
Subcutaneous ω-Conotoxins Alleviate Mechanical Pain in Rodent Models of Acute Peripheral Neuropathy


 , and
, and 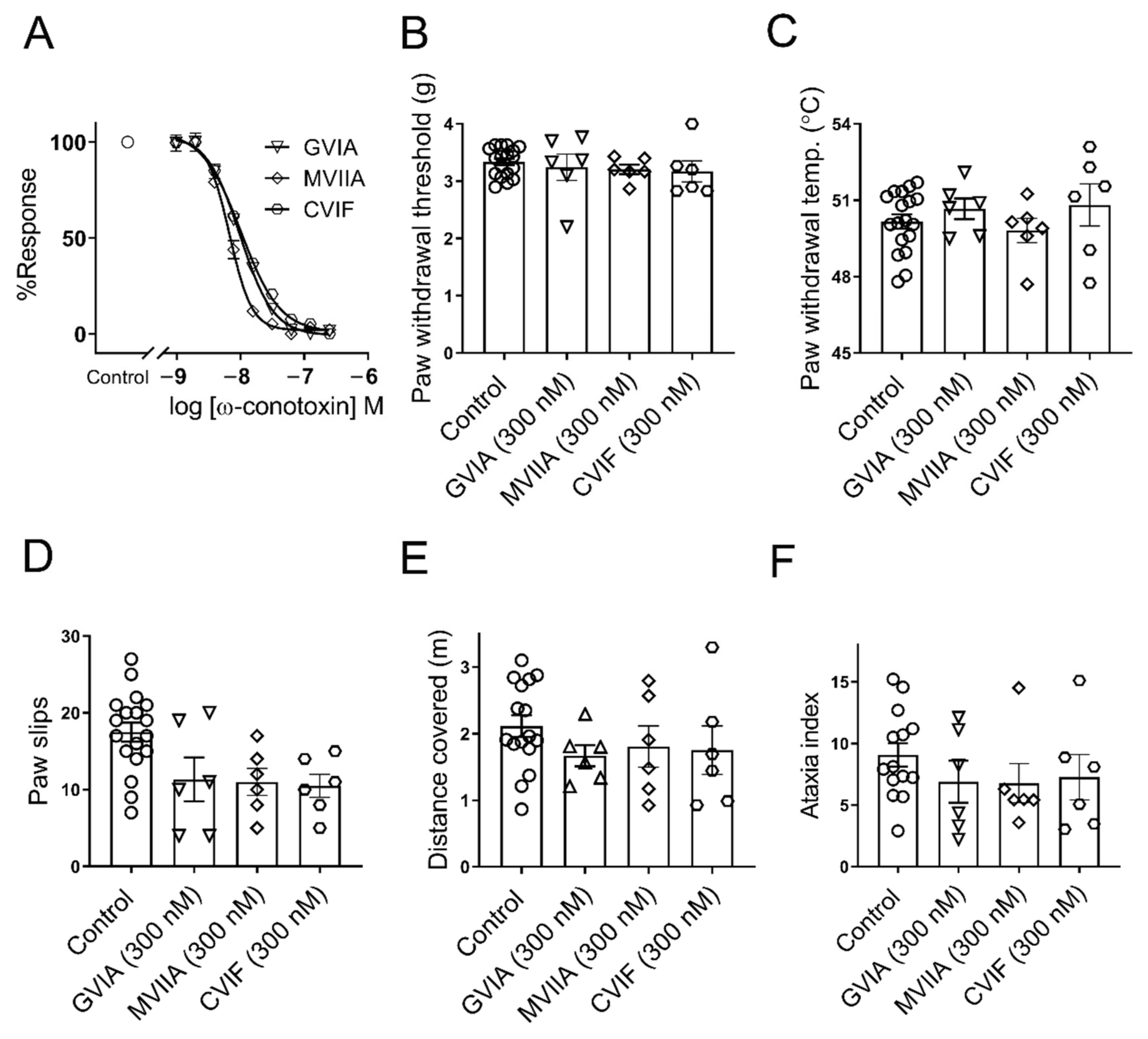{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. ω-Conotoxins Inhibit Endogenous CaV2.2 Channel In Vitro and Lack Side Effects In Vivo
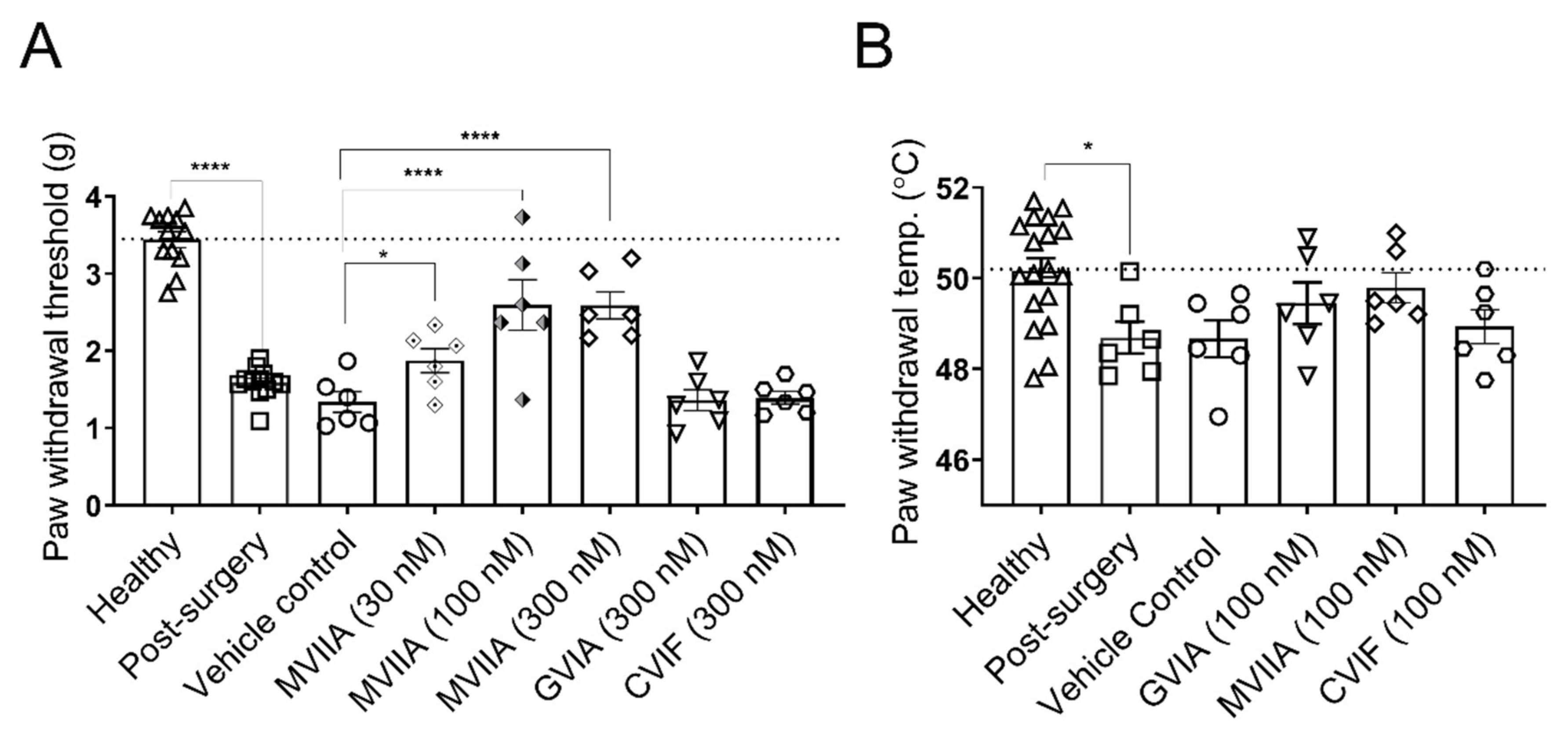
2.2. Local Effects of ω-Conotoxins on Surgery-Induced Mechanical and Thermal Allodynia
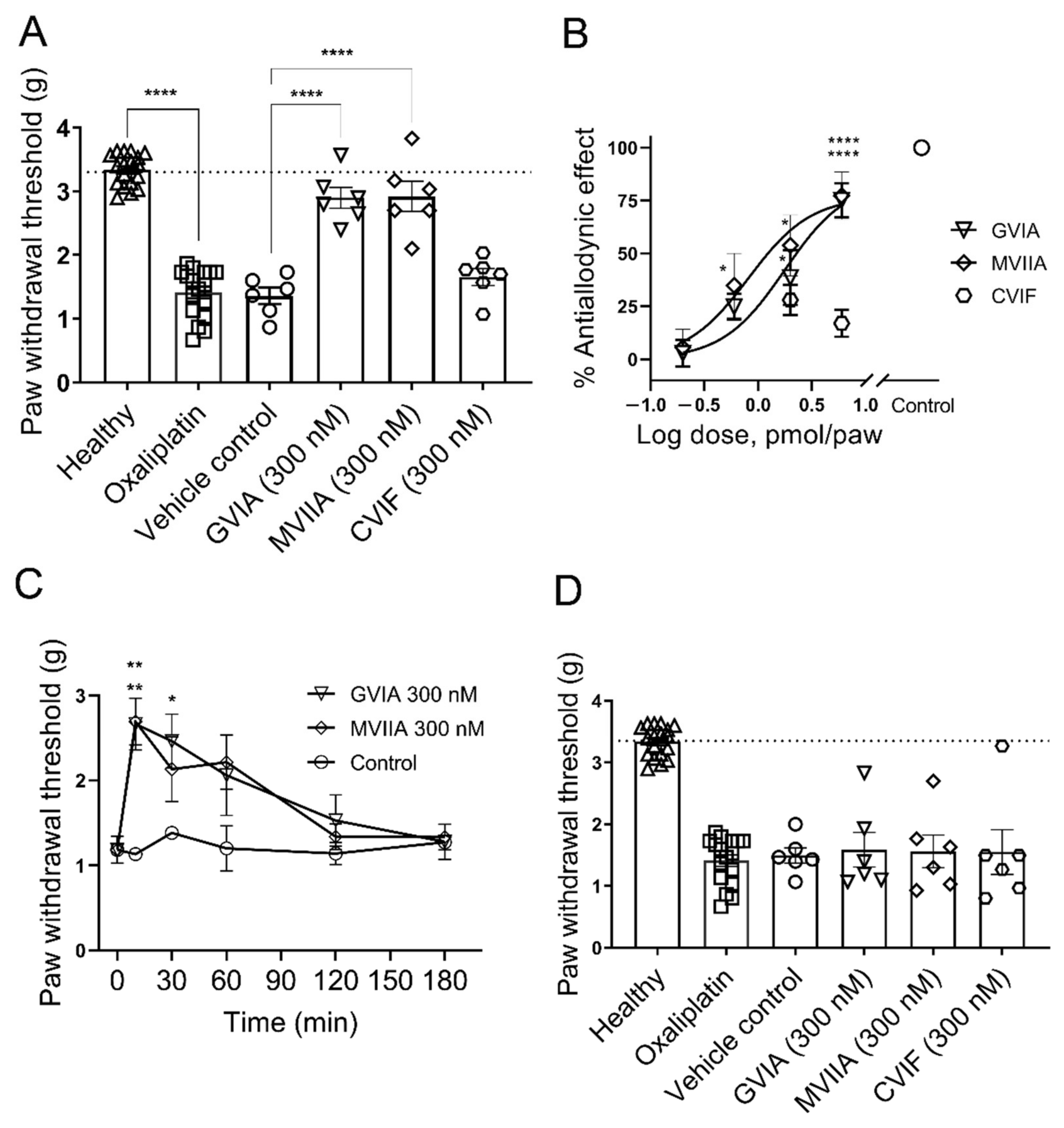
2.3. Local Effects of ω-Conotoxins on Chemotherapy-Induced Mechanical Allodynia
2.3.1. Effect of ω-Conotoxins in Oxaliplatin-Induced Mechanical Allodynia
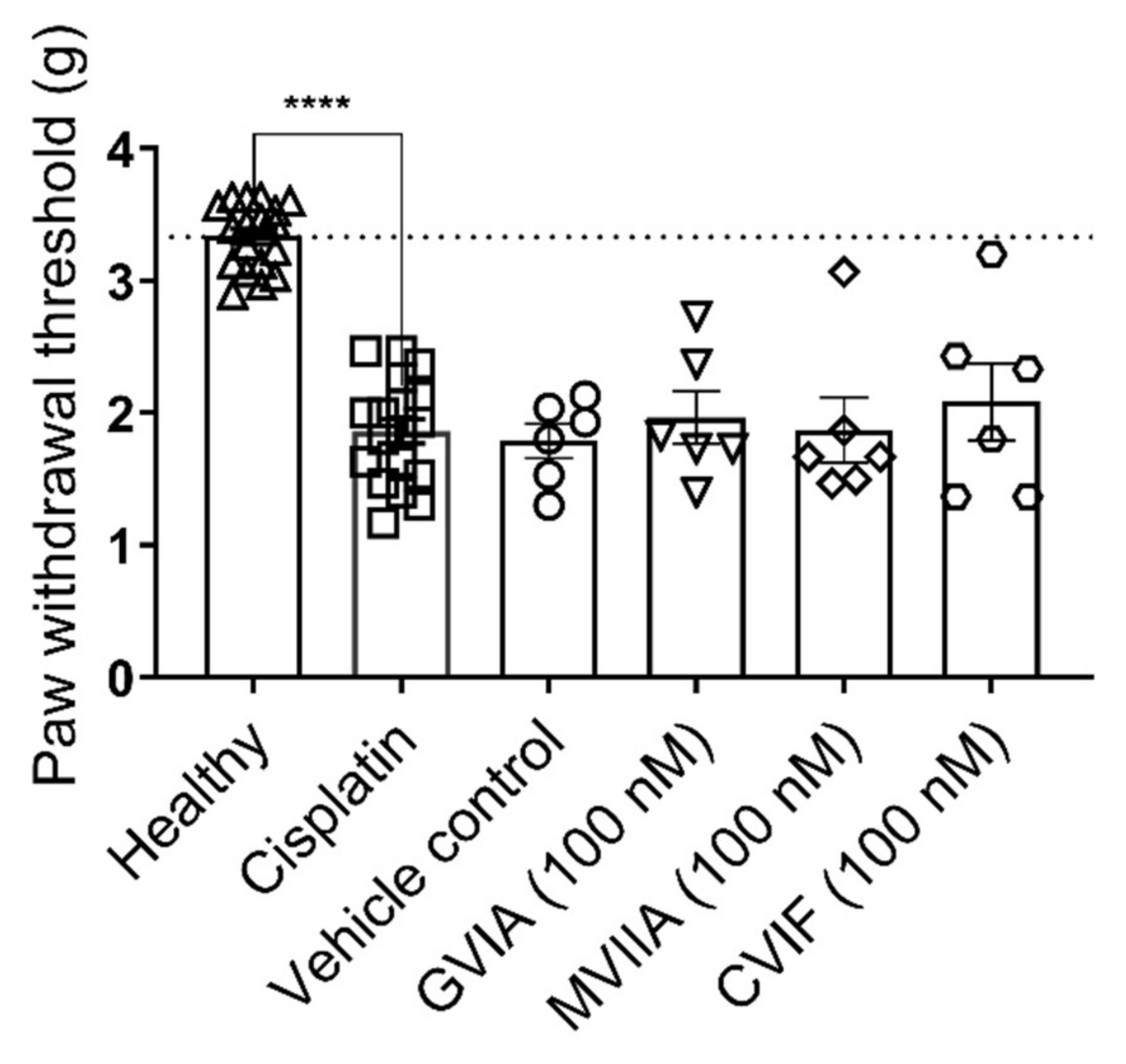
2.3.2. Effect of ω-Conotoxins in Cisplatin-Induced Mechanical Allodynia
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Animals
4.4. Fluorescence-Imaging Assays
4.5. Cisplatin- and Oxaliplatin-Induced Neuropathy
4.6. Postsurgical Pain
4.7. Mechanical Paw Withdrawal Threshold Measurements
4.8. Thermal Paw Withdrawal Threshold Measurements
4.9. Locomotor Performance Assessment
4.10. Treatments
4.11. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus venom peptide pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Seger, J.; Horvath, M.P.; Fedosov, A.E. Prey-capture strategies of fish-hunting cone snails: Behavior, neurobiology and evolution. Brain Behav. Evol. 2015, 86, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, C.I.; Lewis, R.J. ω-Conotoxins GVIA, MVIIA and CVID: SAR and clinical potential. Mar. Drugs 2006, 4, 193–214. [Google Scholar] [CrossRef] [Green Version]
- Gohil, K.; Bell, J.R.; Ramachandran, J.; Miljanich, G.P. Neuroanatomical distribution of receptors for a novel voltage-sensitive calcium-channel antagonist, SNX-230 (ω-conopeptide MVIIC). Brain Res. 1994, 653, 258–266. [Google Scholar] [CrossRef]
- Westenbroek, R.E.; Hell, J.W.; Warner, C.; Dubel, S.J.; Snutch, T.P.; Catterall, W.A. Biochemical properties and subcellular distribution of an N-type calcium hannel and α2δ subunit. Neuron 1992, 9, 1099–1115. [Google Scholar] [CrossRef]
