
Modeling Occurrence of Urban Mosquitos Based on Land Use Types and Meteorological Factors in Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Data
2.2. Environmental Data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Variables | Abbreviation | Mean (±SD *) |
|---|---|---|---|
| Land use types | Residential area (%) | RESD | 28.4 (±23.8) |
| Commercial area (%) | COMM | 16.2 (±13.2) | |
| Culture and sport area (%) | CULT | 7.9 (±13.5) | |
| Public facilities area (%) | PUBL | 8.5 (±11.8) | |
| Artificial grassland (%) | GRAS | 7.5 (±10.3) | |
| Inland wetland (%) | WETL | 0.0 (±0.1) | |
| Bare soil (%) | BARE | 2.6 (±4.1) | |
| Inland water (%) | WATE | 5.0 (±6.4) | |
| Industrial area (%) | INDU | 0.1 (±0.3) | |
| Traffic area (%) | TRAF | 23.8 (±6.7) | |
| Meteorological factors | Average daily temperature (°C) | TempAVE | 21.4 (±4.7) |
| Maximum daily temperature (°C) | TempMAX | 25.6 (±4.6) | |
| Minimum daily temperature (°C) | TempMIN | 17.7 (±5.2) | |
| Average daily wind speed (m/s) | WindAVE | 1.4 (±0.7) | |
| Average daily precipitation (mm) | PrcpAVE | 0.3 (±1.1) | |
| Average daily humidity (%) | HumiAVE | 70.6 (±13.3) | |
| Total daily precipitation (mm) | PrcpTOT | 5.7 (±22.6) |
2.3. Data Analysis
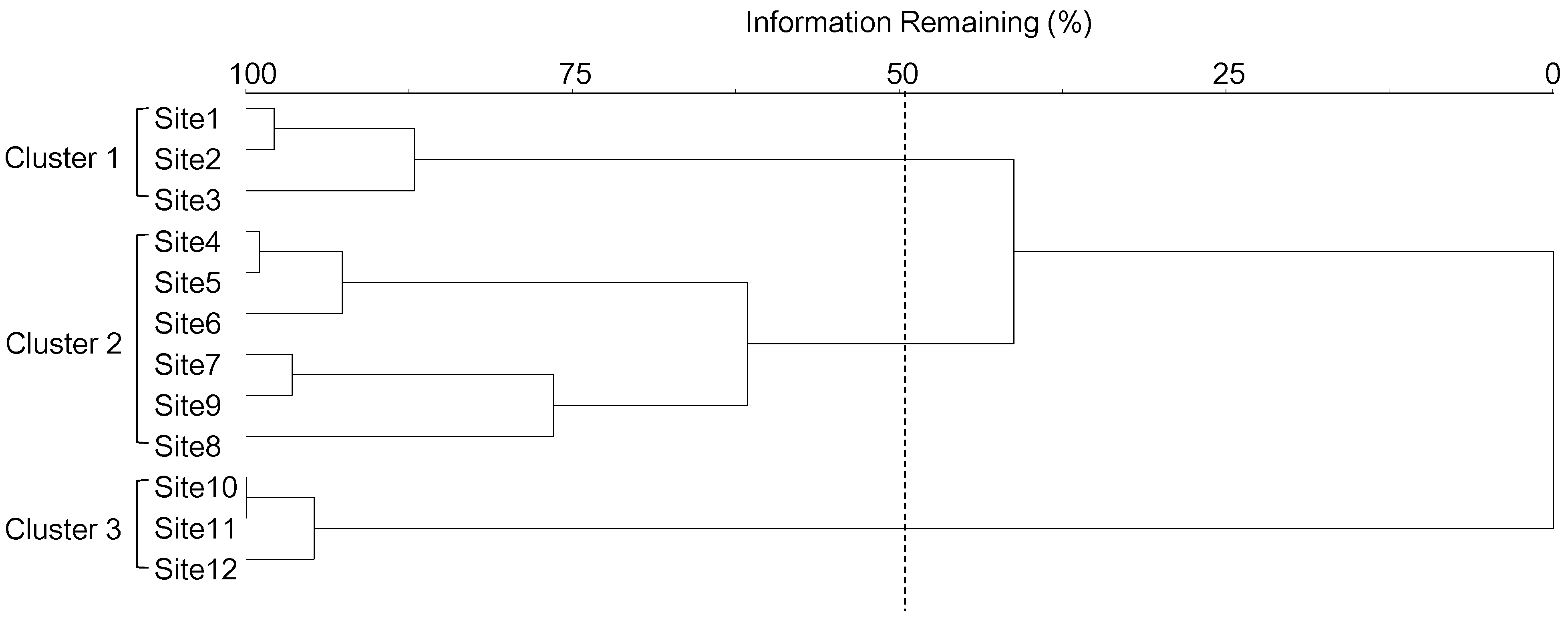
2.3.1. Classification of Habitats

2.3.2. Prediction of Mosquito Occurrence
2.3.3. Statistical Analysis
| Abundance Category | Cluster | ||
|---|---|---|---|
| 1 | 2 | 3 | |
| A (<25%) | ≤21.6 | ≤1.0 | ≤1.8 |
| B (25–50%) | 21.6–63.3 | 1.0–2.9 | 1.8–6 |
| C (50–75%) | 63.3–221.4 | 2.9–8.9 | 6–15.8 |
| D (>75%) | >221.4 | >8.93 | >15.8 |
3. Results
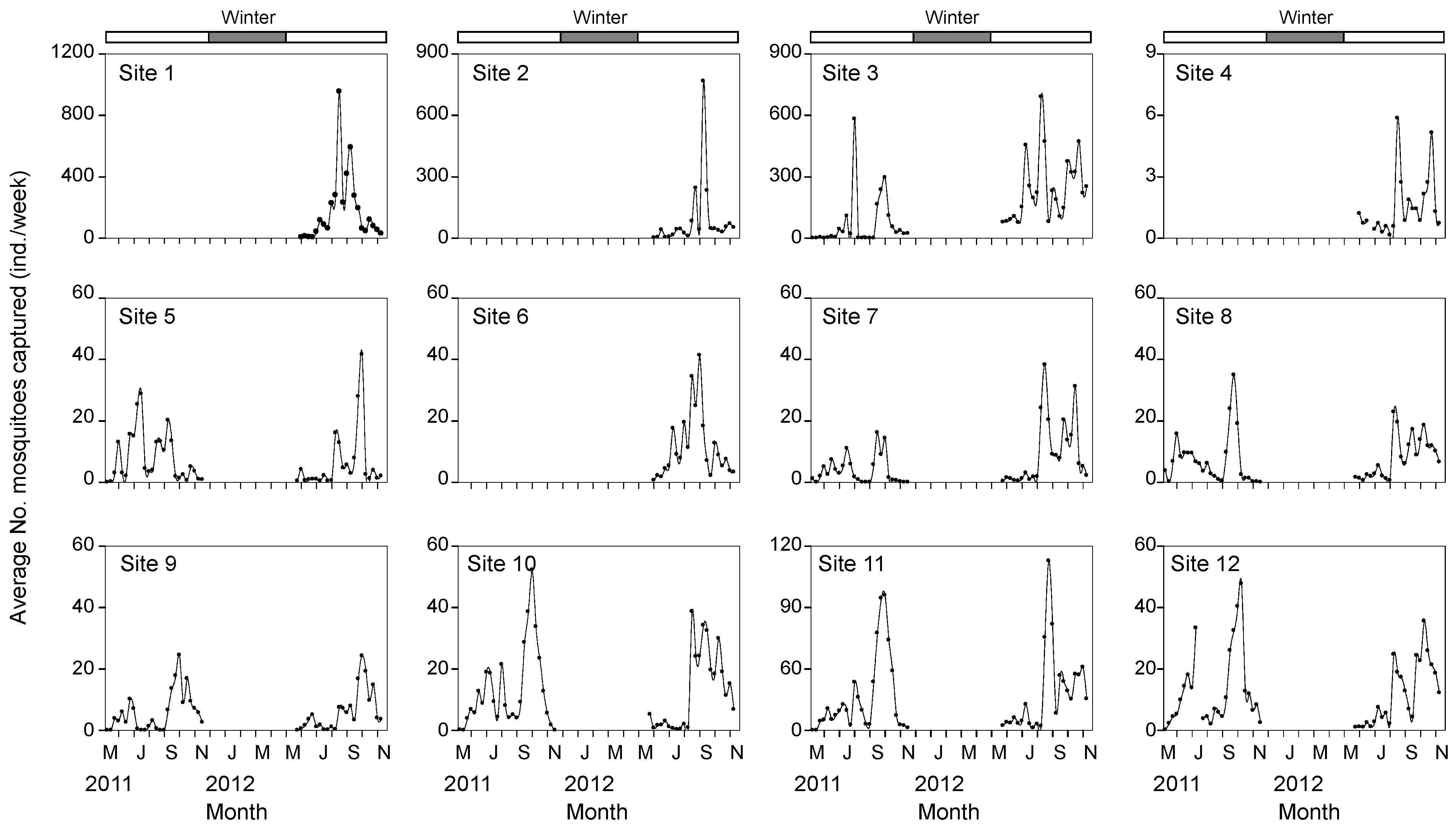
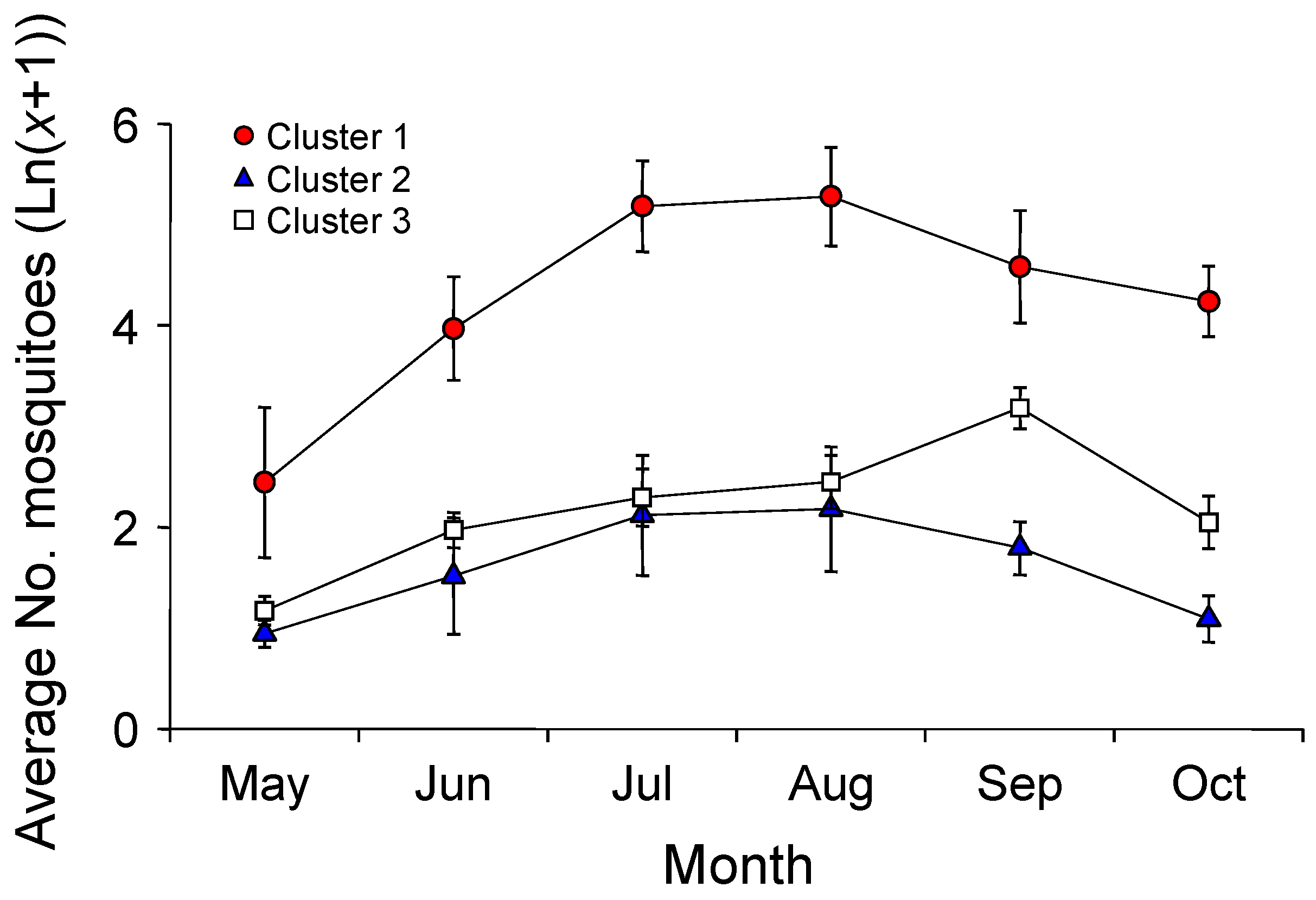
3.1. Mosquito Occurrence


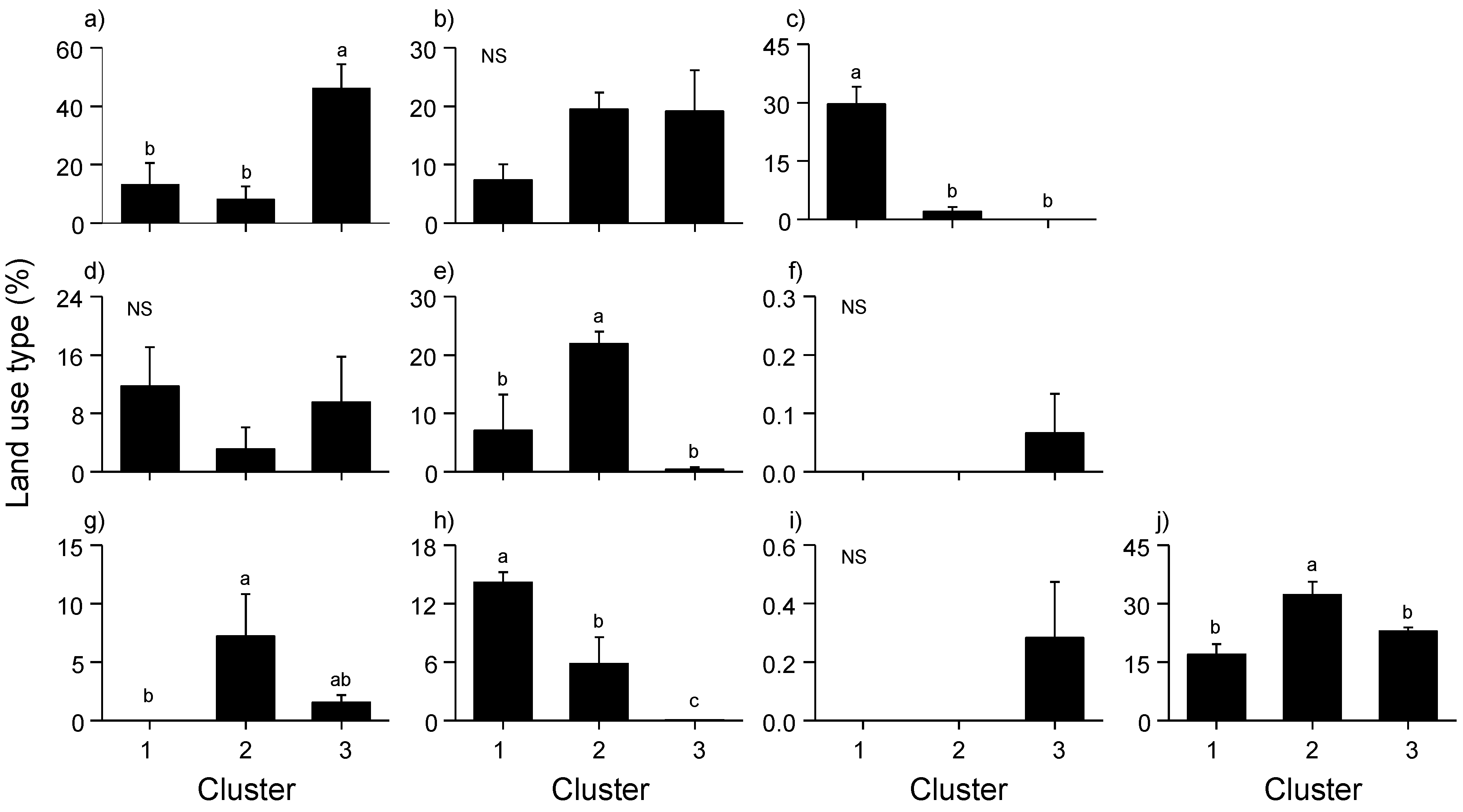
3.2. Relationships between Mosquito Abundance and Land Use Types



