
DNA Damage, Cell Cycle Arrest, and Apoptosis Induction Caused by Lead in Human Leukemia Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Cell/Tissue Culture
2.3. Biochemical Test for Live Cells and Necrotic Death Cells by Cellometer Imaging
2.4. Biochemical Test for DNA Damage by Comet Assay
2.5. Biochemical Test for Cell Cycle Distribution by Flow Cytometry
2.6. Biochemical Test for Apoptosis by DNA Laddering Assay
2.7. Statistical Analysis
3. Results and Discussion
3.1. Lead Nitrate Inhibited Cell Growth

3.2. Lead Nitrate Induced Necrotic Cell Death

3.3. Lead Nitrate Induced Genotoxic Damage


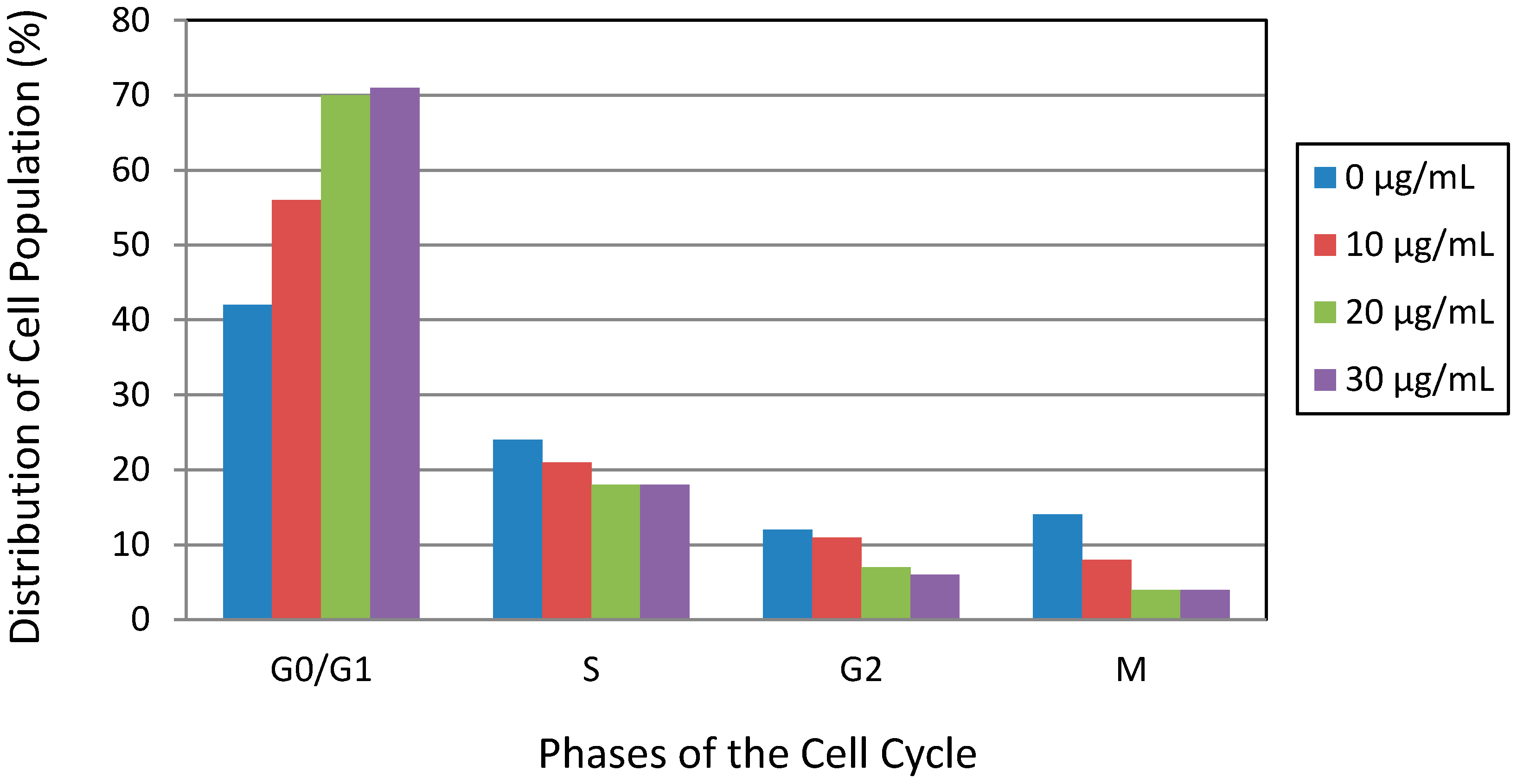
3.4. Lead Nitrate Induced Cell Cycle Arrest