- Catterall, W.A.; Perez-Reyes, E.; Snutch, T.P.; Striessnig, J. Nomenclature and structure-function relationships of voltage-gated calcium channels. Pharmacol. Rev. 2005, 57, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Bourinet, E.; Soong, T.W.; Sutton, K.; Slaymaker, S.; Mathews, E.; Monteil, A.; Zamponi, G.W.; Nargeot, J.; Snutch, T.P. Splicing of α1A subunit gene generates phenotypic variants of P- and Q-type calcium channels. Nature Neurosci. 1999, 2, 407–415. [Google Scholar] [CrossRef]
- Richards, K.S.; Swensen, A.M.; Lipscombe, D.; Bommert, K. Novel CaV2.1 clone replicates many properties of Purkinje cell CaV2.1 current. Eur. J. Neurosci. 2007, 26, 2950–2961. [Google Scholar] [CrossRef]
- Perez-Reyes, E. Molecular physiology of low-voltage-activated T-type calcium channels. Physiol. Rev. 2003, 83, 117–161. [Google Scholar] [CrossRef] [Green Version]
- Takemura, M.; Kiyama, H.; Fukui, H.; Tohyama, M.; Wada, H. Autoradiographic visualization in rat brain of receptors for ω-conotoxin GVIA, a newly discovered calcium antagonist. Brain Res. 1988, 451, 386–389. [Google Scholar] [CrossRef]
- Kerr, L.M.; Yoshikami, D. A venom peptide with a novel presynaptic blocking action. Nature 1984, 308, 282–284. [Google Scholar] [CrossRef]
- Miljanich, G.P. Ziconotide: Neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef]
- Olivera, B.M.; Gray, W.R.; Zeikus, R.; McIntosh, J.M.; Varga, J.; Rivier, J.; de Santos, V.; Cruz, L.J. Peptide neurotoxins from fish-hunting cone snails. Science 1985, 230, 1338–1343. [Google Scholar] [CrossRef]
- Atanassoff, P.G.; Hartmannsgruber, M.W.; Thrasher, J.; Wermeling, D.; Longton, W.; Gaeta, R.; Singh, T.; Mayo, M.; McGuire, D.; Luther, R.R. Ziconotide, a new N-type calcium channel blocker, administered intrathecally for acute postoperative pain. Reg. Anesth. Pain Med. 2000, 25, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.E.; Robertson, A.D.; Whorlow, S.L.; Angus, J.A. Cardiovascular and autonomic effects of ω-conotoxins MVIIA and CVID in conscious rabbits and isolated tissue assays. Br. J. Pharmacol. 2000, 131, 1325–1336. [Google Scholar] [CrossRef]
- Penn, R.D.; Paice, J.A. Adverse effects associated with the intrathecal administration of ziconotide. Pain 2000, 85, 291–296. [Google Scholar] [CrossRef]
- McGivern, J.G.; McDonough, S.I. Voltage-gated calcium channels as targets for the treatment of chronic pain. Curr. Drug Targets CNS Neurol. Disord. 2004, 3, 457–478. [Google Scholar] [CrossRef]