3.3. Prediction of Mosquito Occurrence

| Dataset | Model | Cluster | ||
|---|---|---|---|---|
| 1 | 2 | 3 | ||
| Four categories of abundance (A–D) | SVM | 0.59 | 0.61 | 0.52 |
| CT | 0.61 | 0.67 | 0.67 | |
| RF | 0.8 | 0.71 | 0.71 | |
| Category with highest abundance (D) | RF | 0.86 | 0.88 | 0.88 |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and their control; Springer: London, UK, 2010. [Google Scholar]
- Mulla, M.S. Mosquito control then, now, and in the future. J. Am. Mosq. Contr. 1994, 190, 574–584. [Google Scholar]
- Maguire, M.; Skelly, C.; Weinstein, P.; Moloney, J. Simulation modelling of Aedes aegypti prevalence, an environmental hazard surveillance tool for the control of dengue epidemics. Int. J. Env. Health Res. 1999, 9, 253–259. [Google Scholar] [CrossRef]
- Rose, R.I. Pesticides and public health: Integrated methods of mosquito management. Emerg. Infect. Diseases 2001, 7, 17–23. [Google Scholar] [CrossRef]
- Hay, S.I.; Cox, J.; Rogers, D.J.; Randolph, S.E.; Stern, D.I.; Shanks, D.W.; Myers, M.F.; Snow, R.W. Climate change and the resurgence of malaria in the East African highlands. Nature 2002, 415, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.M.; Knight, T.M. Drought-induced mosquito outbreaks in wetlands. Ecol. Lett. 2003, 6, 1017–1024. [Google Scholar] [CrossRef]
- Hamer, G.L.; Kitron, U.D.; Brawn, J.D.; Loss, S.R.; Ruix, M.O.; Goldberg, T.L.; Walker, E.D. Culex pipiens (Diptera: Culicidae): A bridge vector of West Nile virus to humans. J. Med. Entomol. 2008, 45, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.C.; Scott, T.W.; Lerdthusnee, K.; Coleman, R.C.; Costero, A.; Clark, G.G.; Jones, J.J.; Kitthawee, S.; Kittayapong, P.; Sithiprasasna, R.; Edman, J.D. Dispersal of the dengue vector Aedes aegypti within and between rural communities. Am. J. Trop. Med. Hyg. 2005, 72, 209–220. [Google Scholar] [PubMed]
