
Anti-Mycoplasma Activity of Bacilotetrins C–E, Cyclic Lipodepsipeptides from the Marine-Derived Bacillus subtilis and Structure Revision of Bacilotetrins A and B


Abstract
:1. Introduction
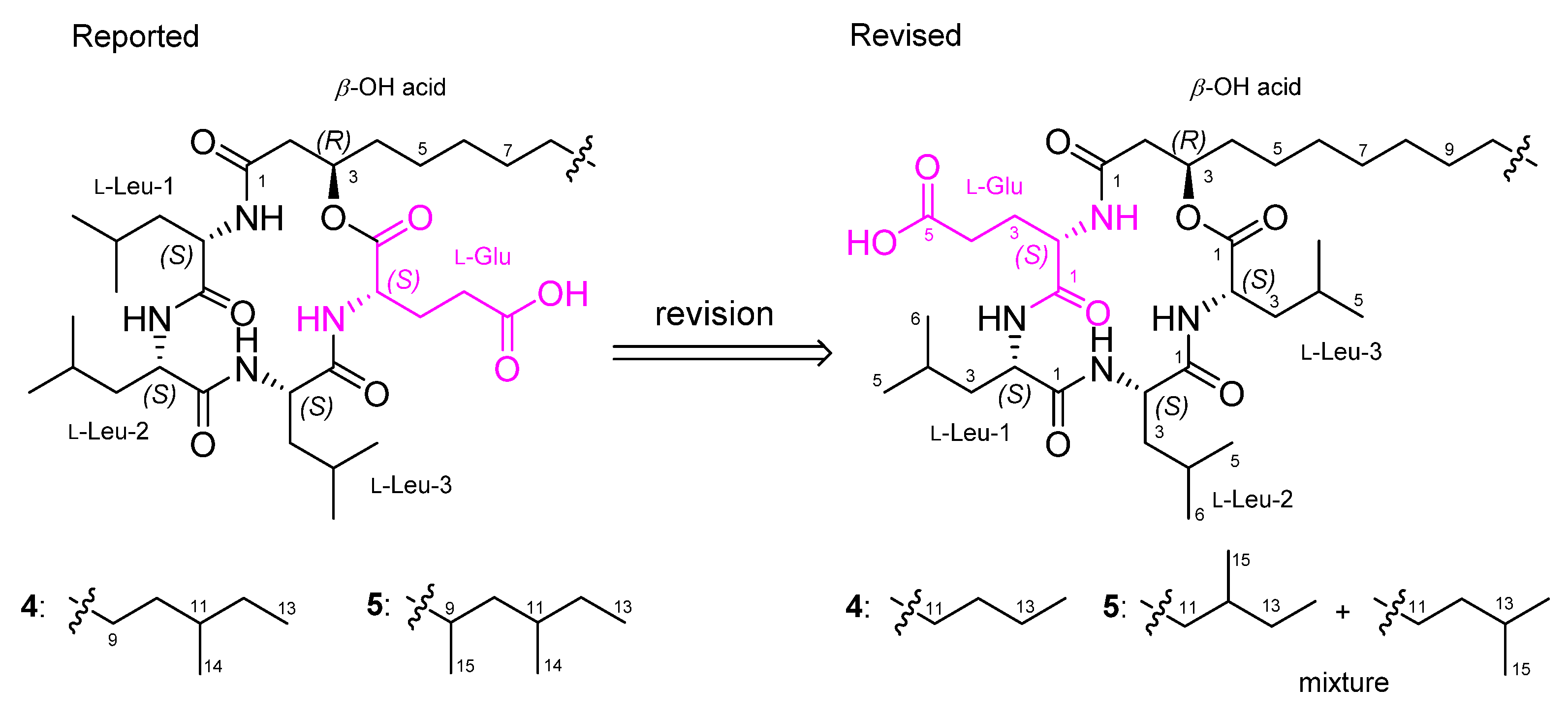
2. Results and Discussion
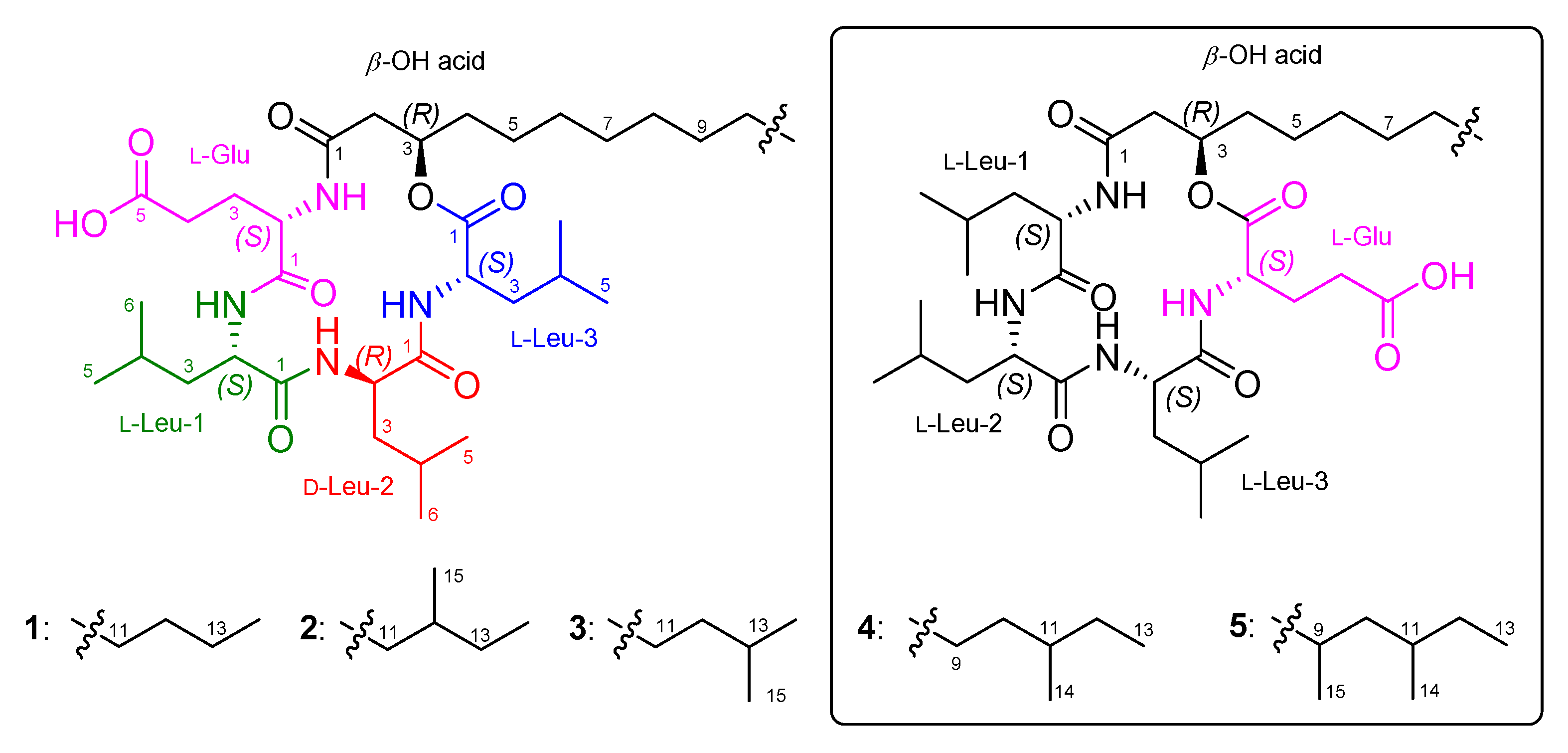
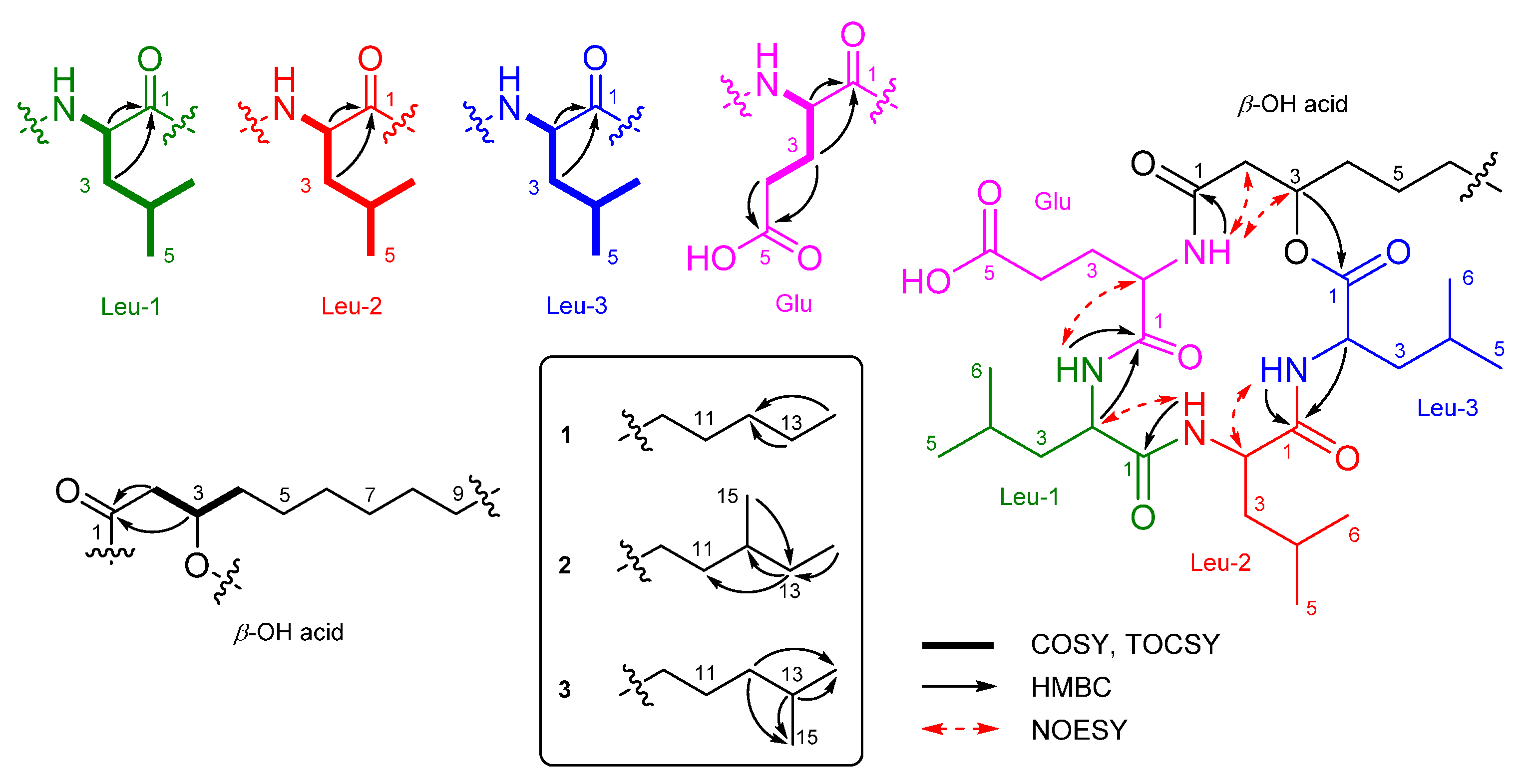
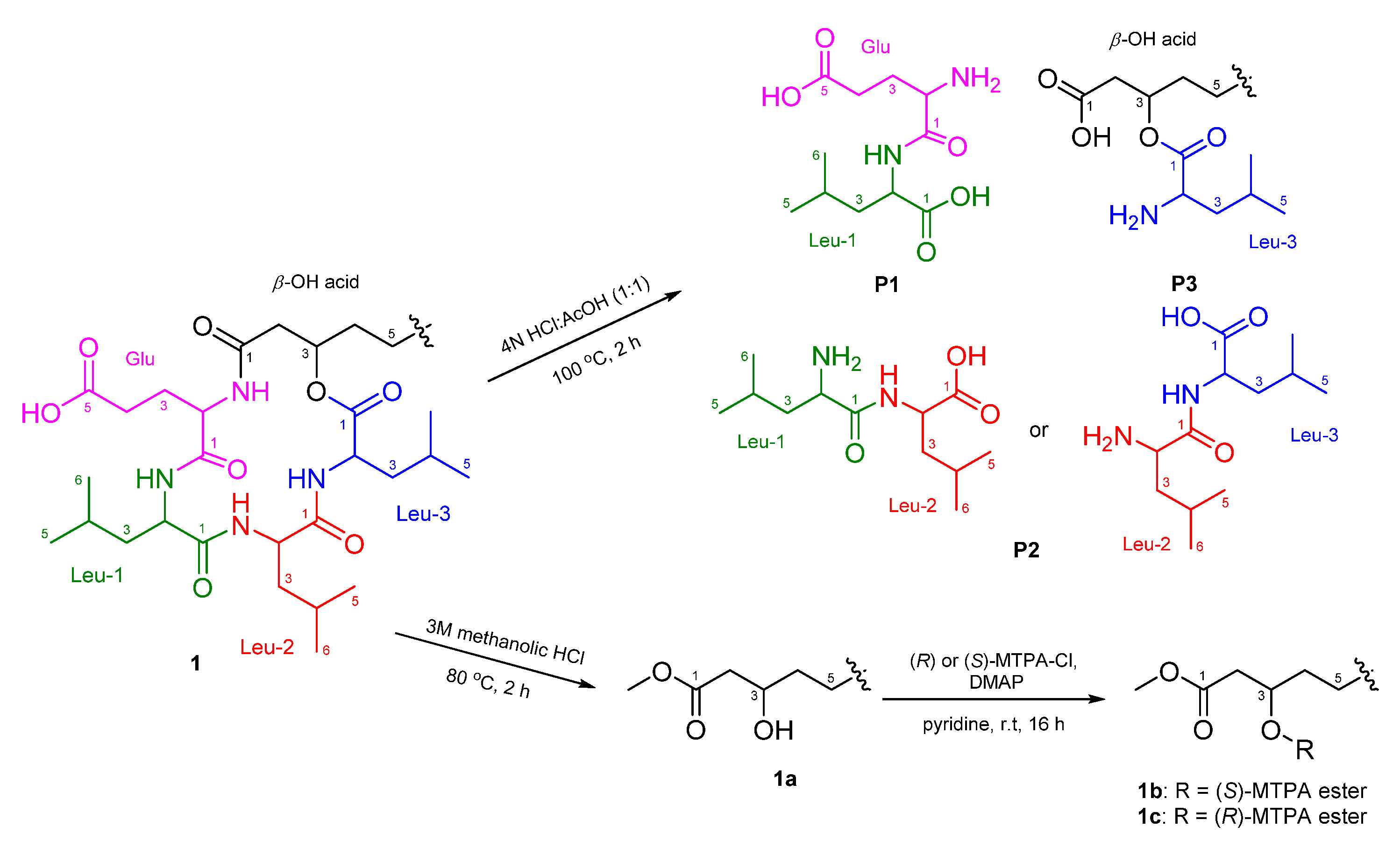
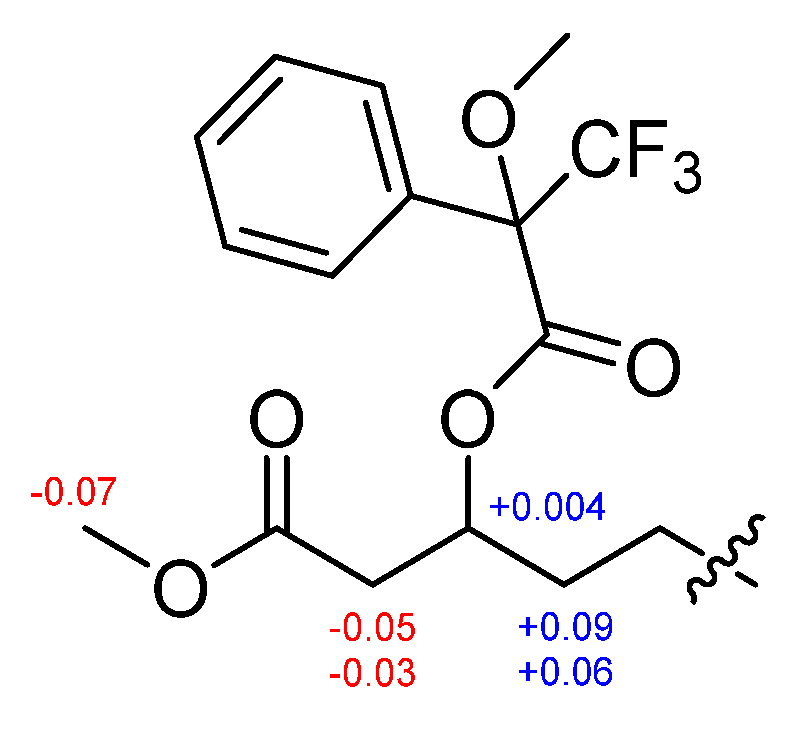
2.1. Identification of Isolated Compounds
2.2. Inhibitory Activity of Isolated Compounds against Mycoplasma hyorhinis
3. Materials and Methods
3.1. General Experimental Procedures and Reagents
3.2. Micro-Organism and Fermentation
3.3. Extraction and Isolation of Compounds 1–3
3.4. Total Hydrolysis and Marfey’s Analysis
3.5. Partial Hydrolysis and Marfey’s Analysis
3.6. Methanolysis of 1
3.7. Preparation of the (S)- and (R)-MTPA Esters (1b and 1c)
3.8. Measurement of Anti-Mycoplasma Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Mondol, M.A.; Shin, H.J.; Islam, M.T. Diversity of secondary metabolites from marine Bacillus species: Chemistry and biological activity. Mar. Drugs 2013, 11, 2846–2872. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, F.; Neubauer, P.; Gimpel, M. Bioactive Secondary Metabolites from Bacillus subtilis: A Comprehensive Review. J. Nat. Prod. 2019, 82, 2038–2053. [Google Scholar] [CrossRef] [PubMed]