- Zamponi, G.W. Regulation of presynaptic calcium channels by synaptic proteins. J. Pharmacol. Sci. 2003, 92, 79–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wermeling, D.P. Ziconotide, an intrathecally administered N-type calcium channel antagonist for the treatment of chronic pain. Pharmacotherapy 2005, 25, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Malmberg, A.B.; Yaksh, T.L. Effect of continuous intrathecal infusion of ω-conopeptides, N-type calcium-channel blockers, on behavior and antinociception in the formalin and hot-plate tests in rats. Pain 1995, 60, 83–90. [Google Scholar] [CrossRef]
- Scott, D.A.; Wright, C.E.; Angus, J.A. Actions of intrathecal omega-conotoxins CVID, GVIA, MVIIA, and morphine in acute and neuropathic pain in the rat. Eur. J. Pharmacol. 2002, 451, 279–286. [Google Scholar] [CrossRef]
- Lewis, R.J.; Nielsen, K.J.; Craik, D.J.; Loughnan, M.L.; Adams, D.A.; Sharpe, I.A.; Luchian, T.; Adams, D.J.; Bond, T.; Thomas, L.; et al. Novel ω-conotoxins from Conus catus discriminate among neuronal calcium channel subtypes. J. Bol. Chem. 2000, 275, 35335–35344. [Google Scholar] [CrossRef] [Green Version]
- Berecki, G.; Motin, L.; Haythornthwaite, A.; Vink, S.; Bansal, P.; Drinkwater, R.; Wang, C.I.; Moretta, M.; Lewis, R.J.; Alewood, P.F.; et al. Analgesic ω-conotoxins CVIE and CVIF selectively and voltage-dependently block recombinant and native N-type calcium channels. Mol. Pharmacol. 2010, 77, 139–148. [Google Scholar] [CrossRef]
- Gan, T.J.; Habib, A.S.; Miller, T.E.; White, W.; Apfelbaum, J.L. Incidence, patient satisfaction, and perceptions of post-surgical pain: Results from a US national survey. Curr. Med. Res. Opin. 2014, 30, 149–160. [Google Scholar] [CrossRef]
- Gan, T.J. Poorly controlled postoperative pain: Prevalence, consequences, and prevention. J. Pain Res. 2017, 10, 2287–2298. [Google Scholar] [CrossRef] [Green Version]
- Sommer, M.; de Rijke, J.M.; van Kleef, M.; Kessels, A.G.; Peters, M.L.; Geurts, J.W.; Gramke, H.F.; Marcus, M.A. The prevalence of postoperative pain in a sample of 1490 surgical inpatients. Eur. J. Anaesth. 2008, 25, 267–274. [Google Scholar] [CrossRef]