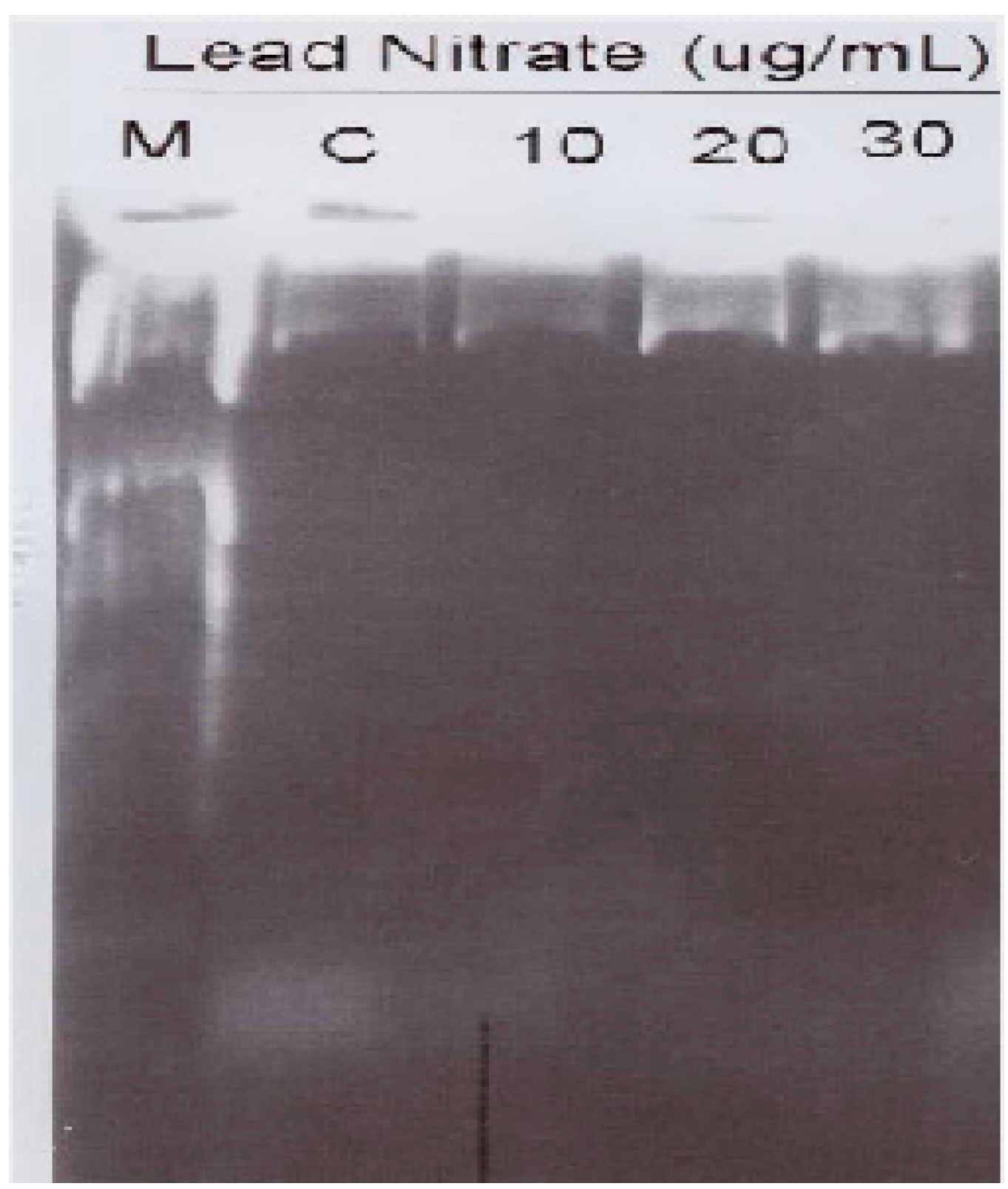
3.5. Lead Nitrate Induced Apoptosis


4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Environmental Protection Agency (EPA). Lead in Drinking Water. 2014. Available online: http://water.epa.gov/drink/info/lead/index.cfm (accessed on 4 January 2014). [Google Scholar]
- Brochin, R.; Leone, S.; Phillips, D.; Shepard, N.; Zisa, D.; Angerio, A. The Cellular Effect of Lead Poisioning and Its Clinical Picture. Available online: https://blogs.commons.georgetown.edu/journal-of-health-sciences/files/Brochin-et-al-2008-GUJHS-5-2-The-Cellular-Effect-of-Lead-Poisoning-and-Its-Clinical-Picture.pdf (accessed on 16 August 2015).
