
Inhibitory Effect of Asplenium incisum on Bacterial Growth, Inflammation, and Osteoclastogenesis


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. A. incisum Extract
2.2. Antibacterial Assay and Sustainability of Antibacterial Activity
2.3. Anti-Inflammatory Activity by Measuring TNF-α, IL-6, and NO Levels
2.4. Anti-Osteoclastogenesis Assay Using TRAP Staining and TRAP Activity
2.5. Cytotoxicity Test
2.6. Statistical Analysis
3. Results
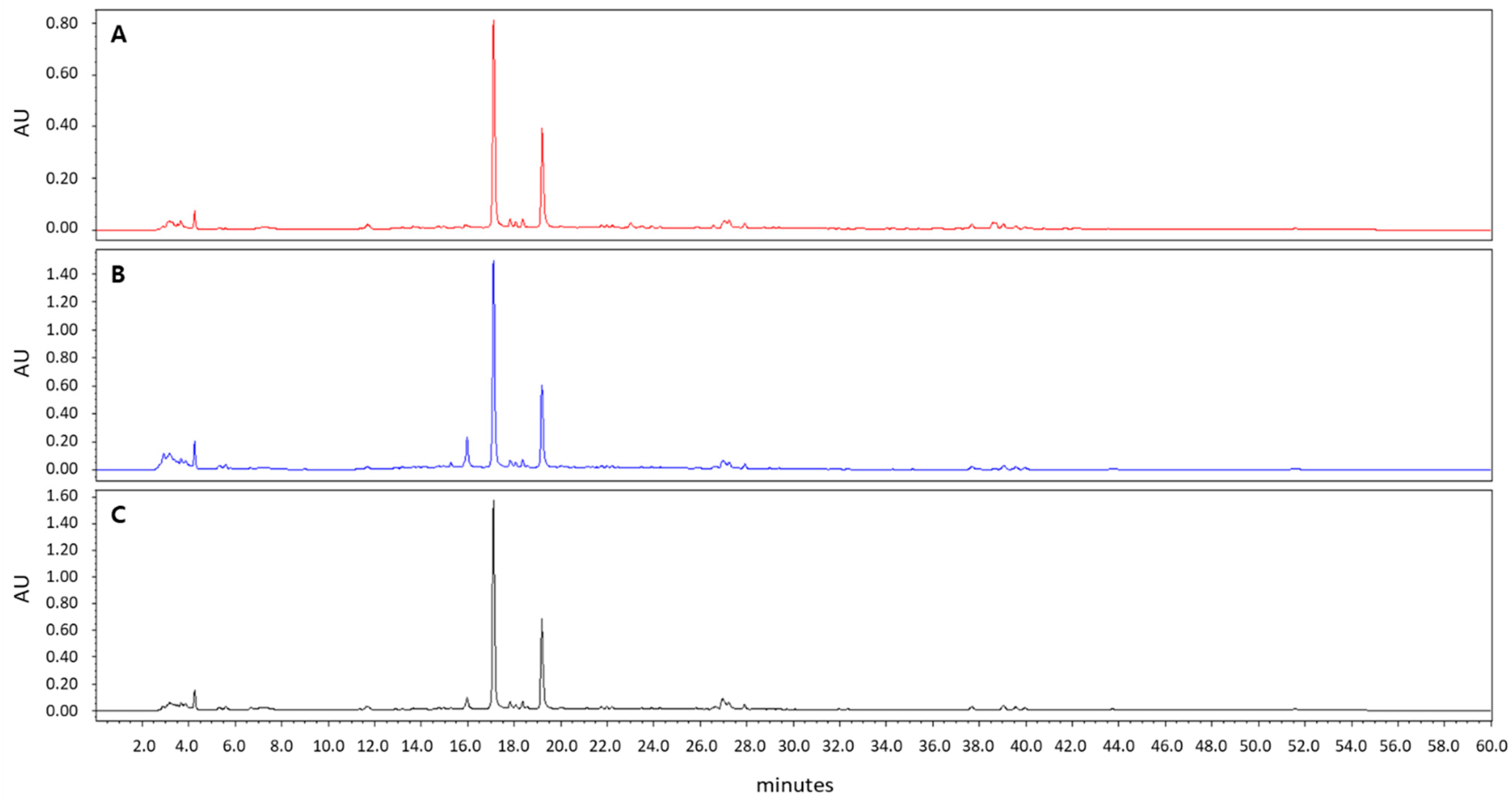
3.1. A. incisum Extract
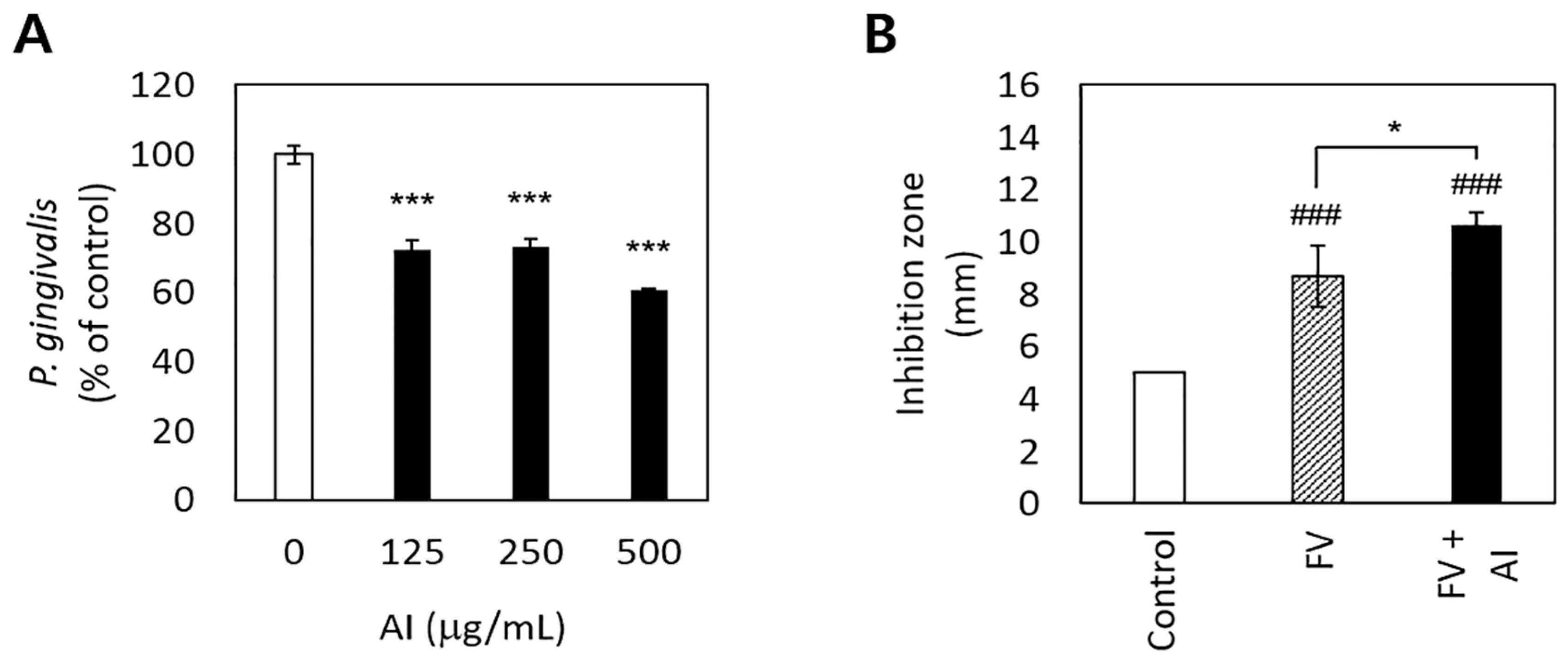
3.2. Antibacterial Effects and the Sustainability of Antibacterial Activity of A. Incisum on P. gingivalis
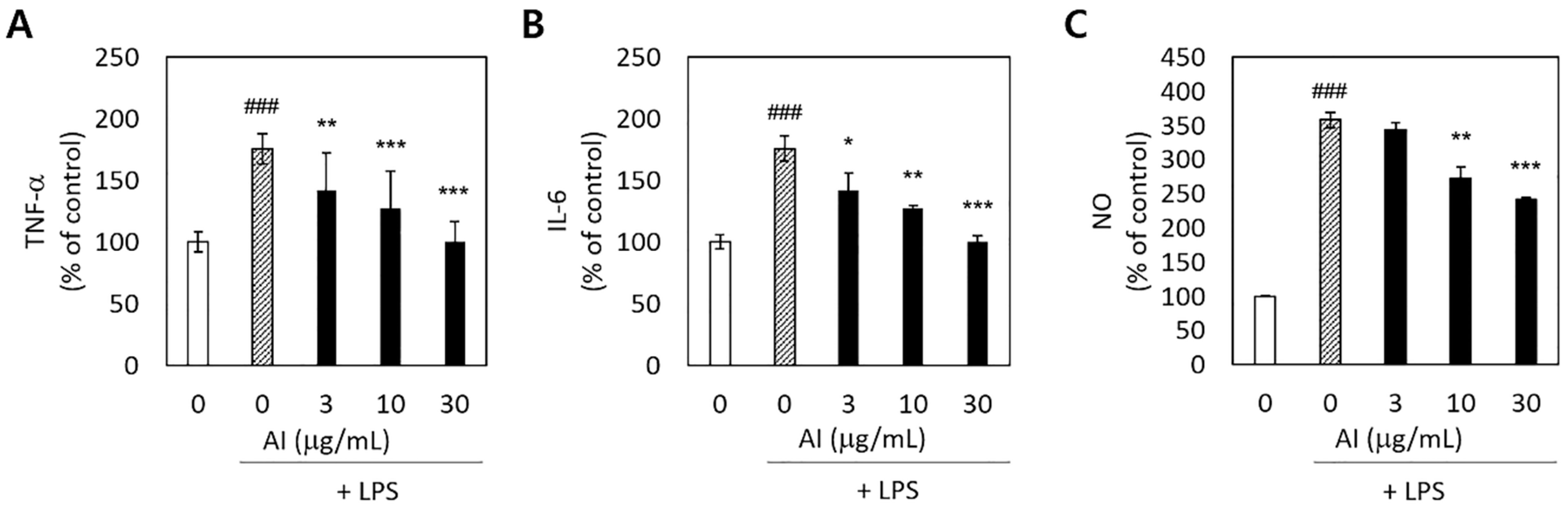
3.3. Anti-Inflammatory Activity of A. incisum
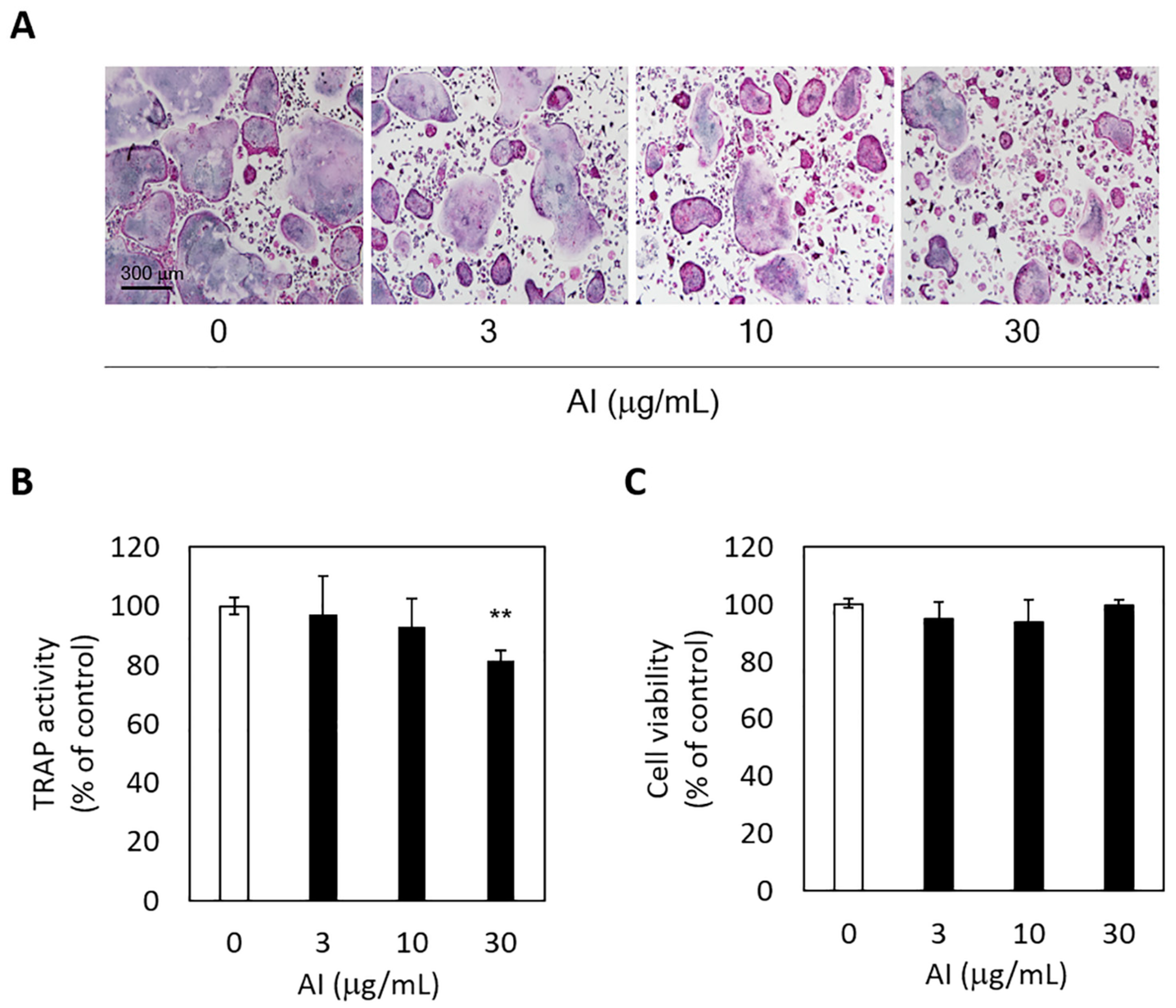
3.4. Anti-Osteoclastogenic Activity of A. incisum and Its Effect on Cell Viability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Listgarten, M.A. Pathogenesis of periodontitis. J. Clin. Periodontol. 1986, 13, 418–430. [Google Scholar] [CrossRef] [PubMed]
- How, K.Y.; Song, K.P.; Chan, K.G. Porphyromonas gingivalis: An overview of periodontopathic pathogen below the gum line. Front. Microbiol. 2016, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Persson, G.R. What has ageing to do with periodontal health and disease? Int. Dent. J. 2006, 56, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Hienz, S.A.; Paliwal, S.; Ivanovski, S. Mechanisms of bone resorption in periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitão, R.F.C.; Ribeiro, R.A.; Chaves, H.V.; Rocha, F.A.C.; Lima, V.; Brito, G.A.C. Nitric oxide synthase inhibition prevents alveolar bone resorption in experimental periodontitis in rats. J. Periodontol. 