- Barrett, A.D.; Higgs, S. Yellow fever: A disease that has yet to be conquered. Annu. Rev. Entomol. 2007, 52, 209–229. [Google Scholar] [CrossRef] [PubMed]
- Ryan, P.A.; Do, K.A.; Kay, B.H. Spatial and temporal analysis of Ross River virus disease patterns at Maroochy Shire, Australia: Association between human morbidity and mosquito (Diptera: Culicidae) abundance. J. Med. Entomol. 1999, 36, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Tong, S.; Mengersen, K.; Oldenburg, B.; Dale, P. Mosquito species (Diptera: Culicidae) and the transmission of Ross River virus in Brisbane, Australia. J. Med. Entomol. 2006, 43, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Udevitz, M.S.; Bloomfield, P.; Apperson, C.S. Prediction of the occurrence of four species of mosquito larvae with logistic regression on water-chemistry variables. Environ. Entomol. 1987, 16, 281–285. [Google Scholar] [CrossRef]
- Hales, S.; De Wet, N.; Maindonald, J.; Woodward, A. Potential effect of population and climate changes on global distribution of dengue fever: An empirical model. The Lancet 2002, 360, 830–834. [Google Scholar] [CrossRef]
- Peterson, A.T.; Martínez-Campos, C.; Nakazawa, Y.; Martínez-Meyer, E. Time-specific ecological niche modeling predicts spatial dynamics of vector insects and human dengue cases. T. Roy. Soc. Trop. Med. Hyg. 2005, 99, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Kearney, M.; Porter, W.P.; Williams, C.; Ritchie, S.; Hoffmann, A.A. Integrating biophysical models and evolutionary theory to predict climatic impacts on species’ ranges: The dengue mosquito Aedes aegypti in Australia. Funct. Ecol. 2009, 23, 528–538. [Google Scholar] [CrossRef]