- Ahmed, Y.F.; Eldebaky, H.A.A.; Mahmoud, K.G.M.; Nawito, M. Effects of lead exposure on DNA damage and apoptosis in reproductive and vital organs in female rabbits. Glob. Vet. 2012, 9, 401–408. [Google Scholar]
- Yedjou, C.G.; Haynes, L.; Dorsey, W.; McMurray, R.; Tchounwou, P.B. Lead-induced cytotoxity and oxidative stress in human leukemia (HL-60) cells. Met. Ions Biol. Med. 2008, 10, 489–494. [Google Scholar]
- Xu, Y.; Li, G.; Han, C.; Sun, L.; Zhao, R.; Cui, S. Protective effect of Hippophae rhamnoides L. juice on lead-induced neurotoxicity in mice. Biol. Pharm. Bull. 2005, 28, 490–494. [Google Scholar] [CrossRef] [PubMed]
- El-Nekeety, A.A.; El-Kady, A.A.; Soliman, M.S.; Hassan, N.S.; Abdel-Wahhab, M.A. Protective effect of Aquilegia vulgaris (L.) against lead acetate-inducedoxidative stress in rats. Food Chem. Toxicol. 2009, 47, 2209–2215. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Z.; Jia, X.Y. Low levels of lead exposure induce oxidative damage and DNA damage in the testes of the frog Rana nigromaculata. Ecotoxicology 2009, 18, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Flora, S.J.S.; Pande, M.; Bhadauria, S.; Kannan, G.M. Combined administration of taurine and meso 2,3-dimercaptosuccinic acid in the treatment of chronic lead intoxication in rats. Hum. Exp. Toxicol. 2004, 23, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Farmand, F.; Ehdaie, A.; Roberts, C.K.; Sindhu, R.K. Leadinduced dysregulation of superoxide dismutases, catalase, glutathione peroxidase, and guanylate cyclase. Environ. Res. 2005, 98, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Bressler, J.; Kim, K.A.; Chakraborti, T.; Goldstein, G. Mechanism of lead neurotoxicity. Neurochem. Res. 1999, 24, 595–600. [Google Scholar] [CrossRef] [PubMed]
- ACOG. Committee Opinion Number 533: Lead Screening During Pregnancy and Lactation. 2012. Available online: http://www.acog.org/Resources-And-Publications/Committee-Opinions/Committee-on-Obstetric-Practice/Lead-Screening-During-Pregnancy-and-Lactation (accessed on 16 August 2015).
- Center for Disease Control and Prevention (CDC). Guidelines for the Identification and Treatment of Lead Exposure in Pregnant and Lactating Women. 2010. Available online: http://www.cdc.gov/nceh/lead/publications/leadandpregnancy2010.pdf (accessed on 16 August 2015). [Google Scholar]
- Kwon, S.Y.; Bae, O.N.; Noh, J.Y.; Kim, K.; Kang, S.; Shin, Y.J.; Lim, K.M.; Chung, J.H. Erythrophagocytosis of lead-exposed erythrocytes by renal tubular cells: Possible role in lead-induced nephrotoxicity. Environ. Health Perspect. 2015, 123, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Mishra, K.P.; Singh, V.K.; Rani, R.; Yadav, V.S.; Chandran, V.; Srivastava, S.P.; Seth, P.K. Effect of lead exposure on the immune response of some occupationally exposed individuals. Toxicology 2003, 188, 251–259. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, R.; Qin, R. Effect of sub-acute low level exposure to lead on cellular immune function in rats. Wei Sheng Yan Jiu 2000, 29, 354–356. [Google Scholar] [PubMed]
- Yedjou, C.; Milner, J.; Howard, C.; Tchounwou, P. Basic apoptotic mechanisms of lead toxicity in human leukemia (HL-60) cells. Int. J. Environ. Res. Public Health 2010, 7, 2008–2017. [Google Scholar] [CrossRef] [PubMed]
- Yedjou, C.G.; Saeed, M.A.; Hossain, A.; Dorsey, W.; Yu, H.; Tchounwou, P.B. Basic apoptotic and necrotic cell death in human liver carcinoma (HepG2) cells induced by synthetic azamacrocycle. Environ. Toxicol. 2014, 29, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. Comet Assay for DNA damage and repair: Principles, applications and limitations. Mol. Biotechnol. 2001, 26, 249–261. [Google Scholar] [CrossRef]