- Tareq, F.S.; Kim, J.H.; Lee, M.A.; Lee, H.S.; Lee, J.S.; Lee, Y.J.; Shin, H.J. Antimicrobial gageomacrolactins characterized from the fermentation of the marine-derived bacterium Bacillus subtilis under optimum growth conditions. J. Agric. Food Chem. 2013, 61, 3428–3434. [Google Scholar] [CrossRef]
- Tareq, F.S.; Lee, M.A.; Lee, H.S.; Lee, Y.J.; Lee, J.S.; Hasan, C.M.; Islam, M.T.; Shin, H.J. Gageotetrins A-C, noncytotoxic antimicrobial linear lipopeptides from a marine bacterium Bacillus subtilis. Org. Lett. 2014, 16, 928–931. [Google Scholar] [CrossRef]
- Tareq, F.S.; Lee, M.A.; Lee, H.S.; Lee, Y.J.; Lee, J.S.; Hasan, C.M.; Islam, M.T.; Shin, H.J. Non-cytotoxic antifungal agents: Isolation and structures of gageopeptides A-D from a Bacillus strain 109GGC020. J. Agric. Food Chem. 2014, 62, 5565–5572. [Google Scholar] [CrossRef] [PubMed]
- Tareq, F.S.; Lee, M.A.; Lee, H.S.; Lee, J.S.; Lee, Y.J.; Shin, H.J. Gageostatins A-C, antimicrobial linear lipopeptides from a marine Bacillus subtilis. Mar. Drugs 2014, 62, 871–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tareq, F.S.; Hasan, C.M.; Lee, H.S.; Lee, Y.J.; Lee, J.S.; Surovy, M.Z.; Islam, M.T.; Shin, H.J. Gageopeptins A and B, new inhibitors of zoospore motility of the phytopathogen Phytophthora capsici from a marine-derived bacterium Bacillus sp. 109GGC020. Bioorg. Med. Chem. Lett. 2015, 25, 3325–3329. [Google Scholar] [CrossRef] [PubMed]
