
Isoquinolinequinone Derivatives from a Marine Sponge (Haliclona sp.) Regulate Inflammation in In Vitro System of Intestine

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
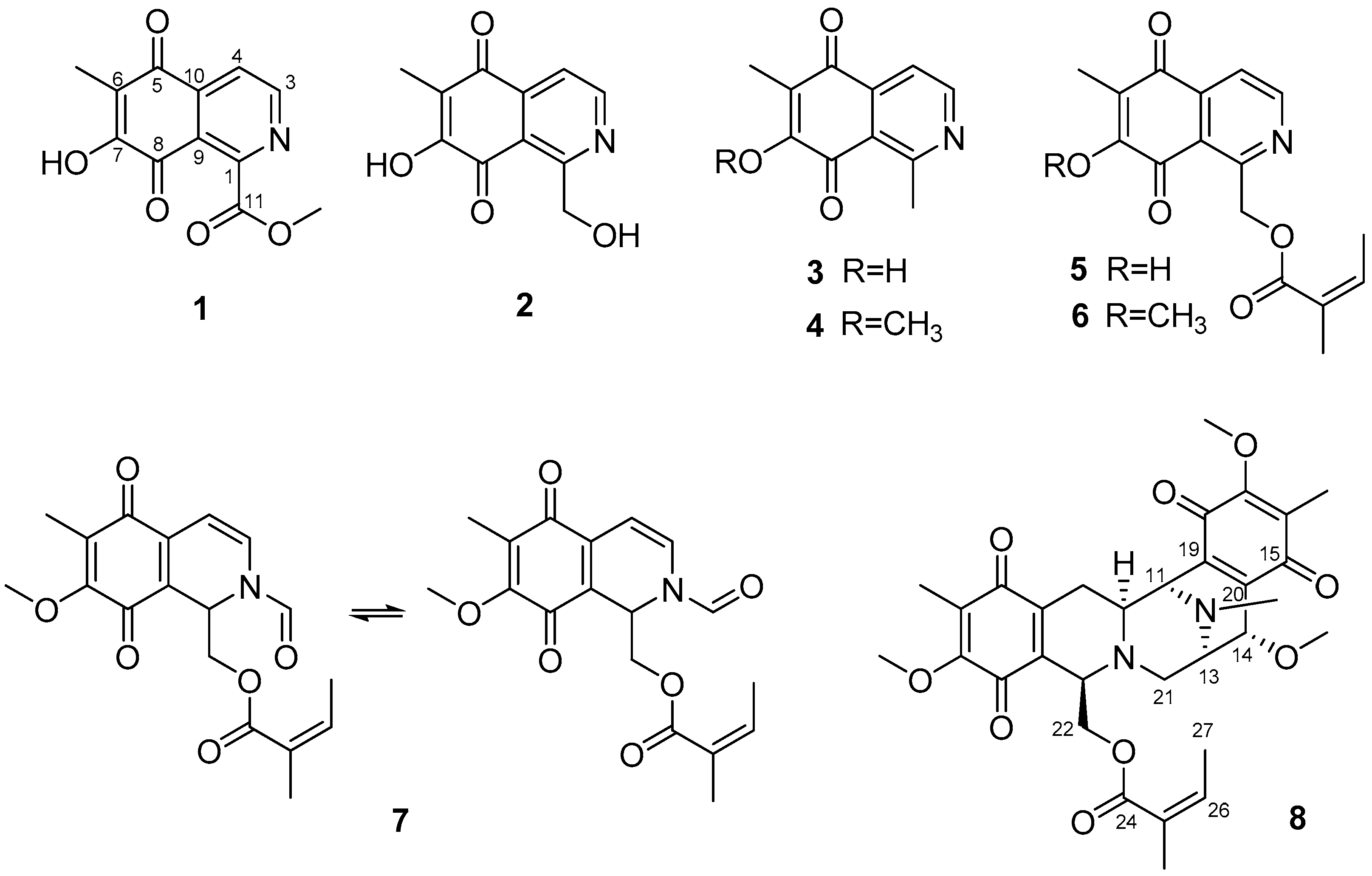
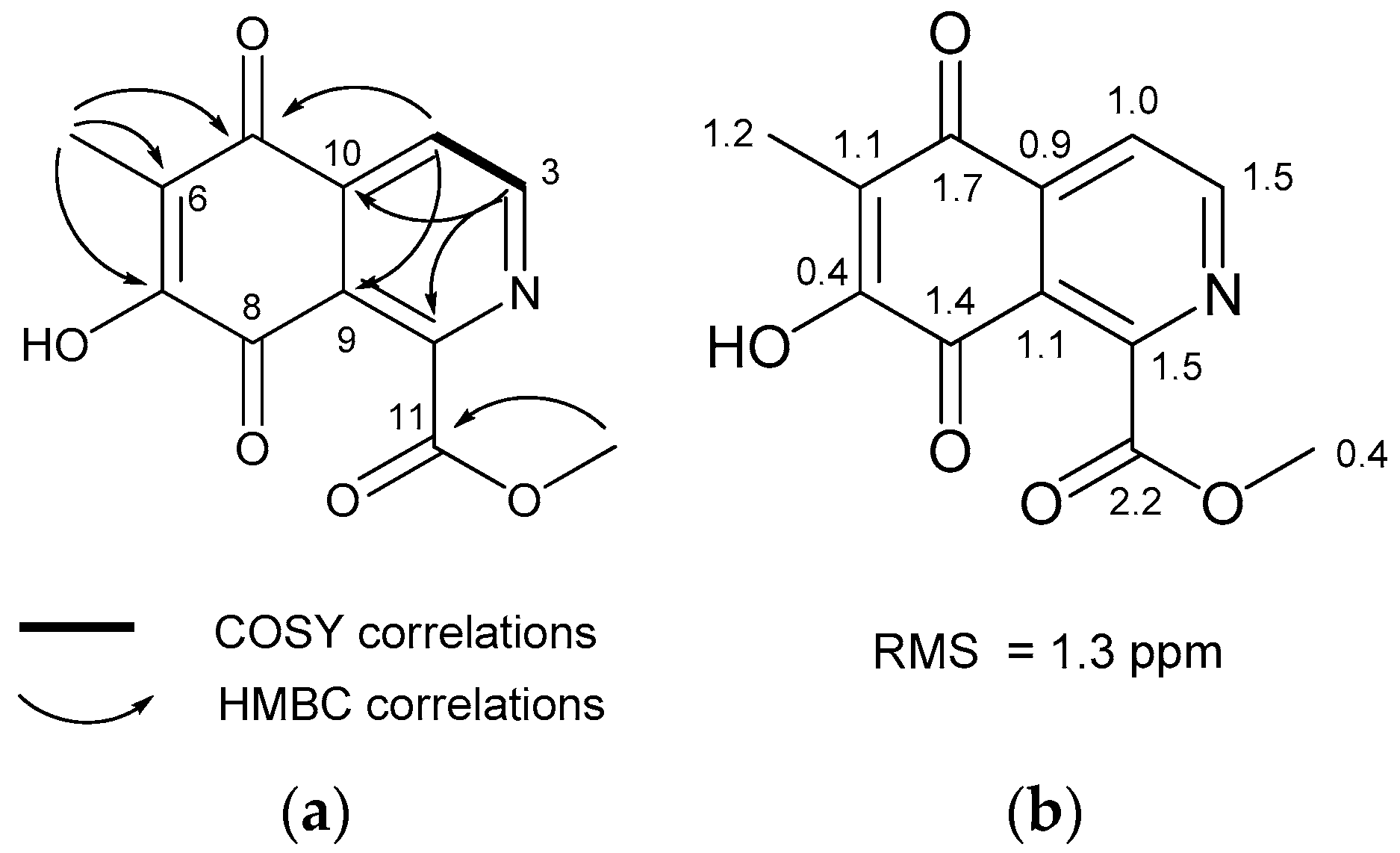
2.1. Structures of Compounds 1–3 and 8 Isolated from Haliclona sp.