- Ruiz, M.O.; Chaves, L.F.; Hamer, G.L.; Sun, T.; Brown, W.M.; Walker, E.D.; Haramis, L.; Goldberg, T.L.; Kitron, U.D. Local impact of temperature and precipitation on West Nile virus infection in Culex species mosquitoes in northeast Illinois, USA. Parasit Vect. 2010, 3. [Google Scholar] [CrossRef] [PubMed]
- Turchin, P. Population regulation: Old arguments and a new synthesis. In Population Dynamics: New Approaches and Synthesis; Cappuccino, N., Price, P., Eds.; Academic Press: San Diego, CA, USA, 1995; pp. 19–40. [Google Scholar]
- Barbraud, C.; Weimerskirch, H. Climate and density shape population dynamics of a marine top predator. Proc. R. Soc. Lond. B 2003, 270, 2111–2116. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Brook, B.W.; Bradshaw, C.J.A. Predicting the timing and magnitude of tropical mosquito population peaks for maximizing control efficiency. PLoS Neglected Tropic. Dis., 2009, 3. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.J.; Brook, B.W.; Whelan, P.I.; Cleland, S.; Bradshaw, C.J.A. Endogenous and exogenous factors controlling temporal abundance patterns of tropical disease-carrying mosquitoes. Ecol. Appl. 2008, 18, 2028–2040. [Google Scholar] [CrossRef] [PubMed]
- Vandyk, J.K.; Rowley, W.A. Response of Iowa mosquito populations to unusual precipitation patterns as measured by New Jersey light trap collections. J. Am. Mosq. Contr. 1995, 11, 200–205. [Google Scholar]
- Wegbreit, J.; Reisen, W.K. Relationships among weather, mosquito abundance, and encephalitis virus activity in California: Kern County 1990–1998. J. Am. Mosq. Control 2000, 16, 22–27. [Google Scholar]
- Russell, R.C. Mosquito-borne arboviruses in Australia: The current scene and implications of climate change for human health. Int. J. Parasitol. 1998, 28, 955–969. [Google Scholar] [CrossRef]