- Tareq, F.S.; Shin, H.J. Bacilotetrins A and B, Anti-Staphylococcal Cyclic-Lipotetrapeptides from a Marine-Derived Bacillus subtilis. J. Nat. Prod. 2017, 80, 2889–2892. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, M.; Mahmud, N.U.; Gupta, D.R.; Tareq, F.S.; Shin, H.J.; Islam, T. Inhibitory Effects of Linear Lipopeptides from a Marine Bacillus subtilis on the Wheat Blast Fungus Magnaporthe oryzae Triticum. Front. Microbiol. 2020, 11, 665. [Google Scholar] [CrossRef]
- Benedetti, F.; Krishnan, S.; Cocchi, F.; Tettelin, H.; Gallo, R.C.; Zella, D.; Curreli, S. Proteome analysis of Mycoplasma fermentans cultured under aerobic and anaerobic conditions. Transl. Med. Commun. 2019, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Pottinger, P.; Reller, L.B.; Ryan, K.J.; Weissman, S. Mycoplasma. In Sherris Medical Microbiology, 7th ed.; Ryan, K.J., Ed.; McGraw-Hill: New York, NY, USA, 2017; pp. 701–705. [Google Scholar]
- Degeling, M.H.; Maguire, C.A.; Bovenberg, M.S.S.; Tannous, B.A. Sensitive Assay for Mycoplasma Detection in Mammalian Cell Culture. Anal. Chem. 2012, 84, 4227–4232. [Google Scholar] [CrossRef] [Green Version]
- Dorottya, F.; Zsuzsa, K.; Miklos, G. Mycoplasma hyorhinis infection in swine. Magy. Allatorv. Lapja 2020, 142, 515–524. [Google Scholar]
- Friis, N.F.; Feenstra, A.A. Mycoplasma Hyorhinis in the Etiology of Serositis among Piglets. Acta. Vet. Scand. 1994, 35, 93–98. [Google Scholar] [PubMed]