- Banach, M.; Juranek, J.K.; Zygulska, A.L. Chemotherapy-induced neuropathies-a growing problem for patients and health care providers. Brain Behav. 2017, 7, e00558. [Google Scholar] [CrossRef] [PubMed]
- Starobova, H.; Vetter, I. Pathophysiology of chemotherapy-induced peripheral neuropathy. Front. Mol. Neurosci. 2017, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- de Moor, J.S.; Mariotto, A.B.; Parry, C.; Alfano, C.M.; Padgett, L.; Kent, E.E.; Forsythe, L.; Scoppa, S.; Hachey, M.; Rowland, J.H. Cancer survivors in the United States: prevalence across the survivorship trajectory and implications for care. Cancer Epidemiol. Biomarkers Prev. 2013, 22, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaplan, S.R.; Pogrel, J.W.; Yaksh, T.L. Role of voltage-dependent calcium channel subtypes in experimental tactile allodynia. J. Pharmacol. Exp. Therap. 1994, 269, 1117–1123. [Google Scholar] [PubMed]
- Wang, Y.X.; Gao, D.; Pettus, M.; Phillips, C.; Bowersox, S.S. Interactions of intrathecally administered ziconotide, a selective blocker of neuronal N-type voltage-sensitive calcium channels, with morphine on nociception in rats. Pain 2000, 84, 271–281. [Google Scholar] [CrossRef]
- Staats, P.S.; Yearwood, T.; Charapata, S.G.; Presley, R.W.; Wallace, M.S.; Byas-Smith, M.; Fisher, R.; Bryce, D.A.; Mangieri, E.A.; Luther, R.R.; et al. Intrathecal ziconotide in the treatment of refractory pain in patients with cancer or AIDS: A randomized controlled trial. J. Am. Med. Assoc. 2004, 291, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, D.M.; Cousins, M.J. Effect of subcutaneous administration of calcium channel blockers on nerve injury-induced hyperalgesia. Brain Res. 1998, 801, 50–58. [Google Scholar] [CrossRef]
- Mueller, A.; Starobova, H.; Morgan, M.; Dekan, Z.; Cheneval, O.; Schroeder, C.I.; Alewood, P.F.; Deuis, J.R.; Vetter, I. Antiallodynic effects of the selective NaV1.7 inhibitor Pn3a in a mouse model of acute postsurgical pain: Evidence for analgesic synergy with opioids and baclofen. Pain 2019, 160, 1766–1780. [Google Scholar] [CrossRef]
- Meller, S.T. Thermal and mechanical hyperalgesia: A distinct role for different excitatory amino acid receptors and signal transduction pathways? APS J. 1994, 3, 215–231. [Google Scholar] [CrossRef]