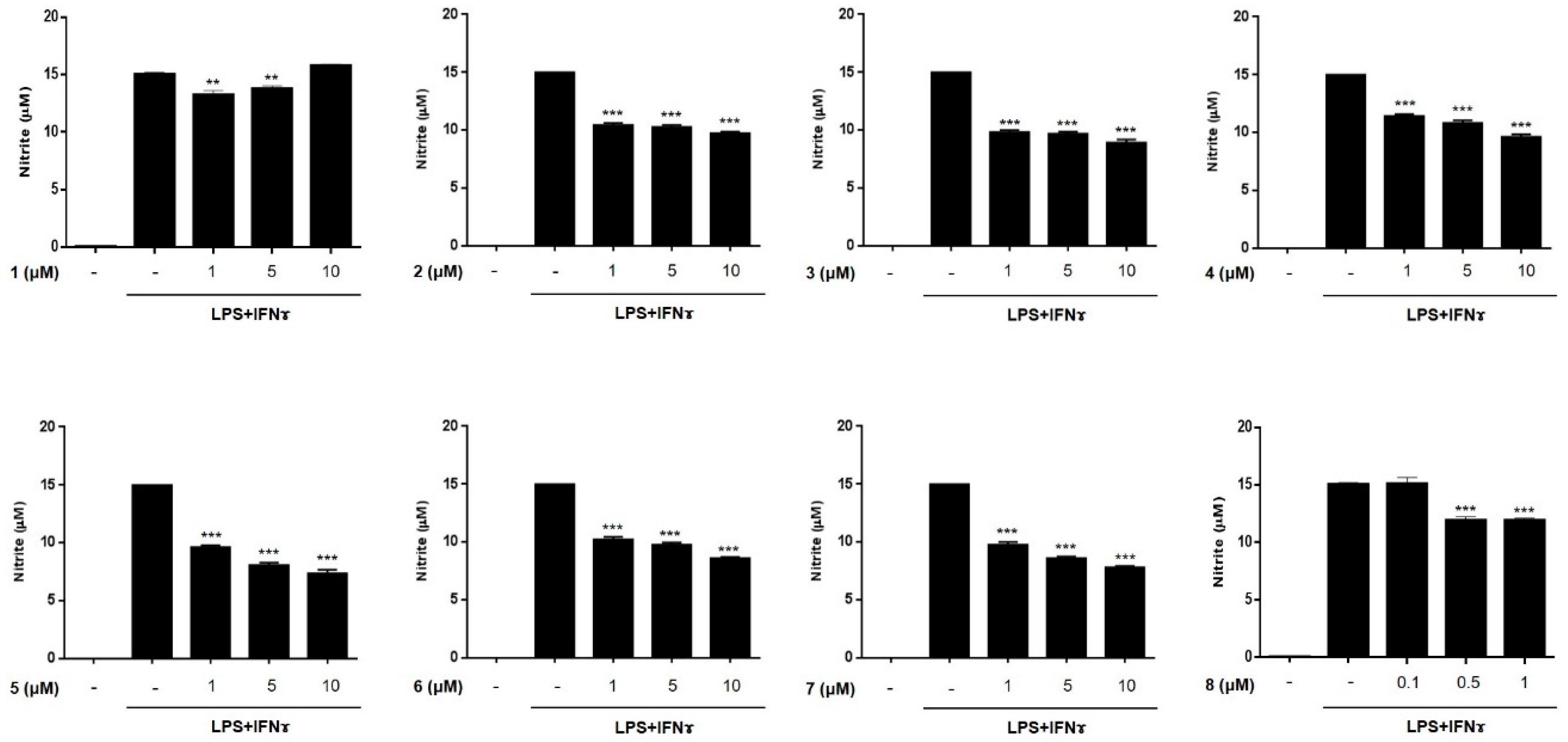
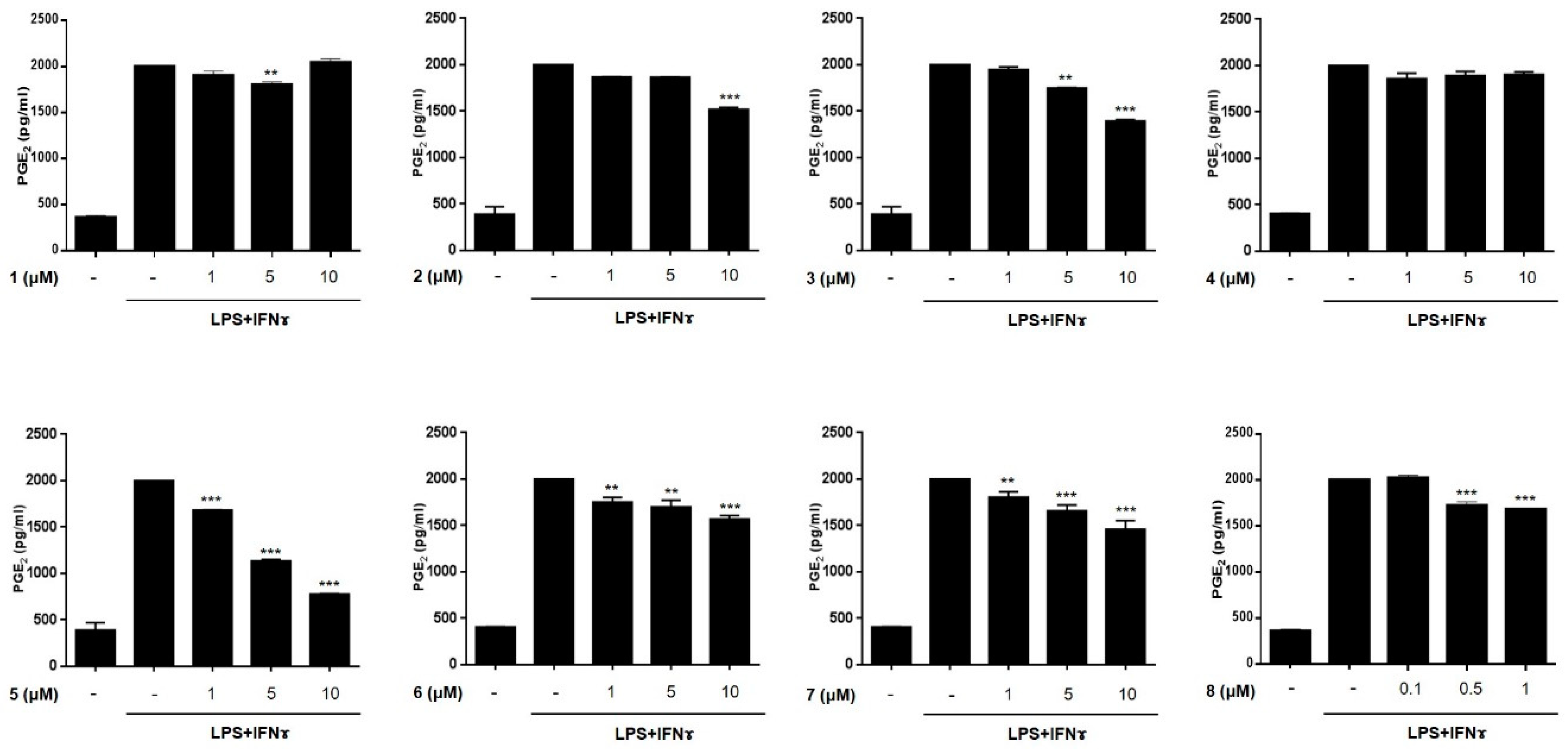
2.2. Anti-Inflammatory Effects of Compounds 1–8 in THP-1 Macrophages Activated Using