- Lindsay, S.W.; Adiamah, J.H.; Miller, J.E.; Pleass, R.J.; Armstrong, J.R.M. Variation in attractiveness of human subjects to malaria mosquitoes (Diptera: Culicidae) in The Gambia. J. Med. Entomol. 1993, 30, 368–368. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, J.S.; Lindsay, M.D.; Broom, A.K. Climate change and vector-borne diseases: Potential conequences for human health. In Health in the Greenhouse: The Medical and Environmental Health Effects of Global Climate Change; Ewan, C.E., Calvert, G.D., Bryant, E.A., Garrick, J.A., Eds.; Aust. Govt. Publ. Serv.: Canberra, Australia, 1993; pp. 229–234. [Google Scholar]
- Jeong, Y.S.; Lee, D.K. Prevalence and seasonal abundance of the dominant mosquito species in a large marsh near coast of Ulsan. Korean J. Appl. Entomol. 2003, 42, 125–132. [Google Scholar]
- Ha, Y.B. Habitats and seasonal prevalence of mosquitoes at dredged soil dumping and near area sites of Busan new port, Republic of Korea. Ph.D. thesis, Kosin University, Busan, South Korea, 2010. [Google Scholar]
- Epstein, P.R. Climate change and infectious disease: Stormy weather ahead? Epidemiology 2002, 13, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.; Malone, J.; Harb, M.; Faris, R.; Huh, O.; Buck, A. Bancroftian filariasis distribution and diurnal temperature differences in the southern Nile Delta. Emerg. Infect. Dis. 1996, 2, 234–235. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Mizusawa, K.; Saughust, E.S. A revision of the adult and larval mosquitoes of Japan (including the Ryukyu Archipelago and the Ogasawara Islands) and Korea (Diptera: Culicidae). Contrib. Am. Entomol. Inst. 1979, 16, 148–152. [Google Scholar]
- Bentley, M.D.; Day, J.F. Chemical ecology and behavioral aspects of mosquito oviposition. Annu. Rev. Entomol. 1989, 34, 401–421. [Google Scholar] [CrossRef] [PubMed]