- Ishida, K.; Kawamata, T.; Tanaka, S.; Shindo, T.; Kawamata, M. Calcitonin gene-related peptide is involved in inflammatory pain but not in postoperative pain. Anesthesiology 2014, 121, 1068–1079. [Google Scholar] [CrossRef] [Green Version]
- Spofford, C.M.; Brennan, T.J. Gene expression in skin, muscle, and dorsal root ganglion after plantar incision in the rat. Anesthesiology 2012, 117, 161–172. [Google Scholar] [CrossRef] [Green Version]
- Hamalainen, M.M.; Subieta, A.; Arpey, C.; Brennan, T.J. Differential effect of capsaicin treatment on pain-related behaviors after plantar incision. J. Pain 2009, 10, 637–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barabas, M.E.; Stucky, C.L. TRPV1, but not TRPA1, in primary sensory neurons contributes to cutaneous incision-mediated hypersensitivity. Mol. Pain 2013, 9, 1744–8069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchytilova, E.; Spicarova, D.; Palecek, J. TRPV1 antagonist attenuates postoperative hypersensitivity by central and peripheral mechanisms. Mol. Pain 2014, 10, 10–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eastman, A. Alkylating and platinum-based agents. Curr. Opin. Oncol. 1990, 2, 1109–1114. [Google Scholar] [CrossRef]
- Rosenberg, B. Noble metal complexes in cancer chemotherapy. In Inorganic and Nutritional Aspects of Cancer; Advances in Experimental Medicine and Biology; Schrauzer, G.N., Ed.; Springer: Boston, MA, USA, 1977. [Google Scholar]
- Reed, E. Alkylating agents and platinum: Is clinical resistance simply a tumor cell phenomenon? Curr. Opin. Oncol. 1991, 3, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Starobova, H.; Mueller, A.; Deuis, J.R.; Carter, D.A.; Vetter, I. Inflammatory and neuropathic gene expression signatures of chemotherapy-induced neuropathy induced by vincristine, cisplatin, and oxaliplatin in C57BL/6J mice. J. Pain 2020, 21, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Joseph, E.K.; Levine, J.D. Comparison of oxaliplatin- and cisplatin-induced painful peripheral neuropathy in the rat. J. Pain 2009, 10, 534–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cersosimo, R.J. Oxaliplatin-associated neuropathy: a review. Ann. Pharmacother. 2005, 39, 128–135. [Google Scholar] [CrossRef]