2005, 76, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Zhou, Y.; Chen, H.; Wang, C.; Wang, H. Antibacterial, anti-inflammatory, and anti-osteoclastogenesis roles of allicin in periodontitis. Int. J. Clin. Exp. Med. 2018, 11, 6721–6730. [Google Scholar]
- Soskolne, W.A.; Heasman, P.A.; Stabholz, A.; Smart, G.J.; Palmer, M.; Flashner, M.; Newman, H.N. Sustained local delivery of chlorhexidine in the treatment of periodontitis: A multi-center study. J. Periodontol. 1997, 68, 32–38. [Google Scholar] [CrossRef]
- Tabenski, L.; Moder, D.; Cieplik, F.; Schenke, F.; Hiller, K.A.; Buchalla, W.; Schmalz, G.; Christgau, M. Antimicrobial photodynamic therapy vs. local minocycline in addition to non-surgical therapy of deep periodontal pockets: A controlled randomized clinical trial. Clin. Oral Investig. 2017, 21, 2253–2264. [Google Scholar] [CrossRef]
- Greenstein, G. Clinical significance of bacterial resistance to tetracyclines in the treatment of periodontal diseases. J. Periodontol. 1995, 66, 925–932. [Google Scholar] [CrossRef]
- Teixeira, A.H.; Freire, J.M.O.; de Sousa, L.H.T.; Parente, A.T.; de Sousa, N.A.; Arriaga, A.M.C.; da Silva, F.R.L.; Melo, I.M.; da Silva, I.I.C.; Pereira, K.M.A. Stemodia maritima L. extract decreases inflammation, oxidative stress, and alveolar bone loss in an experimental periodontitis rat model. Front. Physiol. 2017, 8, 988. [Google Scholar] [CrossRef] [Green Version]