- Siqueira, F.M.; Thompson, C.E.; Virginio, V.G.; Gonchoroski, T.; Reolon, L.; Almeida, L.G.; da Fonsêca, M.M.; de Souza, R.; Prosdocimi, F.; Schrank, I.S.; et al. New insights on the biology of swine respiratory tract mycoplasmas from a comparative genome analysis. BMC Genom. 2013, 14, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resende, T.P.; Pieters, M.; Vannucci, F.A. Swine conjunctivitis outbreaks associated with Mycoplasma hyorhinis. J. Vet. Diagn. Invest. 2019, 31, 766–769. [Google Scholar] [CrossRef]
- Morita, T.; Ohiwa, S.; Shimada, A.; Kazama, S.; Yagihashi, T.; Umemura, T. Intranasally inoculated Mycoplasma hyorhinis causes eustachitis in pigs. Vet. Pathol. 1999, 36, 174–178. [Google Scholar] [CrossRef]
- Robertson, A.W.; Sandoval, J.; Mohamed, O.G.; Zhuang, Y.; Gallagher, E.E.; Schmidt, J.; Caratelli, L.; Menon, A.; Schultz, P.J.; Torrez, R.M.; et al. Discovery of Surfactins as Inhibitors of MicroRNA Processing Using Cat-ELCCA. ACS Med. Chem. Lett. 2021, 12, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Ikai, Y.; Oka, H.; Suzuki, M.; Harada, K. A nonempirical method using LC/MS for determination of the absolute configuration of constituent amino acids in a peptide: Combination of Marfey’s method with mass spectrometry and its practical application. Anal. Chem. 1997, 69, 5146–5151. [Google Scholar] [CrossRef]