- Extra, J.M.; Marty, M.; Brienza, S.; Misset, J.L. Pharmacokinetics and safety profile of oxaliplatin. Semin. Oncol. 1998, 25, 13–22. [Google Scholar] [PubMed]
- Deuis, J.R.; Zimmermann, K.; Romanovsky, A.A.; Possani, L.D.; Cabot, P.J.; Lewis, R.J.; Vetter, I. An animal model of oxaliplatin-induced cold allodynia reveals a crucial role for NaV1.6 in peripheral pain pathways. Pain 2013, 154, 1749–1757. [Google Scholar] [CrossRef] [Green Version]
- Sadeghi, M.; Murali, S.S.; Lewis, R.J.; Alewood, P.F.; Mohammadi, S.; Christie, M.J. Novel ω-conotoxins from C. catus reverse signs of mouse inflammatory pain after systemic administration. Mol. Pain 2013, 9, 1744. [Google Scholar] [CrossRef] [Green Version]
- Sann, S.B.; Xu, L.; Nishimune, H.; Sanes, J.R.; Spitzer, N.C. Neurite outgrowth and in vivo sensory innervation mediated by a CaV2.2-laminin β2 stop signal. J. Neurosci. 2008, 28, 2366–2374. [Google Scholar] [CrossRef] [Green Version]
- Barzan, R.; Pfeiffer, F.; Kukley, M. N- and L-type voltage-gated calcium channels mediate fast calcium transients in axonal shafts of mouse peripheral nerve. Front. Cell. Neurosci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Gold, M.S.; White, D.M.; Ahlgren, S.C.; Guo, M.; Levine, J.D. Catecholamine-induced mechanical sensitization of cutaneous nociceptors in the rat. Neurosci. Lett. 1994, 175, 166–170. [Google Scholar] [CrossRef]
- Taiwo, Y.O.; Heller, P.H.; Levine, J.D. Characterization of distinct phospholipases mediating bradykinin and noradrenaline hyperalgesia. Neuroscience 1990, 39, 523–531. [Google Scholar] [CrossRef]
- Adams, D.J.; Smith, A.B.; Schroeder, C.I.; Yasuda, T.; Lewis, R.J. ω-Conotoxin CVID inhibits a pharmacologically distinct voltage-sensitive calcium channel associated with transmitter release from preganglionic nerve terminals. J. Biol. Chem. 2003, 278, 4057–4062. [Google Scholar] [CrossRef] [Green Version]
- Mould, J.; Yasuda, T.; Schroeder, C.I.; Beedle, A.M.; Doering, C.J.; Zamponi, G.W.; Adams, D.J.; Lewis, R.J. The α2δ auxiliary subunit reduces affinity of ω-conotoxins for recombinant N-type (Cav2.2) calcium channels. J. Biol. Chem. 2004, 279, 34705–34714. [Google Scholar] [CrossRef] [Green Version]
