 mg/kg | 6.17 ± 1.25 a,b | 46.36 ± 13.36 d,g | 0.28 ± 0.06 a,c | 17.39 ± 2.19 d,f | 3.21 ± 0.92 d,f |
| 125 mg/kg | 8.22 ± 1.40 a | 66.31 ± 22.46 d | 0.18 ± 0.08 a | 12.77 ± 2.59 d | 2.19 ± 0.59 d |
| Groups Targets | Controls | Reference | TCcp Treated Mice (mg/kg) | |||
|---|---|---|---|---|---|---|
| Intact | GLU | Oxymetholone | 500 | 250 | 125 | |
| Atrogin-1 | 0.99 ± 0.07 | 4.89 ± 0.69 c | 2.26 ± 0.55 c,e | 3.00 ± 0.74 c,e | 3.59 ± 0.81 c,e | 4.59 ± 0.64 c |
| MuRF 1 | 1.08 ± 0.22 | 6.29 ± 1.10 c | 3.06 ± 0.67 c,e | 4.36 ± 1.07 c,f | 4.76 ± 0.67 c,e | 5.99 ± 1.88 c |
| PI3K p85α | 1.04 ± 0.14 | 0.59 ± 0.11 c | 1.15 ± 0.37 e | 0.88 ± 0.10 d,e | 0.77 ± 0.12 c,f | 0.64 ± 0.14 c |
| Akt 1 | 1.02 ± 0.05 | 0.54 ± 0.06 c | 0.87 ± 0.11 d,e | 0.74 ± 0.06 c,e | 0.67 ± 0.07 c,e | 0.57 ± 0.12 c |
| A1R | 1.05 ± 0.14 | 0.51 ± 0.13 a | 0.87 ± 0.07 a,b | 0.77 ± 0.08 a,b | 0.70 ± 0.10 a,b | 0.54 ± 0.17 a |
| TRPV4 | 1.11 ± 0.10 | 0.37 ± 0.09 a | 0.66 ± 0.10 a,b | 0.61 ± 0.13 a,b | 0.55 ± 0.10 a,b | 0.40 ± 0.10 a |
| Myostatin | 1.03 ± 0.10 | 7.02 ± 0.99 c | 3.23 ± 0.78 c,e | 4.72 ± 1.01 c,e | 5.12 ± 0.79 c,e | 6.51 ± 1.24 c |
| SIRT1 | 1.03 ± 0.17 | 10.73 ± 3.03 c | 3.73 ± 1.11 c,e | 4.57 ± 1.19 c,e | 6.27 ± 1.35 c,e | 10.02 ± 1.57 c |
| Groups Index | Controls | Reference | TCcp Treated Mice (mg/kg) | |||
|---|---|---|---|---|---|---|
| Intact | GLU | Oxymetholone | 500 | 250 | 125 | |
| General histomorphometry | ||||||
| Fiber diameter (μm) | 50.88 ± 11.00 | 23.10 ± 5.10 a | 38.38 ± 7.06 a,b | 34.85 ± 4.20 a,b | 31.67 ± 5.12 a,b | 24.94 ± 5.92 a |
| Collagen (%) | 4.23 ± 1.65 | 32.84 ± 4.89 a | 17.69 ± 5.06 a,b | 21.56 ± 2.96 a,b | 23.82 ± 3.51 a,b | 31.32 ± 4.81 a |
| Immunohistomorphometry (fibers/mm2) | ||||||
| Caspase-3 | 2.38 ± 2.50 | 42.13 ± 11.06 d | 21.00 ± 4.81 d,e | 27.38 ± 3.16 d,e | 29.75 ± 5.55 d,f | 39.50 ± 5.37 d |
| PARP | 5.13 ± 2.59 | 76.25 ± 10.95 a | 33.25 ± 10.35 a,b | 45.88 ± 10.45 a,b | 54.88 ± 10.49 a,b | 71.25 ± 12.06 a |
| Nitrotyrosine | 5.13 ± 2.64 | 68.38 ± 12.08 a | 32.13 ± 12.73 a,b | 46.88 ± 10.99 a,b | 51.50 ± 10.97 a,b | 66.00 ± 14.77 a |
| 4-HNE | 3.63 ± 2.07 | 76.50 ± 12.17 a | 44.13 ± 10.83 a,b | 48.75 ± 11.23 a,b | 56.75 ± 12.58 a,b | 71.50 ± 12.86 a |
| iNOS | 8.00 ± 3.16 | 52.88 ± 12.19 d | 21.75 ± 6.48 d,e | 34.00 ± 6.48 d,e | 38.25 ± 8.71 d,f | 51.00 ± 15.51 d |
| Myostatin | 1.38 ± 1.06 | 52.75 ± 10.63 d | 22.13 ± 4.76 d,e | 29.00 ± 8.73 d,e | 36.13 ± 11.24 d,e | 50.50 ± 11.38 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ku, S.-K.; Lim, J.-M.; Cho, H.-R.; Bashir, K.M.I.; Kim, Y.S.; Choi, J.-S. Tart Cherry (Fruit of Prunus cerasus) Concentrated Powder (TCcp) Ameliorates Glucocorticoid-Induced Muscular Atrophy in Mice. Medicina 2021, 57, 485. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57050485
Ku S-K, Lim J-M, Cho H-R, Bashir KMI, Kim YS, Choi J-S. Tart Cherry (Fruit of Prunus cerasus) Concentrated Powder (TCcp) Ameliorates Glucocorticoid-Induced Muscular Atrophy in Mice. Medicina. 2021; 57(5):485. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57050485
Chicago/Turabian StyleKu, Sae-Kwang, Jong-Min Lim, Hyung-Rae Cho, Khawaja Muhammad Imran Bashir, Young Suk Kim, and Jae-Suk Choi. 2021. "Tart Cherry (Fruit of Prunus cerasus) Concentrated Powder (TCcp) Ameliorates Glucocorticoid-Induced Muscular Atrophy in Mice" Medicina 57, no. 5: 485. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57050485