- Harada, K.-I.; Fujii, K.; Hayashi, K.; Suzuki, M.; Ikai, Y.; Oka, H. Application of D,L-FDLA derivatization to determination of absolute configuration of constituent amino acids in peptide by advanced Marfey’s method. Tetrahedron Lett. 1996, 37, 3001–3004. [Google Scholar] [CrossRef]
- Bonmatin, J.M.; Laprevote, O.; Peypoux, F. Diversity among microbial cyclic lipopeptides: Iturins and surfactins. Activity-structure relationships to design new bioactive agents. Comb. Chem. High Throughput Screen. 2003, 6, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Bruner, S.D.; Weber, T.; Kohli, R.M.; Schwarzer, D.; Marahiel, M.A.; Walsh, C.T.; Stubbs, M.T. Structural basis for the cyclization of the lipopeptide antibiotic surfactin by the thioesterase domain SrfTE. Structure 2002, 10, 301–310. [Google Scholar] [CrossRef]
- Koglin, A.; Lohr, F.; Bernhard, F.; Rogov, V.V.; Frueh, D.P.; Strieter, E.R.; Mofid, M.R.; Guntert, P.; Wagner, G.; Walsh, C.T.; et al. Structural basis for the selectivity of the external thioesterase of the surfactin synthetase. Nature 2008, 454, 907–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morozumi, M.; Hasegawa, K.; Kobayashi, R.; Inoue, N.; Iwata, S.; Kuroki, H.; Kawamura, N.; Nakayama, E.; Tajima, T.; Shimizu, K.; et al. Emergence of macrolide-resistant Mycoplasma pneumoniae with a 23S rRNA gene mutation. Antimicrob. Agents Chemother. 2005, 49, 2302–2306. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 | 2 | 3 | |||