- Toker, H.; Ozan, F.; Ozer, H.; Ozdemir, H.; Eren, K.; Yeler, H. A morphometric and histopathologic evaluation of the effects of propolis on alveolar bone loss in experimental periodontitis in rats. J. Periodontol. 2008, 79, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Yoshinaga, Y.; Ukai, T.; Nakatsu, S.; Kuramoto, A.; Nagano, F.; Yoshinaga, M.; Montenegro, J.L.; Shiraishi, C.; Hara, Y. Green tea extract inhibits the onset of periodontal destruction in rat experimental periodontitis. J. Periodontal Res. 2014, 49, 652–659. [Google Scholar] [CrossRef]
- Inchingolo, F.; Tatullo, M.; Marrelli, M.; Inchingolo, A.M.; Picciariello, V.; Inchingolo, A.D.; Dipalma, G.; Vermesan, D.; Cagiano, R. Clinical trial with bromelain in third molar exodontia. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 771–774. [Google Scholar] [PubMed]
- Xu, Z.; Deng, M. Identification and Control of Common Weeds: Vol 2; Springer: Dordrecht, The Netherlands, 2017; Volume 2, pp. 66–67. [Google Scholar]
- Jeong, J.G. A herbalogical study on the plants of Aspleniaceae in Korea. Korea J. Herbol. 2011, 26, 59–65. [Google Scholar]
- Chen, C.; Peng, X.; Chen, J.; Wan, C. Antioxidant, antifungal activities of ethnobotanical Ficus hirta Vahl. and analysis of main constituents by HPLC-MS. Biomedicines 2020, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.H.; Ji, S.H.; Son, J.L.; Shin, S.J.; Oh, S.H.; Kim, S.; Bae, J.M. Antibacterial, anti-inflammatory, and anti-osteoclastogenic activities of Colocasia antiquorum var. esculenta: Potential applications in preventing and treating periodontal diseases. Dent. Mater. J. 2020, 39, 1096–1102. [Google Scholar]