- Beehler, J.W.; Mulla, M.S. Effects of organic enrichment on temporal distribution and abundance of culicine egg rafts. J. Am. Mosquito. Contr. 1995, 11, 167–171. [Google Scholar]
- Reisen, W.K.; Lothrop, H.D.; Hardy, J.L. Bionomics of Culex tarsalis (Diptera: Culicidae) in relation to arbovirus transmission in southeastern California. J. Med. Entomol. 1995, 32, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.C.; Lee, K.W.; Jones, J.W.; Korch, G.W. Seasonal prevalence of mosquitoes collected from light trap in Korea (1993–1994). Korean J. Entomol. 1997, 27, 21–28. [Google Scholar]
- Kim, H.C.; Chong, S.T.; Pike, J.G.; O’guinn, M.L.; Pacha, L.A.; Lee, H.C.S.; Klein, T.A. Seasonal prevalence of mosquitoes collected from light traps in the Republic of Korea, 2002. Korean J. Entomol. 2004, 34, 177–186. [Google Scholar] [CrossRef]
- Zou, L.; Miller, S.N.; Schmidtmann, E.T. Mosquito larval habitat mapping using remote sensing and GIS: Implications of coalbed methane development and West Nile virus. J. Med Entomol. 2006, 43, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- Chaves, L.F.; Hamer, G.L.; Walker, E.D.; Brown, W.M.; Ruiz, M.O.; Kitron, U.D. Climatic variability and landscape heterogeneity impact urban mosquito diversity and vector abundance and infection. Ecosphere 2001, 2. [Google Scholar] [CrossRef]
- Yi, H.; Devkota, B.R.; Yu, J.-S.; Oh, K.-C.; Kim, J.; Kim, H.-J. Effects of global warming on mosquitoes & mosquito-borne diseases and the new strategies for mosquito control. Entomol. Res. 2014, 44, 215–235. [Google Scholar]
- Yi, H.; Yu, J.-S.; Park, K.; Kim, S.; Shin, E.-H.; Lee, W.; Park, M. Development of the smart mosquito counter & its capability: Can we measure the mosquito population? In Proceeding of the Korean Society of Applied Entomology Conference 2013, Seoul, Korea, 24 October 2013. Abstract P163.