Lipopolysacchride (LPS) and IFNγ
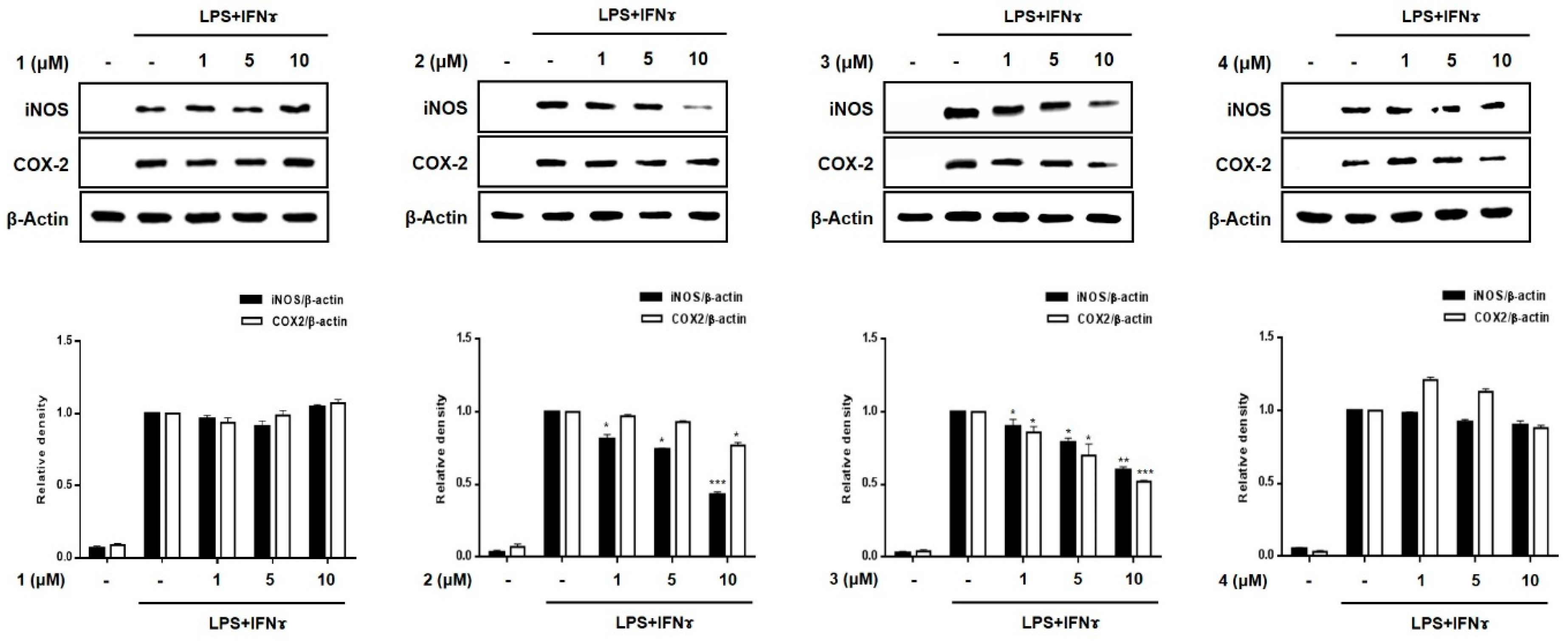
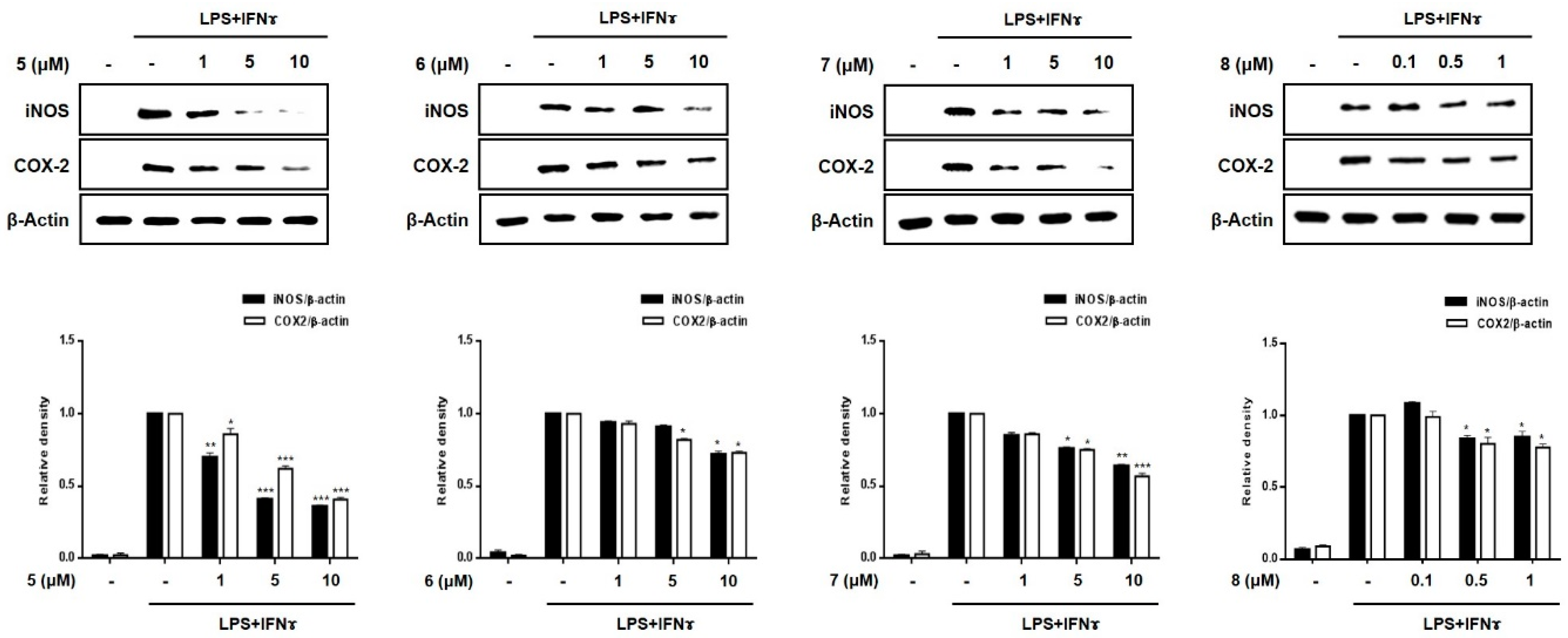
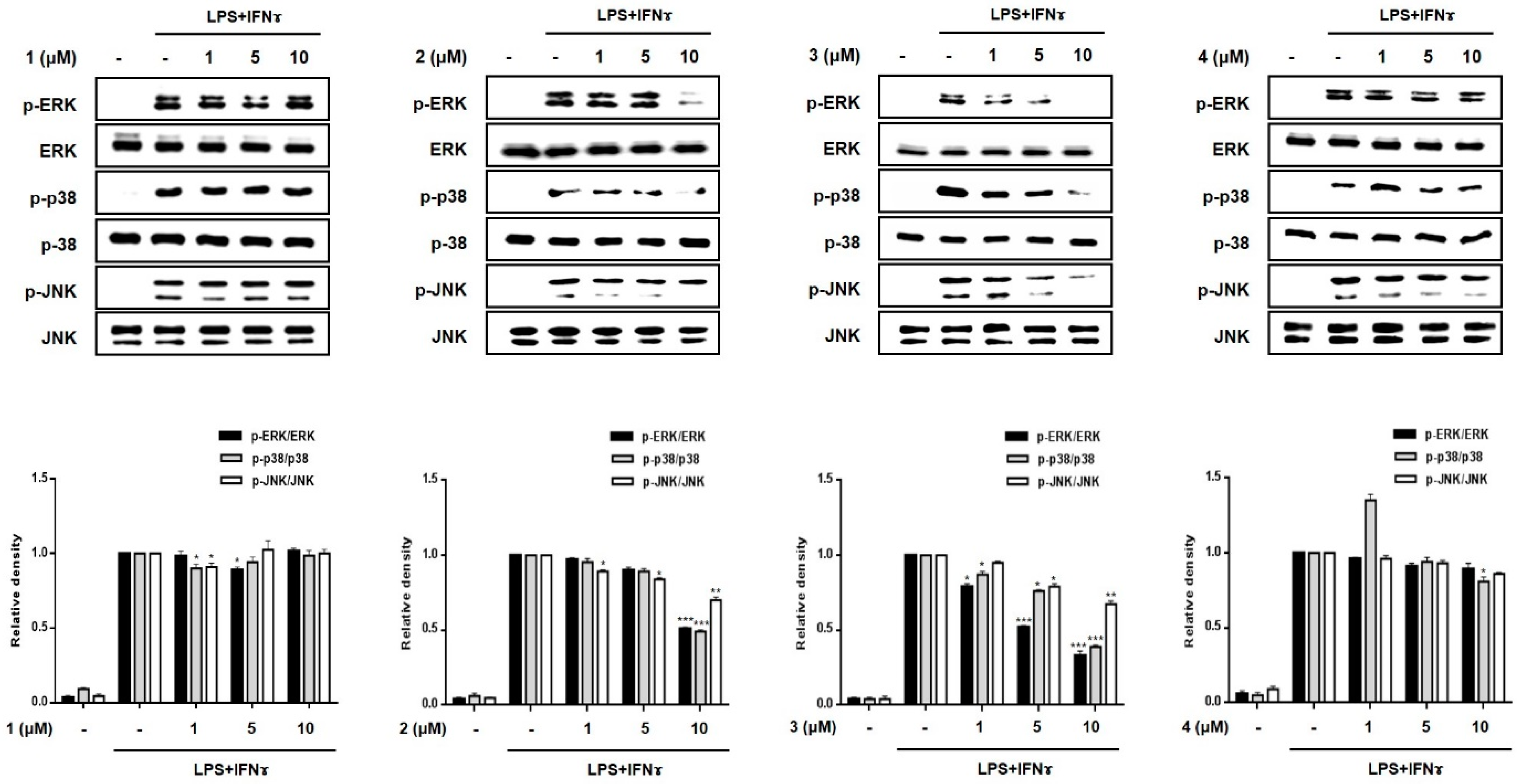