- Lee, S.U.; Choi, Y.H.; Kim, Y.S.; Min, Y.K.; Rhee, M.; Kim, S.H. Anti-resorptive saurolactam exhibits in vitro anti-inflammatory activity via ERK-NF-kappaB signaling pathway. Int. Immunopharmacol. 2010, 10, 298–303. [Google Scholar] [CrossRef]
- Yeon, J.T.; Kim, K.J.; Son, Y.J.; Park, S.J.; Kim, S.H. Idelalisib inhibits osteoclast differentiation and pre-osteoclast migration by blocking the PI3Kδ-Akt-c-Fos/NFATc1 signaling cascade. Arch. Pharm. Res. 2019, 42, 712–721. [Google Scholar] [CrossRef]
- Moon, S.H.; Choi, S.W.; Kim, S.H. In vitro anti-osteoclastogenic activity of p38 inhibitor doramapimod via inhibiting migration of pre-osteoclasts and NFATc1 activity. J. Pharmacol. Sci. 2015, 129, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Cheng, N.; Tai, B.; Jiang, H.; Li, J.; Bian, Z. Randomized controlled trial on fluoride varnish application for treatment of white spot lesion after fixed orthodontic treatment. Clin. Oral Investig. 2012, 16, 463–468. [Google Scholar] [CrossRef]
- Iwashina, T.; Lopez-Saez, J.A.; Herrero, A.; Kitajima, J.; Matsumoto, S. Flavonol glycosides from Asplenium foreziense and its five related taxa and A. incisum. Biochem. Syst. Ecol. 2000, 28, 665–671. [Google Scholar] [CrossRef]
- Zeng, Y.; Nikikova, A.; Abdelsalam, H.; Li, J.; Xiao, J. Activity of quercetin and kaemferol against Streptococcus mutans biofilm. Arch. Oral Biol. 2019, 98, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Cvitkovitch, D.G.; Levesque, C.M. Cell