- Mielke Jr, P.W.; Berry, K.J.; Johnson, E.S. Multi-response permutation procedures for a priori classifications. Commun. Stat. Theory 1976, 5, 1409–1424. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, Second English edition; Elsevier Science BV: Amsterdam, The Netherlands, 1998. [Google Scholar]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM software design: Gleneden Beach, OR, USA, 2002; Vol. 28. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD: Multivariate Analysis of Ecological Data. Version 2.0, MjM software design: Gleneden Beach, OR, USA, 1999.
- Cristianini, N.; Shawe-Taylor, J. An Introduction to Support Vector Machines and Other Kernel-Based Learning Methods; Cambridge university press: Cambridge, UK, 2000. [Google Scholar]
- Pouteau, R.; Meyer, J.Y.; Taputuarai, R.; Stoll, B. Support vector machines to map rare and endangered native plants in Pacific islands forests. Ecol. Infor. 2012, 9, 37–46. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.; Stone, C.J.; Olshen, R.A. Classification and Regression Trees; CRC press: Boca Raton, FL, USA, 1984. [Google Scholar]
- Hansen, B.; Fotel, F.L.; Jensen, N.J.; Madsen, S.D. Bacteria associated with a marine planktonic copepod in culture. II. Degradation of fecal pellets produced on a diatom, a nanoflagellate or a dinoflagellate diet. J. Plankton Res. 1996, 18, 275–288. [Google Scholar] [CrossRef]
- Vayssières, M.P.; Plant, R.E.; Allen-Diaz, B.H. Classification trees: An alternative non-parametric approach for predicting species distributions. J. Veg. Sci. 2000, 11, 679–694. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Casajus, N.; Lek, S.; Grenouillet, G. Uncertainty in ensemble forecasting of species distribution. Glob. Chan. Biol. 2010, 16, 1145–1157. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C., Jr.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Buisson, L.; Grenouillet, G.; Casajus, N.; Lek, S. Predicting the potential impacts of climate change on stream fish assemblages. Am. Fish Soc. Symp. 2010, 73, 327–346. [Google Scholar]
- Meyer, D.; Dimitriadou, E.; Hornik, K.; Weingessel, A.; Leisch, F.; Chang, C.C.; Lin, C.C. e1071: Misc Functions of the Department of Statistics (e1071), TU Wien. R Package Version 1. 6–3. Available online: http://www.r-project.org (accessed on 12 September 2014).
- Therneau, T.; Atkinson, B.; Ripley, B. rpart: Recursive Partitioning and Regression Trees. R Package Version 4. 1–8. Available online: http://www.r-project.org (accessed on 12 May 2014).
- Robnik-Sikonja, M.; Savicky, P.; Robnik-Sikonja, M.M. Package ‘CORElearn’. Available online: http://lkm.fri.uni-lj.si/rmarko/software/ (accessed on 20 August 2013).
- Robnik-Sikonja, M. Improving random forests. Mach. Learn 2004, 3201, 359–370. [Google Scholar]
- StatSoft. Statistica. Version 6, Statsof: Tulsa, OK, USA, 2004.