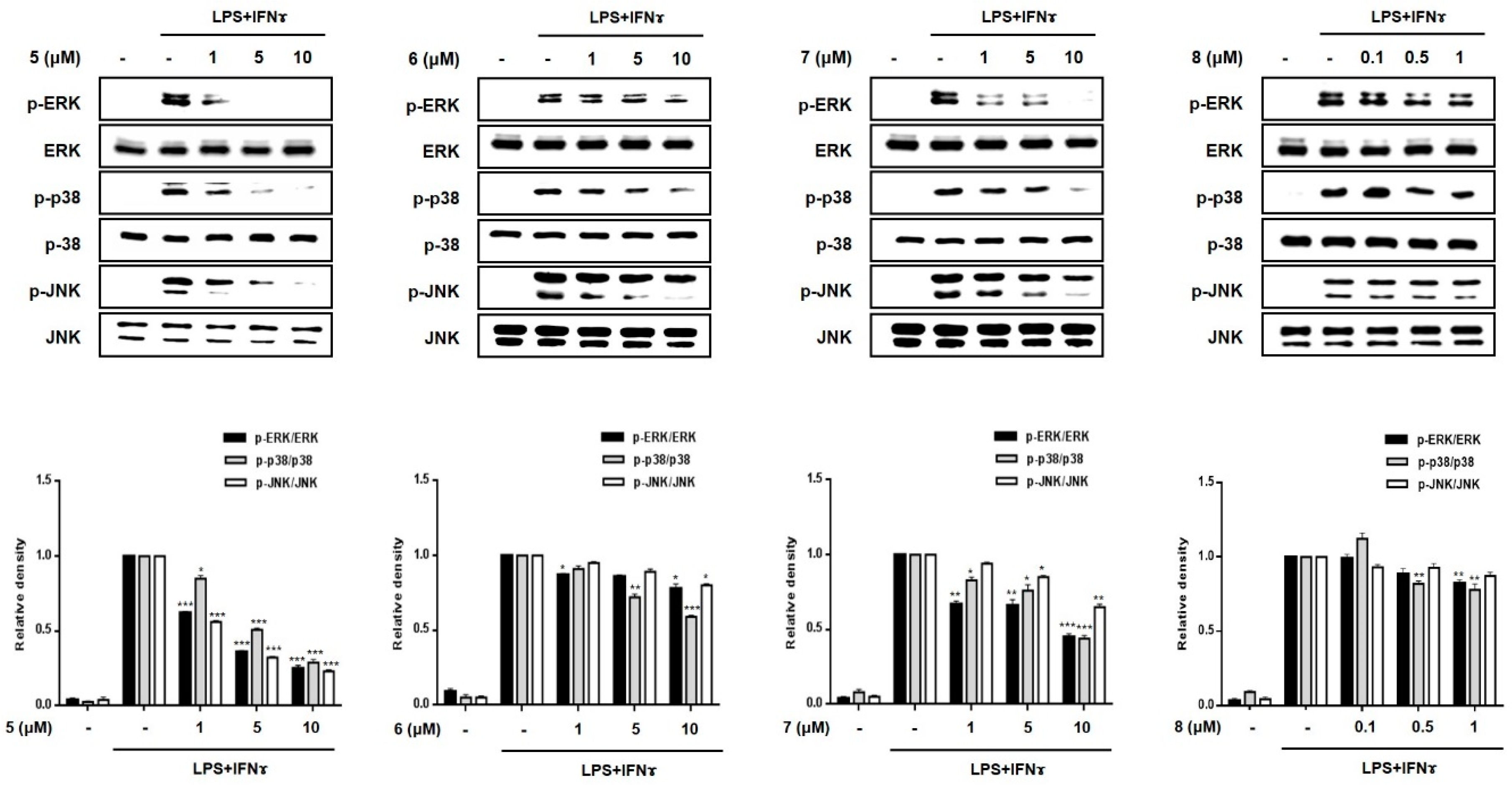
2.3. Effects of Compounds 1–8 on Pro-Inflammatory Protein Expression and MAPK Phosphorylation in THP-1 Macrophages Co-Cultured with Caco-2 Cells