|---|---|---|---|---|---|---|
| δC, Type | δH, m (J in Hz) | δC, Type | δH, m (J in Hz) | δC, Type | δH, m (J in Hz) | |
| Glu | ||||||
| 1 | 175.9, C | 175.9, C | 175.9, C | |||
| 2 | 55.6, CH | 4.11, td (7.3, 4.0) | 55.5, CH | 4.11, m | 55.5, CH | 4.11, m |
| 3 | 27.3, CH2 | 1.95, q (7.5) | 27.3, CH2 | 1.95, q (7.5) | 27.3, CH2 | 1.94, q (7.4) |
| 4 | 31.1, CH2 | 2.42, m | 31.1, CH2 | 2.43, m | 31.1, CH2 | 2.42, m |
| 5 | 176.3, C | 176.3, C | 176.3, C | |||
| NH | 8.39, d (4.7) | 8.42, d (4.5) | 8.41, d (4.5) | |||
| Leu-1 | ||||||
| 1 | 173.9, C | 173.9, C | 173.9, C | |||
| 2 | 55.0, CH | 3.74, m | 55.0, CH | 3.75, m | 55.0, CH | 3.74, m |
| 3 | 38.5, CH2 | 2.01, ddd (14.6, 11.0, 3.9) 1.80, o.l a | 38.5, CH2 | 2.01, m 1.80, o.l a | 38.5, CH2 | 2.01, m 1.80, o.l a |
| 4 | 26.3, CH | 1.61, m | 26.3, CH | 1.60, o.l a | 26.3, CH | 1.60, o.l a |
| 5 | 21.3, CH3 | 0.93, d (5.8) | 21.3, CH3 | 0.93, d (5.4) | 21.3, CH3 | 0.94, d (5.3) |
| 6 | 24.0, CH3 | 0.95, d (5.8) | 24.0, CH3 | 0.95, d (5.4) | 24.0, CH3 | 0.95, d (5.3) |
| NH | 9.10, d (6.7) | 9.13, d (6.6) | 9.12, d (6.7) | |||
| Leu-2 | ||||||
| 1 | 174.5, C | 174.5, C | 174.5, C | |||
| 2 | 53.4, CH | 4.43, m | 53.3, CH | 4.42, m | 53.4, CH | 4.42, m |
| 3 | 40.4, CH2 | 1.81, o.l a | 40.4, CH2 | 1.76, o.l a | 40.4, CH2 | 1.78, o.l a |
| 4 | 26.2, CH | 1.70, m | 26.2, CH | 1.71, o.l a | 26.2, CH | 1.71, o.l a |