- Bi, P.; Hiller, J.; Cameron, A.S.; Zhang, Y.; Givney, R. Climate variability and Ross River virus infections in River land, South Australia, 1992–2004. Epidemiol. Infect. 2009, 137, 1486–1493. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Uchida, K.; Mori, A.; Mine, M.; Eshita, Y.; Kurokawa, K.; Kato, K.; Tahara, H. Effects of high temperature on the emergence and survival of adult Culex pipiens molestus and Culex quinquefasciatus in Japan. J. Am. Mosq. Contr. 1999, 15, 153–156. [Google Scholar]
- Lee, D.K.; Kim, H.C.; Yang, Y.C.; Lee, W.J. Directory for Control and Management Against Vector Mosquitoes; Korea Center for Disease Control and Prevention: Seoul, South Korea, 2005. [Google Scholar]
- Mercer, D.R.; Sheeley, S.L.; Brown, E.J. Mosquito (Diptera: Culicidae) development within microhabitats of an Iowa wetland. J. Med. Entomol. 2005, 42, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, O.; Liu, W.; Miao, J.; Xu, W. Seasonal change in the oviposition activity of Culex pipiens pallens in Shangai, China. Tropic. Med. 1982, 24, 229–234. [Google Scholar]
- Chaves, L.F.; Higa, Y.; Lee, S.H.; Jeong, J.Y.; Heo, S.T.; Kim, M.; Minakawa, N.; Lee, K.H. Environmental forcing shapes regional house mosquito synchrony in a warming temperate island. Environ. Entomol. 2013, 42, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Cianci, D.; Hartemink, N.; Ibáñez-Justicia, A. Modelling the potential spatial distribution of mosquito species using three different techniques. Int. J. Health Geogr. 2015, 14. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; de Baets, B.; Waegeman, W.; Verwaeren, J. Habitat prediction and knowledge extraction for spawning European grayling (Thymallus thymallus L.) using a broad range of species distribution models. Environ. Model Softw. 2013, 47, 1–6. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Bae, M.J.; Hwang, S.J.; Kim, S.H.; Park, Y.S. Predicting potential impacts of climate change on freshwater fish in Korea. Ecol. Infor. 2015, 29, 156–165. [Google Scholar] [CrossRef]
- Lebl, K.; Brugger, K.; Rubel, F. Predicting Culex pipiens/restuans population dynamics by interval lagged weather data. Parasit Vect. 2013, 6. [Google Scholar] [CrossRef] [PubMed]
- Vinogradova, E.B. Culex pipiens pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and Control; Pensoft Publishers: Sofia, Bulgaria, 2000. [Google Scholar]
- Russell, R.C. Ross River virus: Ecology and distribution. Annu. Rev. Entomol. 2002, 47, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.T.; Williams-Newkirk, A.J.; Kitron, U.D.; Chaves, L.F. Seasonal weather, nutrient dynamics and conspecific presence impacts on the southern house mosquito oviposition dynamics in combined sewage overflows. J. Med. Entomol. 2012, 49, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.W.; Hildreth, M.B.; Vanroekel, D.L.; Wimberly, M.C. Weather and land cover influences on mosquito populations in Sioux Falls, South Dakota. J. Med. Entomol. 2011, 48, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ogden, N.H.; Zhu, H. The impact of weather conditions on Culex pipiens and Culex restuans (Diptera: Culicidae) abundance: A case study in peel region. J. Med. Entomol. 2011, 48, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Clements, A.N. The Physiology of Mosquitoes: International Series of Monographs on Pure and Applied Biology; Pergamon Press: Oxford, UK, 1963. [Google Scholar]
- McMichael, A.J.; Haines, A.; Kovats, R.S.; Slooff, R. Climate Changes and Human Health; World Health Organization: Geneva, Switzerland, 1996. [Google Scholar]
- Bi, P.; Zhang, Y.; Parton, K.A. Weather variables and Japanese encephalitis in the metropolitan area of Jinan city, China. J. Infection 2007, 55, 551–556. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, Y.-S.; Bae, M.-J.; Chung, N.; Lee, Y.-R.; Hwang, S.; Kim, S.-A.; Choi, Y.J.; Park, Y.-S. Modeling Occurrence of Urban Mosquitos Based on Land Use Types and Meteorological Factors in Korea. Int. J. Environ. Res. Public Health 2015, 12, 13131-13147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph121013131
Kwon Y-S, Bae M-J, Chung N, Lee Y-R, Hwang S, Kim S-A, Choi YJ, Park Y-S. Modeling Occurrence of Urban Mosquitos Based on Land Use Types and Meteorological Factors in Korea. International Journal of Environmental Research and Public Health. 2015; 12(10):13131-13147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph121013131
Chicago/Turabian StyleKwon, Yong-Su, Mi-Jung Bae, Namil Chung, Yeo-Rang Lee, Suntae Hwang, Sang-Ae Kim, Young Jean Choi, and Young-Seuk Park. 2015. "Modeling Occurrence of Urban Mosquitos Based on Land Use Types and Meteorological Factors in Korea" International Journal of Environmental Research and Public Health 12, no. 10: 13131-13147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph121013131