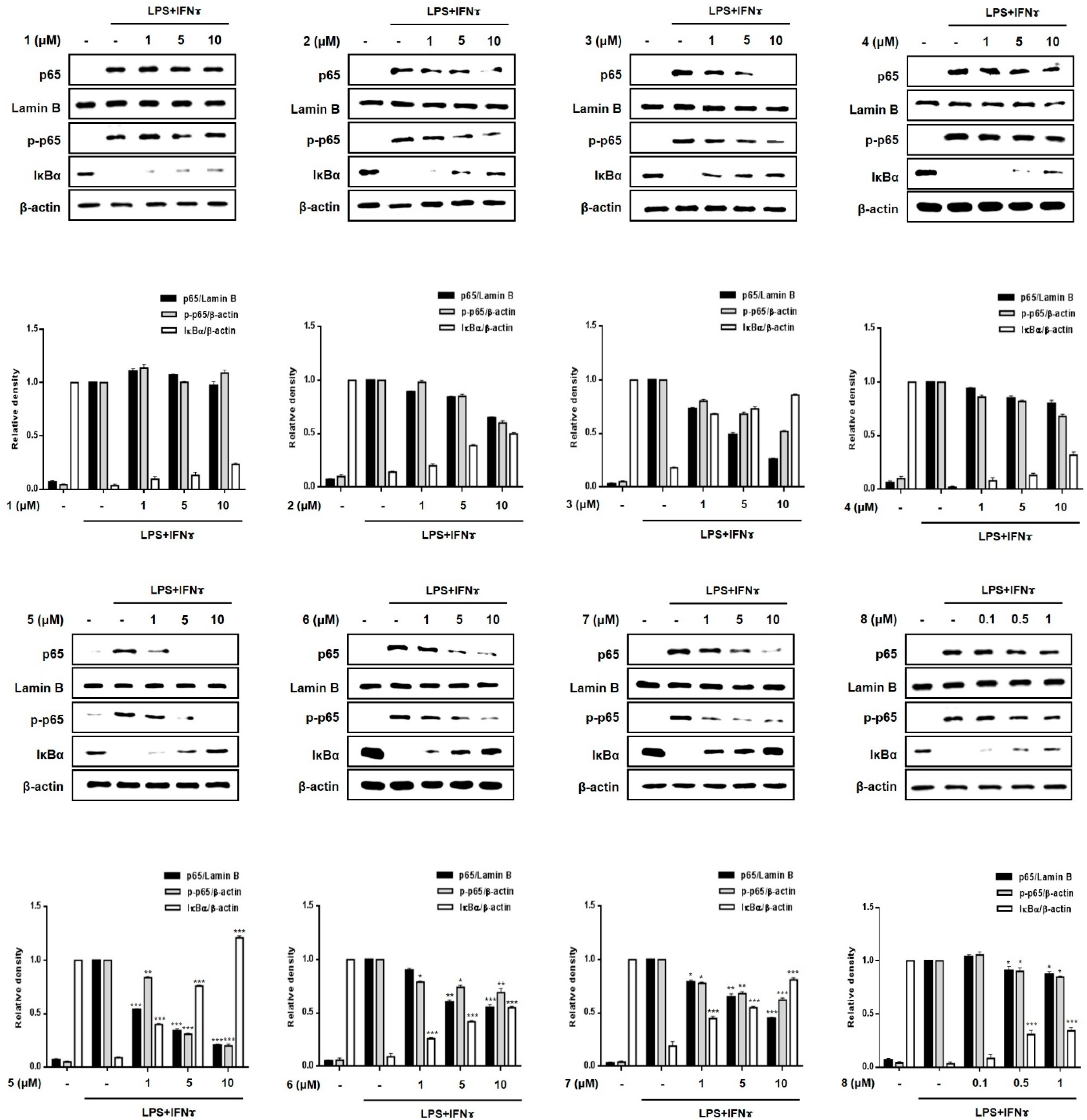
2.4. Effects of Compounds 1–8 on the Nuclear Translocation of NF-κB and Its Inhibitor, IκB-α
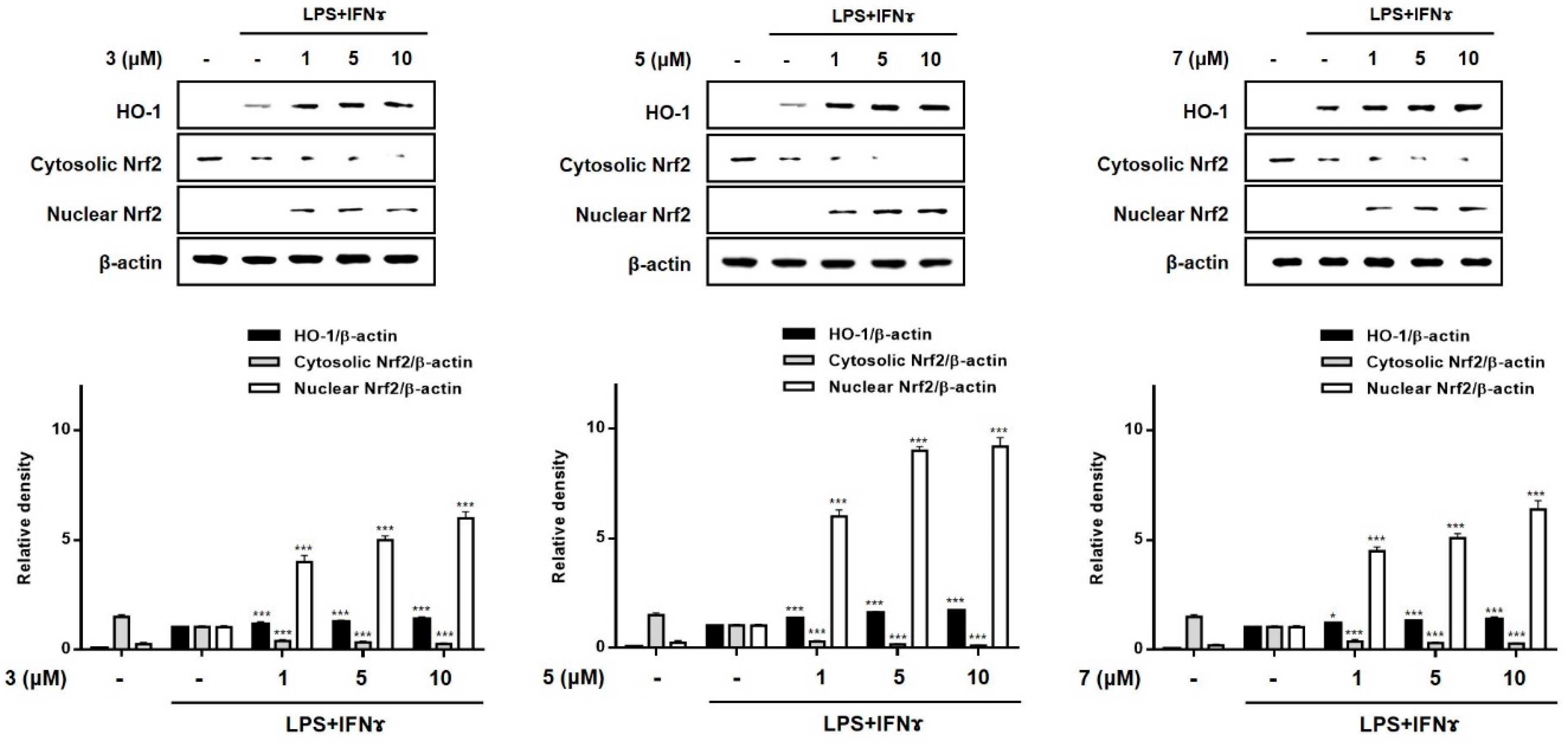
2.5. Effects of Compounds 3, 5, and 7 on the Expression of HO-1 in THP-1 Macrophages Co-Cultured with Caco-2 Cells
2.6. Structure-Activity Relationship of the Haliclona sp. Isoquinolinequinone Derivatives
3. Materials and Methods
3.1. General Procedures
3.2. Materials
3.3. Animal Material
3.4. Isolation of Isoquinolinequinones from Haliclona sp.
3.5. Quantum Calculations of 13C NMR Chemical Shifts
3.6. Cell Cultures
3.7. Differentiation of THP-1 to Macrophages
3.8. In Vitro Intestinal Co-Culture Model
3.9. TEER Measurement
3.10. Cell Viability Assay
3.11. Measurement of NO and PGE2 Production
3.12. Western Blot
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine Natural Products. Nat. Prod. Rep. 2020, 37, 175. [Google Scholar] [CrossRef] [PubMed]
- Perdicaris, S.; Vlachogianni, T.; Valavanidis, A. Bioactive Natural Substances from Marine Sponges: New Developments and Prospects for Future Pharmaceuticals. Nat. Prod. Chem. Res. 2013, 1, 3. [Google Scholar] [CrossRef]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine Sponge Derived Natural Products between 2001 and 2010: Trends and Opportunities for Discovery of Bioactives. Mar. Drugs 2014, 12, 4539. [Google Scholar] [CrossRef] [Green Version]
- Edrada, R.A.; Proksch, P.; Wray, V.; Christ, R.; Witte, L.; Soest, R.W.M.V. Bioactive Isoquinoline Quinone from an Undescribed Philippine Marine Sponge pf the Genus Xestospongia. J. Nat. Prod. 1996, 59, 973. [Google Scholar] [CrossRef]
- Standoval, I.T.; Davis, R.A.; Bugni, T.S.; Concepcion, G.P.; Harper, M.K.; Ireland, C.M. Cytotoxic Isoquinoline quinones from Sponge of the Genus Petrosia. Nat. Prod. Res. 2004, 18, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntyre, D.E.; Faulkner, D.J.; Engen, D.V.; Clardy, J. Renierone, an antimicrobial metabolites from a marine sponge. Tetrahedron Lett. 1979, 43, 4163. [Google Scholar] [CrossRef]
- Koh, S.-I.; Shin, H.-S. The Anti-Rotaviral and Anti-Inflammatory Effects of Hyrtios and Haliclona Species. J. Microbiol. Biotechnol. 2016, 26, 2006. [Google Scholar] [CrossRef]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [Green Version]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Kucharzik, T.; Maaser, C.; Lügering, A.; Kagnoff, M.; Mayer, L.; Targan, S.; Domschke, W. Recent understanding of IBD pathogenesis: Implications for future therapies. Inflamm. Bowel Dis. 2006, 12, 1068–1083. [Google Scholar] [CrossRef] [PubMed]
- Kleiveland, C.R. Chapter 18: Co-culture Caco-2/Immune Cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Eds.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Abraham, C.; Cho, J.H. Inflammatory bowel disease. N. Engl. J. Med. 2009, 361, 2066–2078. [Google Scholar] [CrossRef]
- Kämpfer, A.A.M.; Urbán, P.; Gioria, S.; Kanase, N.; Stone, V.; Kinsner-Ovaskainen, A. Development of an in vitro co-culture model to mimic the human intestine in healthy and diseased state. Toxicol. In Vitro 2017, 45, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Rubas, W.; Jezyk, N.; Grass, G.M. Comparison of the permeability characteristics of a human colonic epithelial (Caco-2) cell line to colon of rabbit, monkey, and dog intestine and human drug absorption. Pharm. Res. 1993, 10, 113–118. [Google Scholar] [CrossRef]
- Lennernas, H.; Palm, K.; Fagerholm, U.; Artursson, P. Comparison between active and passive drug transport in human intestinal epithelial (caco-2) cells in vitro and human jejunum in vivo. Int. J. Pharm. 1996, 127, 103–107. [Google Scholar] [CrossRef]