| 5 | 21.2, CH3 | 0.90, d (6.4) | 21.2, CH3 | 0.90, d (6.4) | 21.2, CH3 | 0.91, d (6.4) |
| 6 | 23.9, CH3 | 0.96, d (6.4) | 23.9, CH3 | 0.96, d (6.4) | 23.9, CH3 | 0.98, d (6.4) |
| NH | 7.74, d (8.6) | 7.75, d (8.5) | 7.74, d (8.8) | |||
| Leu-3 | ||||||
| 1 | 172.9, C | 173.0, C | 173.0, C | |||
| 2 | 51.6, CH | 4.57, m | 51.6, CH | 4.57, m | 51.6, CH | 4.57, m |
| 3 | 40.6, CH2 | 1.80, m 1.69, m | 40.6, CH2 | 1.81, m 1.69, m | 40.6, CH2 | 1.80, m 1.70, m |
| 4 | 25.8, CH | 1.65, o.l a | 25.7, CH | 1.65, o.l a | 25.7, CH | 1.66, m |
| 5 | 21.8, CH3 | 0.89, o.l a | 21.7, CH3 | 0.89, d (6.4) | 21.7, CH3 | 0.90, d (6.5) |
| 6 | 23.8, CH3 | 0.92, d (6.5) | 23.7, CH3 | 0.92, d (6.4) | 23.7, CH3 | 0.93, d (6.5) |
| NH | 7.76, d (9.5) | 7.77, d (9.5) | 7.77, d (9.4) | |||
| β-OH acid | ||||||
| 1 | 173.3, C | 173.3, C | 173.0, C | |||
| 2 | 41.5, CH2 | 2.72, dd (13.8, 4.6) 2.29, dd (13.8, 8.1) | 41.5, CH2 | 2.72, dd (13.8, 4.6) 2.29, dd (13.8, 8.1) | 41.5, CH2 | 2.72, dd (13.8, 4.7) 2.29, dd (13.8, 8.1) |
| 3 | 73.8, CH | 5.16, tt (7.8, 5.3) | 73.8, CH | 5.15, m | 73.8, CH | 5.15, m |
| 4 | 35.5, CH2 | 1.80, o.l a 1.57, o.l a | 35.4, CH2 | 1.81, o.l a 1.56, o.l a | 35.4, CH2 | 1.81, o.l a 1.62, o.l a |
| 5 | 26.4– 30.8, CH2 | 1.29, o.l a | 28.2– 30.6, CH2 | 1.29, o.l a | 26.3, CH2 | 1.28, o.l a |
| 6 | 28.6– 31.0, CH2 | 1.28, o.l a | ||||
| 7 | ||||||
| 8 | ||||||