- Newton, R.A.; Bingham, S.; Case, P.C.; Sanger, G.J.; Lawson, S.N. Dorsal root ganglion neurons show increased expression of the calcium channel α2δ subunit following partial sciatic nerve injury. Brain Res. Mol. Brain Res. 2001, 95, 1–8. [Google Scholar] [CrossRef]
- Costigan, M.; Befort, K.; Karchewski, L.; Griffin, R.S.; D'Urso, D.; Allchorne, A.; Sitarski, J.; Mannion, J.W.; Pratt, R.E.; Woolf, C.J. Replicate high-density rat genome oligonucleotide microarrays reveal hundreds of regulated genes in the dorsal root ganglion after peripheral nerve injury. BMC Neurosci. 2002, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Li, C.-Y.; Song, Y.-H.; Higuera, E.S.; Luo, Z.D. Spinal dorsal horn calcium channel α2δ1 subunit upregulation contributes to peripheral nerve injury-induced tactile allodynia. J. Neurosci. 2004, 24, 8494–8499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, C.S.; Nieto-Rostro, M.; Rahman, W.; Tran-Van-Minh, A.; Ferron, L.; Douglas, L.; Kadurin, I.; Sri Ranjan, Y.; Fernandez-Alacid, L.; Millar, N.S.; et al. The increased trafficking of the calcium channel subunit α2δ to presynaptic terminals in neuropathic pain is inhibited by the α2δ ligand pregabalin. J. Neurosci. 2009, 29, 4076–4088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jay, S.D.; Sharp, A.H.; Kahl, S.D.; Vedvick, T.S.; Harpold, M.M.; Campbell, K.P. Structural characterization of the dihydropyridine-sensitive calcium channel α2δ subunit and the associated δ peptides. J. Biol. Chem. 1991, 266, 3287–3293. [Google Scholar] [CrossRef]
- Ellinor, P.T.; Zhang, J.F.; Horne, W.A.; Tsien, R.W. Structural determinants of the blockade of N-type calcium channels by a peptide neurotoxin. Nature 1994, 372, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.P.; Doering, C.J.; Winkfein, R.J.; Beedle, A.M.; Spafford, J.D.; Zamponi, G.W. Determinants of inhibition of transiently expressed voltage-gated calcium channels by ω-conotoxins GVIA and MVIIA. J. Biol. Chem. 2003, 278, 20171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarotskyy, V.; Elmslie, K.S. ω-Conotoxin GVIA alters gating charge movement of N-type (CaV2.2) calcium channels. J. Neurophysiol. 