- Collins, A.R.; Dusinská, M.; Horská, A. Detection of alkylation damage in human lymphocyte DNA with the comet assay. Acta Biochim. Pol. 2001, 48, 611–614. [Google Scholar] [PubMed]
- Yedjou, C.G.; Tchounwou, P.B. In-vitro genotoxic effect of arsenic trioxide to human leukemia (HL-60) cells using the comet assay. Mol. Cell. Biochem. 2007, 301, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.C. Accelerated apoptotic DNA laddering protocol. BioTechniques 2002, 33, 734–736. [Google Scholar] [PubMed]
- Tchounwou, P.B.; Yedjou, C.G.; Foxx, D.; Ishaque, A.; Shen, E. Lead induced cytotoxicity and transcriptional activation of stress genes in human liver carcinoma cells. Mol. Cell. Biochem. 2004, 255, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Tully, D.B.; Collins, B.J.; Overstreet, J.D.; Smith, C.S.; Dinse, G.E.; Mumtaz, M.M.; Chapin, R.E. Effects of arsenic, cadmium, chromium and lead on gene expression regulated by a battery of 13 different promoters in recombinant HepG2 cells. Toxicol. Appl. Pharmacol. 2000, 168, 79–90. [Google Scholar] [CrossRef] [PubMed]
- White, L.D.; Cory-Slechta, D.A.; Gilbert, M.E.; Tiffany-Castiglioni, E.; Zawia, N.H.; Virgolini, M.; Rossi-George, A.; Lasley, S.M.; Qian, Y.C.; Basha, M.R. New and evolving concepts in the neurotoxicology of lead. Toxicol. Appl. Pharmacol. 2007, 225, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Standard Surveillance Definitions and Classifications. 2012. Available online: www.cdc.gov/nceh/lead/data/definitions.htm (accessed on 6 August 2013). [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Blood lead levels in young children—United States and selected states, 1996–1999. Morb. Mortal. Wkly. Rep. 2000, 49, 1133–1137. [Google Scholar]
- Iavicoli, I.; Marinaccio, A.; Castellino, N.; Carelli, G. Altered cytokine production in mice exposed to lead acetate. Int. J. Immunopathol. Pharmacol. 2004, 17, 97–102. [Google Scholar] [PubMed]
- Iavicoli, I.; Carelli, G.; Stanek, E.J.; Castellino, N.; Calabrese, E.J. Below background levels of blood lead impact cytokine levels in male and female mice. Toxicol. Appl. Pharmacol. 2006, 210, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Wyllie, A.H. Death from inside out: An overview. Philos. Trans. R. Soc. Lond. B: Biol. Sci. 1994, 345, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Paramjit, G.; Saleha, B.B.; Dana, D.K.; Sayeeda, B. In vivo genotoxic effects of mercuric chloride in rat peripheral blood leucocytes using comet assay. Toxicology 2001, 167, 191–197. [Google Scholar]
- Rabbani-Chadegani, A.; Abdosamadi, S.; Fani, N.; Mohammadian, S. A comparison of the effect of lead nitrate on rat liver chromatin, DNA, and histone proteins in solution. Arch. Toxicol. 2010, 83, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Pagliara, P.; Carla, E.C.; Caforio, S.; Chionna, A.; Massa, S.; Abbro, L.; Luciana, D. Kupffer cells promote lead nitrate induced hepatocyte apoptosis via oxidative stress. Comp. Hepatol. 2003, 2, 8–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza, M.E.; Bijur, G.N.; Williams, M.V. Lead and mercury mutagenesis: Role of H2O2, superoxide dismutase, and xanthine oxidase. Environ. Mol. Mutagen. 1998, 31, 352–361. [Google Scholar] [CrossRef]
- Yang, J.L.; Yeh, S.C.; Chang, C.Y. Lead acetate mutagenicity and mutational spectrum in the hypoxanthine guanine phosphoribosyltransferase gene of Chinese hamster ovary K1 cells. Mol. Carcinog. 1996, 17, 181–191. [Google Scholar] [CrossRef]