- Frincke, J.M.; Faulkner, J. Antimicrobial metabolites of the sponge Reniera sp. J. Am. Chem. Soc. 1982, 104, 265. [Google Scholar] [CrossRef]
- Mckee, T.C.; Ireland, C.M. Cytotoxic and antimicrobial alkaloids from the Fijian sponge Xestospongia calcedoi. J. Nat. Prod. 1987, 50, 754. [Google Scholar] [CrossRef] [PubMed]
- He, H.-Y.; Faulkner, D.J. Renieramycins E and F from the sponge Reniera sp.: Reassignments of the stereochemistry of the renieramycins. J. Org. Chem. 1989, 54, 5822. [Google Scholar] [CrossRef]
- Pettus, B.J.; Bielawski, J.; Porcelli, A.M.; Reames, D.L.; Johnson, K.R.; Morrow, J.; Chalfant, C.E.; Obeid, L.M.; Hannun, Y.A. The sphingosine kinase 1/sphingosine-1-phosphate pathway mediates COX-2 induction and PGE2 production in response to TNF-alpha. FASEB J. 2003, 17, 1411–1421. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.H.; Hsieh, M.C.; Hsu, P.C. 6-Shogaol suppressed lipopolysaccharide-induced up-expression of iNOS and COX-2 in murine macrophages. Mol. Nutr. Food Res. 2008, 52, 1467–1477. [Google Scholar] [CrossRef]
- Wilson, K.T.; Ramanujam, K.S.; Mobley, H.L.; Musselman, R.F.; James, S.P.; Meltzer, S.J. Helicobacter pylori stimulates inducible nitric oxide synthase expression and activity in a murine macrophage cell line. Gastroenterology 1996, 111, 1524–1533. [Google Scholar] [CrossRef]
- Reinecker, H.C.; Steffen, M.; Withoeft, T.; Pflueger, I.; Schreiber, S.; MacDermott, R.P.; Raedler, A. Enhanced secretion of tumour necrosis factor-alpha, IL-6, and IL-1 beta by isolated lamina propria mononuclear cells from patients with ulcerative colitis and Crohn’s disease. Clin. Exp. Immunol. 1993, 94, 174–181. [Google Scholar] [CrossRef]
- Lappas, M.; Permezel, M.; Georgiou, H.M.; Rice, G.E. Nuclear factor kappa B regulation of proinflammatory cytokines in human gestational tissues in vitro. Biol. Reprod. 2002, 67, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Takada, Y.; Shishodia, S.; Gutierrez, A.M.; Oommen, O.V.; Ichikawa, H. Nuclear transcription factor NF-kappa B: Role in biology and medicine. Indian J. Exp. Biol. 2004, 42, 341–353. [Google Scholar] [PubMed]
- Economou, J.S.; Rhoades, R.; Essner, W.H.; McBride, J.C.; Morton, D.L. Genetic analysis of the human tumor necrosis factor a/cachectin promoter region in a macrophage cell line. J. Exp. Med. 1989, 170, 321. [Google Scholar] [CrossRef] [PubMed]
- Appleby, S.B.; Ristimäki, A.; Neilson, K.; Narko, K.; Hla, T. Structure of the human cyclo-oxygenase-2 gene. Biochem. J. 1994, 302, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, I.T.; Won, J.H. Anti-inflammatory activities of ent16alphaH,17-hydroxy-kauran-19-oic acid isolated from the roots of Siegesbeckia pubescens are due to the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via NF-kappaB inactivation. Eur. J. Pharmacol. 2007, 558, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Saegusa, M.; Hashimura, M.; Kuwata, T. Pin1 acts as a modulator of cell proliferation through alteration in NF-kappaB but not beta-catenin/TCF4 signalling in a subset of endometrial carcinoma cells. J. Pathol. 2010, 222, 410–420. [Google Scholar] [CrossRef]
- Otterbein, L.E.; Bach, F.H.; Alam, J.; Soares, M.; Tao, L.H.; Wysk, M.; Davis, R.J.; Flavell, R.A.; Choi, A.M. Carbon monoxide has antiinflammatory effects involving the mitogen-activated protein kinase pathway. Nat. Med. 2000, 6, 422–428. [Google Scholar] [CrossRef]