death in Streptococcus mutans biofilms: A link between CSP and extracellular DNA. FEMS. Microbiol. Lett. 2009, 299, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Tagousop, C.N.; Tamokou, J.D.; Ekom, S.E.; Ngnokam, D.; Voutquenne-Nazabadioko, L. Antimicrobial activities of flavonoid glycosides from Graptophyllum grandulosum and their mechanism of antibacterial action. BMC Complement. Altern. Med. 2018, 18, 252. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Han, N.; Du, J.; Guo, L.; Luo, Z.; Liu, Y. Pathogenesis of important virulence factors of Porphyromonas gingivalis via Toll-like receptors. Front. Cell. Infect. Microbiol. 2019, 9, 262. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.B.; Kim, E.K.; Park, S.J.; Bang, S.G.; Kim, T.G.; Chung, D.W. Isolation and characterization of nicotiflorin obtained by enzymatic hydrolysis of two precursors in tea seed extract. J. Agric. Food Chem. 2010, 58, 4808–4813. [Google Scholar] [CrossRef] [PubMed]
- Nhiem, N.X.; Tai, B.H.; Quang, T.H.; Kiem, P.V.; Minh, C.V.; Nam, N.H.; Kim, J.H.; Im, L.R.; Lee, Y.M.; Kim, Y.H. A new ursane-type triterpenoid glycoside from Centella asiatica leaves modulates the production of nitric oxide and secretion of TNF-alpha in activated RAW 264.7 cells. Bioorg. Med. Chem. Lett. 2011, 21, 1777–1781. [Google Scholar] [CrossRef]
- Uğar-Cankal, D.; Ozmeric, N. A multifaceted molecule, nitric oxide in oral and periodontal diseases. Clin. Chim. Acta. 2006, 366, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Mira-Pascual, L.; Patlaka, C.; Desai, S.; Paulie, S.; Näreoja, T.; Lång, P.; Andersson, G. A