| 9 | ||||||
| 10 | ||||||
| 11 | 30.7, CH2 | 1.34, o.l a 1.13, o.l a | ||||
| 12 | 33.1, CH2 | 1.27, o.l a | 35.7, CH | 1.29, o.l a | 40.3, CH2 | 1.16, m |
| 13 | 23.8, CH2 | 1.30, o.l a | 37.8, CH2 | 1.29, o.l a 1.09, o.l a | 29.2, CH | 1.51, m |
| 14 | 14.5, CH3 | 0.89, o.l a | 11.8, CH3 | 0.87, o.l a | 23.1, CH3 | 0.87, d (6.4) |
| 15 | 19.7, CH3 | 0.85, d (4.8) | 23.1, CH3 | 0.87, d (6.4) | ||
| Compounds | ||||
|---|---|---|---|---|
| 1 | 2 | 3 | BioMycoX® 1 | |
| MIC (μg/mL) | 31 | 31 | 31 | 62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-S.; Shin, H.J. Anti-Mycoplasma Activity of Bacilotetrins C–E, Cyclic Lipodepsipeptides from the Marine-Derived Bacillus subtilis and Structure Revision of Bacilotetrins A and B. Mar. Drugs 2021, 19, 528. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100528
Lee H-S, Shin HJ. Anti-Mycoplasma Activity of Bacilotetrins C–E, Cyclic Lipodepsipeptides from the Marine-Derived Bacillus subtilis and Structure Revision of Bacilotetrins A and B. Marine Drugs. 2021; 19(10):528. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100528
Chicago/Turabian StyleLee, Hwa-Sun, and Hee Jae Shin. 2021. "Anti-Mycoplasma Activity of Bacilotetrins C–E, Cyclic Lipodepsipeptides from the Marine-Derived Bacillus subtilis and Structure Revision of Bacilotetrins A and B" Marine Drugs 19, no. 10: 528. https://0-doi-org.brum.beds.ac.uk/10.3390/md19100528