- Lee, D.S.; Li, B.; Im, N.K.; Kim, Y.C.; Jeong, G.S. 4,2′,5′-Trihydroxy4′-methoxychalcone from Dalbergia odorifera exhibits anti-inflammatory properties by inducing heme oxygenase-1 in murine macrophage. Int. Immunopharmacol. 2013, 16, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xu, C.; Liu, D.; Han, M.K.; Wang, L.; Merlin, D. Oral delivery of nanoparticles loaded with ginger active compound, 6-Shogaol, attenuates ulcerative colitis and promotes wound healing in a murine model of ulcerative colitis. J. Crohns Colitis 2018, 12, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.Y.; Chen, W.T.; Kurtan, T.; Mandi, A.; Ding, J.; Li, J.; Li, X.W.; Guo, Y.W. Bioactive isoquinolinequinone alkaloids from the South China Sea nudibranch Jorunna funebris and its sponge-prey Xestospongia sp. Future Med. Chem. 2016, 8, 17–27. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16, Revision C. 01; Gaussian, Inc.: Wallingford, CT, USA, 2019. [Google Scholar]
- Starr, T.; Bauler, T.J.; Malik-Kale, P.; Steele-Mortimer, O. The phorbol 12-myristate-13-acetate differentiation protocol is critical to the interaction of THP-1 macrophages with Salmonella Typhimurium. PLoS ONE 2018, 13, e0193601. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, N.H.; Ji, Y.K.; Kim, Y.N.; Jeon, Y.-J.; Heo, J.D.; Jeong, E.J.; Rho, J.-R. Sulfoquinovosylmonoacylglycerols regulating intestinal inflammation in co-culture system from the brown alga Turbinaria ornata. Algae 2020, 35, 201–212. [Google Scholar] [CrossRef]
- Lee, S.M.; Kim, N.H.; Lee, S.B.; Kim, Y.N.; Heo, J.D.; Jeong, E.J.; Rho, J.-R. Deacetylphylloketal, a New Phylloketal Derivative from a Marine Sponge, Genus Phyllospongia, with Potent Anti-Inflammatory Activity in In Vitro Co-Culture Model of Intestine. Mar. Drugs 2019, 17, 634. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | 3 | |||
|---|---|---|---|---|---|---|
| 13C, mult | 1H, m(J Hz) | 13C, mult | 1H, m(J Hz) | 13C, mult | 1H, m(J Hz) | |
| 1 | 150.6, C | 160.4, C | 160.5, C | |||
| 3 | 155.6, CH | 9.04, d(4.9) | 154.1, CH | 9.00, d(4.9) | 155.1, CH | 8.91, d(4.9) |
| 4 | 120.5, CH | 8.06, d(4.9) | 118.9, CH | 7.99, d(4.9) | 118.0, CH | 7.88, d(4.9) |
| 5 | 182.8, C | 183.6, C | 184.0, C | |||
| 6 | 121.9, C | 120.4, C | 121.0, C | |||
| 7 | 153.4, C | 153.5, C | 153.7, C | |||
| 8 | 179.2, C | 181.1, C | 181.6, C | |||
| 9 | 120.1, C | 119.7, C | 119.7, C | |||
| 10 | 139.0, C | 140.1, C | 140.0, C | |||
| 11 | 166.6, C | 64.0, CH2 | 5.22, d(4.2) | 25.6, CH3 | 3.02, s | |
| 6-CH3 | 8.8, CH3 | 2.15, s | 8.6, CH3 | 2.12, s | 8.5, CH3 | 2.11, s |
| OCH3 | 53.4, CH3 | 4.08, s | ||||
| Compounds (10 μM) | NO | PGE2 |
|---|---|---|
| Relative Content (%) vs. LPS + IFN-γ-Treated Cells | ||
| 1 | 100.0 | 102.1 |
| 2 | 63.3 | 76.5 |
| 3 | 59.4 | 70.6 |
| 4 | 64.2 | 95.4 |
| 5 | 49.0 | 39.1 |
| 6 | 57.7 | 78.3 |
| 7 | 52.6 | 73.6 |
| 8 * | 80.2 | 84.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.N.; Ji, Y.K.; Kim, N.-H.; Van Tu, N.; Rho, J.-R.; Jeong, E.J. Isoquinolinequinone Derivatives from a Marine Sponge (Haliclona sp.) Regulate Inflammation in In Vitro System of Intestine. Mar. Drugs 2021, 19, 90. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020090
Kim YN, Ji YK, Kim N-H, Van Tu N, Rho J-R, Jeong EJ. Isoquinolinequinone Derivatives from a Marine Sponge (Haliclona sp.) Regulate Inflammation in In Vitro System of Intestine. Marine Drugs. 2021; 19(2):90. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020090
Chicago/Turabian StyleKim, Yun Na, Yeong Kwang Ji, Na-Hyun Kim, Nguyen Van Tu, Jung-Rae Rho, and Eun Ju Jeong. 2021. "Isoquinolinequinone Derivatives from a Marine Sponge (Haliclona sp.) Regulate Inflammation in In Vitro System of Intestine" Marine Drugs 19, no. 2: 90. https://0-doi-org.brum.beds.ac.uk/10.3390/md19020090