2009, 101, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The Future of peptide-based drugs. Chem. Biol. Drug Des. 2013, 81, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Degim, I.T.; Celebi, N. Controlled delivery of peptides and proteins. Curr. Pharm. Des. 2007, 13, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Deuis, J.R.; Lim, Y.L.; Rodrigues de Sousa, S.; Lewis, R.J.; Alewood, P.F.; Cabot, P.J.; Vetter, I. Analgesic effects of clinically used compounds in novel mouse models of polyneuropathy induced by oxaliplatin and cisplatin. Neuro-Oncology 2014, 16, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Deuis, J.R.; Dvorakova, L.S.; Vetter, I. Methods used to evaluate pain behaviors in rodents. Front. Mol. Neurosci. 2017, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Deuis, J.R.; Wingerd, J.S.; Winter, Z.; Durek, T.; Dekan, Z.; Sousa, S.R.; Zimmermann, K.; Hoffmann, T.; Weidner, C.; Nassar, M.A.; et al. Analgesic effects of GpTx-1, PF-04856264 and CNV1014802 in a mouse model of NaV1.7-mediated pain. Toxins 2016, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Kiehn, O. Decoding the organization of spinal circuits that control locomotion. Nat. Rev. Neurosci. 2016, 17, 224–238. [Google Scholar] [CrossRef]
- Deuis, J.R.; Vetter, I. The thermal probe test: A novel behavioral assay to quantify thermal paw withdrawal thresholds in mice. Temperature 2016, 3, 199–207. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, M.M.; Starobova, H.; Mueller, A.; Vetter, I.; Lewis, R.J. Subcutaneous ω-Conotoxins Alleviate Mechanical Pain in Rodent Models of Acute Peripheral Neuropathy. Mar. Drugs 2021, 19, 106. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020106
Hasan MM, Starobova H, Mueller A, Vetter I, Lewis RJ. Subcutaneous ω-Conotoxins Alleviate Mechanical Pain in Rodent Models of Acute Peripheral Neuropathy. Marine Drugs. 2021; 19(2):106. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020106
Chicago/Turabian StyleHasan, Md. Mahadhi, Hana Starobova, Alexander Mueller, Irina Vetter, and Richard J. Lewis. 2021. "Subcutaneous ω-Conotoxins Alleviate Mechanical Pain in Rodent Models of Acute Peripheral Neuropathy" Marine Drugs 19, no. 2: 106. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020106