- Poma, A.; Pittaluga, E.; Tucci, A. Lead acetate genotoxicity on human melanoma cells in vitro. Melanoma Res. 2003, 13, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.; Rossman, T. Mutagenesis and comutagenesis by lead compounds. Mutat. Res. 1992, 298, 97–103. [Google Scholar] [CrossRef]
- Wise, J.P.; Orenstein, J.M.; Patierno, S.R. Inhibition of lead chromate clastogenesis by ascorbate: Relationship to particle dissolution and uptake. Carcinogenesis 1993, 14, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Darzynkiewicz, Z.; Zhao, H.; Halicka, H.D.; Rybak, P.; Dobrucki, J.; Wlodkowic, D. DNA damage signaling assessed in individual cells in relation to the cell cycle phase and induction of apoptosis. Crit. Rev. Clin. Lab. Sci. 2011, 49, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Schnittger, A. Cell cycle and differentiation. Curr. Opin. Plant Biol. 2004, 7, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ohnishi, K.; Shigeno, K.; Fugisawa, S.; Naito, K.; Nakamura, S.; Takeshita, K.; Takeshita, A.; Ohno, R. The induction of apoptosis and cell cycle arrest by arsenic trioxide in lymphoid neoplasms. Leukemia 1998, 12, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.C.; Sun, Y.H.; Chang, K.Z.; Ma, X.F.; Huang, S.L.; Bai, Y.H.; Kang, J.; Liu, Y.G.; Chu, J.J. Selective induction of apoptosis of NB4 cells from G2+M phase by sodium arsenite at lower doses. Eur. J. Haematol. 1998, 61, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ji, L.D.; Xu, L.H. Lead-induced apoptosis in PC-12 cells: Involvement of p53, Bcl-2 family and caspase-3. Toxicol. Lett. 2006, 166, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Naoi, M.; Maruyama, W.; Akao, Y.; Zhang, J.; Parvez, H. Apoptosis induced by an endogenous neurotoxin, N-methyl(R) salsolinol, in dopamine neurons. Toxicology 2000, 153, 123–141. [Google Scholar] [CrossRef]
- Wyllie, A.H.; Kerr, J.F.; Currie, A.R. The significance of apoptosis. Int. Rev. Cytol. 1980, 68, 251–306. [Google Scholar] [PubMed]
- Pulido, M.D.; Parrish, A.R. Metal-induced apoptosis: Mechanisms. Mutat. Res. 2003, 533, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Antonio, A.M.; Druse, M.J. Antioxidants prevent ethanol-associated apoptosis in fetal rhombencephalic neurons. Brain Res. 2008, 1204, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Dai, S.; Yin, Z.; Lu, H.; Jia, R.; Xu, J.; Song, X.; Li, L.; Shu, Y.; Zhao, X.; et al. Sub-chronic lead and cadmium co-induce apoptosis protein expression in liver and kidney of rats. Int. J. Clin. Exp. Pathol. 2014, 7, 2905–2914. [Google Scholar] [PubMed]
- Agarwal, S.; Roy, S.; Ray, A.; Mazumder, S.; Bhattacharya, S. Arsenic trioxide and lead acetate induce apoptosis in adult rat hepatic stem cells. Cell Biol. Toxicol. 2009, 25, 403–413. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yedjou, C.G.; Tchounwou, H.M.; Tchounwou, P.B. DNA Damage, Cell Cycle Arrest, and Apoptosis Induction Caused by Lead in Human Leukemia Cells. Int. J. Environ. Res. Public Health 2016, 13, 56. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph13010056
Yedjou CG, Tchounwou HM, Tchounwou PB. DNA Damage, Cell Cycle Arrest, and Apoptosis Induction Caused by Lead in Human Leukemia Cells. International Journal of Environmental Research and Public Health. 2016; 13(1):56. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph13010056
Chicago/Turabian StyleYedjou, Clement G., Hervey M. Tchounwou, and Paul B. Tchounwou. 2016. "DNA Damage, Cell Cycle Arrest, and Apoptosis Induction Caused by Lead in Human Leukemia Cells" International Journal of Environmental Research and Public Health 13, no. 1: 56. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph13010056