novel sandwich ELISA for tartrate-resistant acid phosphatase 5a and 5b protein reveals that both isoforms are secreted by differentiating osteoclasts and correlate to the type I collagen degradation marker CTX-I in vivo and in vitro. Calcif. Tissue Int. 2020, 106, 194–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt, A.; Koperski, K.; Schumacher, M.; Gelinsky, M. Relevance of osteoclast-specific enzyme activities in cell-based in vitro resorption assays. Eur. Cell. Mater. 2017, 33, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.S.; Eyre, D.R.; Gundberg, C.M. Molecular basis and clinical application of biological markers of bone turnover. Endocr. Rev. 1996, 17, 333–368. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, S.-H.; Son, J.-L.; Shin, S.-J.; Oh, S.-H.; Kim, S.-H.; Bae, J.-M. Inhibitory Effect of Asplenium incisum on Bacterial Growth, Inflammation, and Osteoclastogenesis. Medicina 2021, 57, 641. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57070641
Moon S-H, Son J-L, Shin S-J, Oh S-H, Kim S-H, Bae J-M. Inhibitory Effect of Asplenium incisum on Bacterial Growth, Inflammation, and Osteoclastogenesis. Medicina. 2021; 57(7):641. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57070641
Chicago/Turabian StyleMoon, Seong-Hee, Ju-Lee Son, Seong-Jin Shin, Seung-Han Oh, Seong-Hwan Kim, and Ji-Myung Bae. 2021. "Inhibitory Effect of Asplenium incisum on Bacterial Growth, Inflammation, and Osteoclastogenesis" Medicina 57, no. 7: 641. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina57070641