Influence of Heat Events on the Composition of Airborne Bacterial Communities in Urban Ecosystems

Abstract
:1. Introduction
2. Materials and Methods
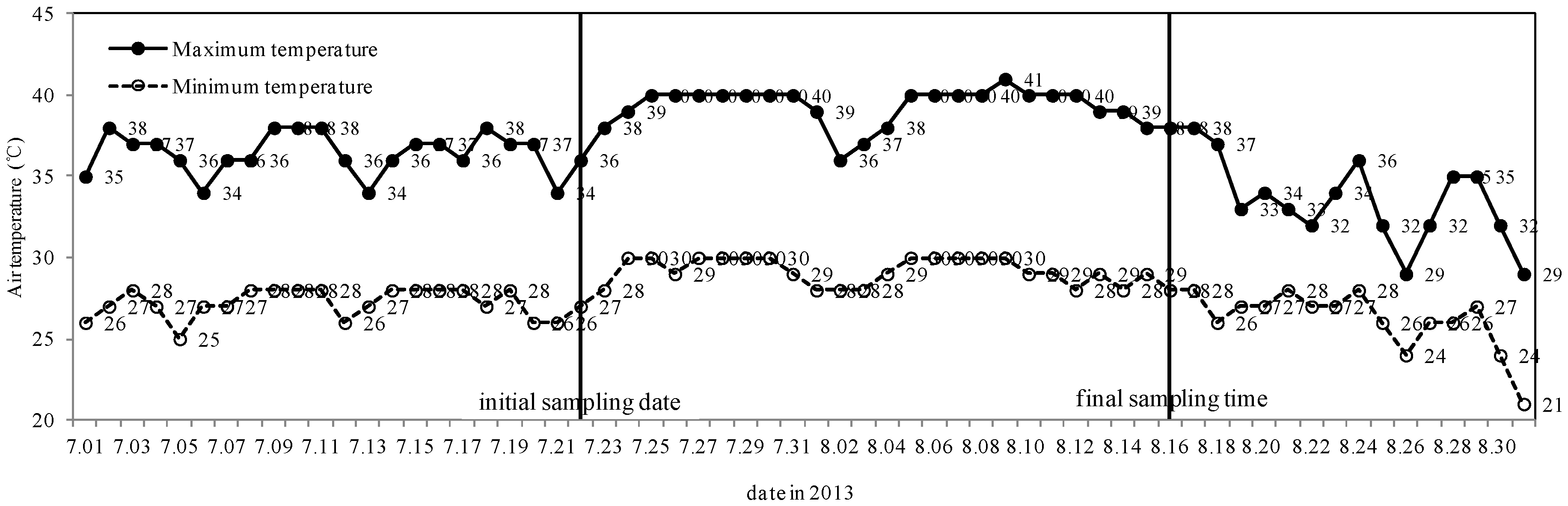
2.1. Selection of Sampling Date
2.2. Description of Sampling Sites
2.3. Sample Collection
2.4. Sample Concentration and DNA Extraction
2.5. Quantitative PCR
2.6. PCR Amplification and Illumina Miseq PE300 Sequencing
2.7. Sequence Analysis
2.8. Statistical Analysis
3. Results
3.1. Weather Conditions during Sampling
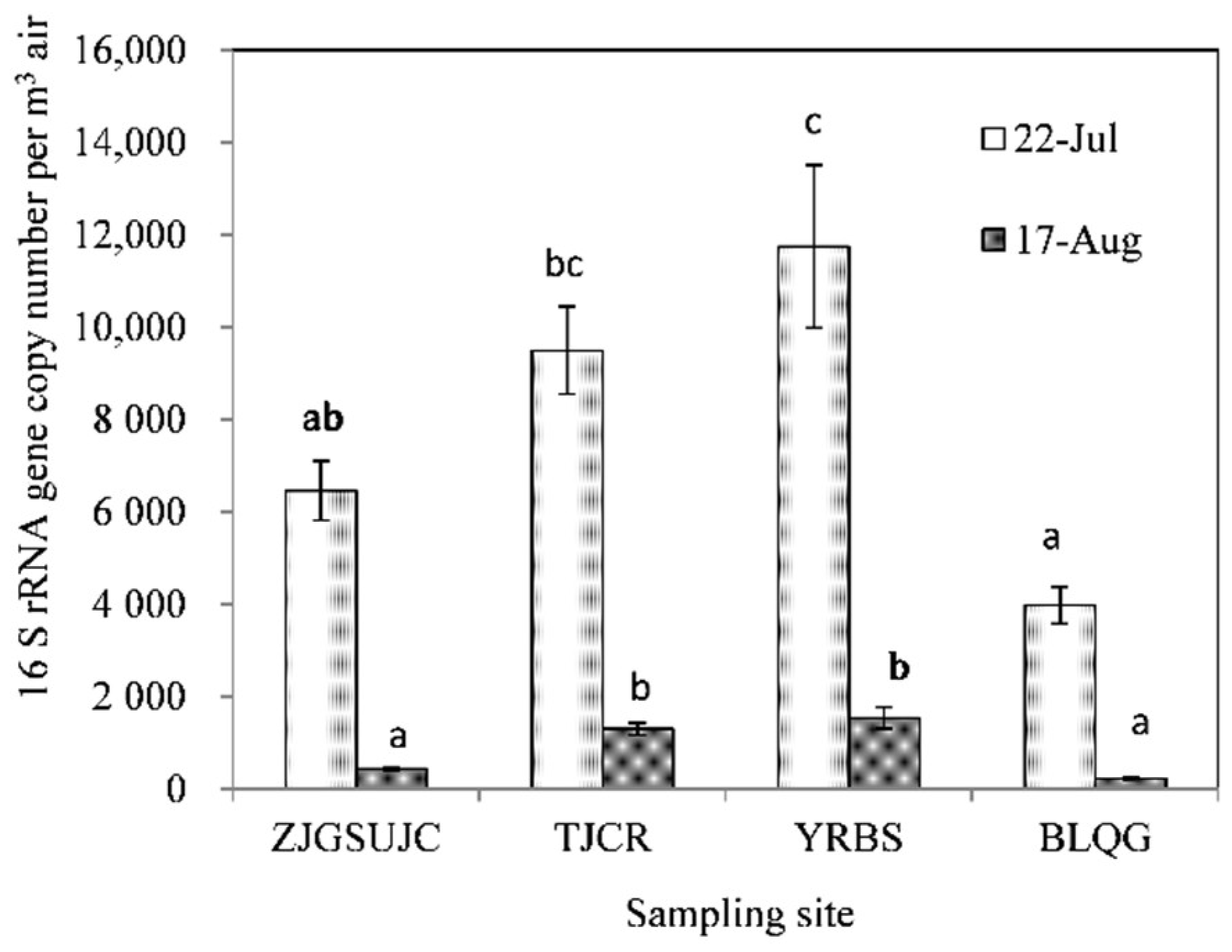
3.2. 16S rRNA Gene Analysis of Airborne Bacteria
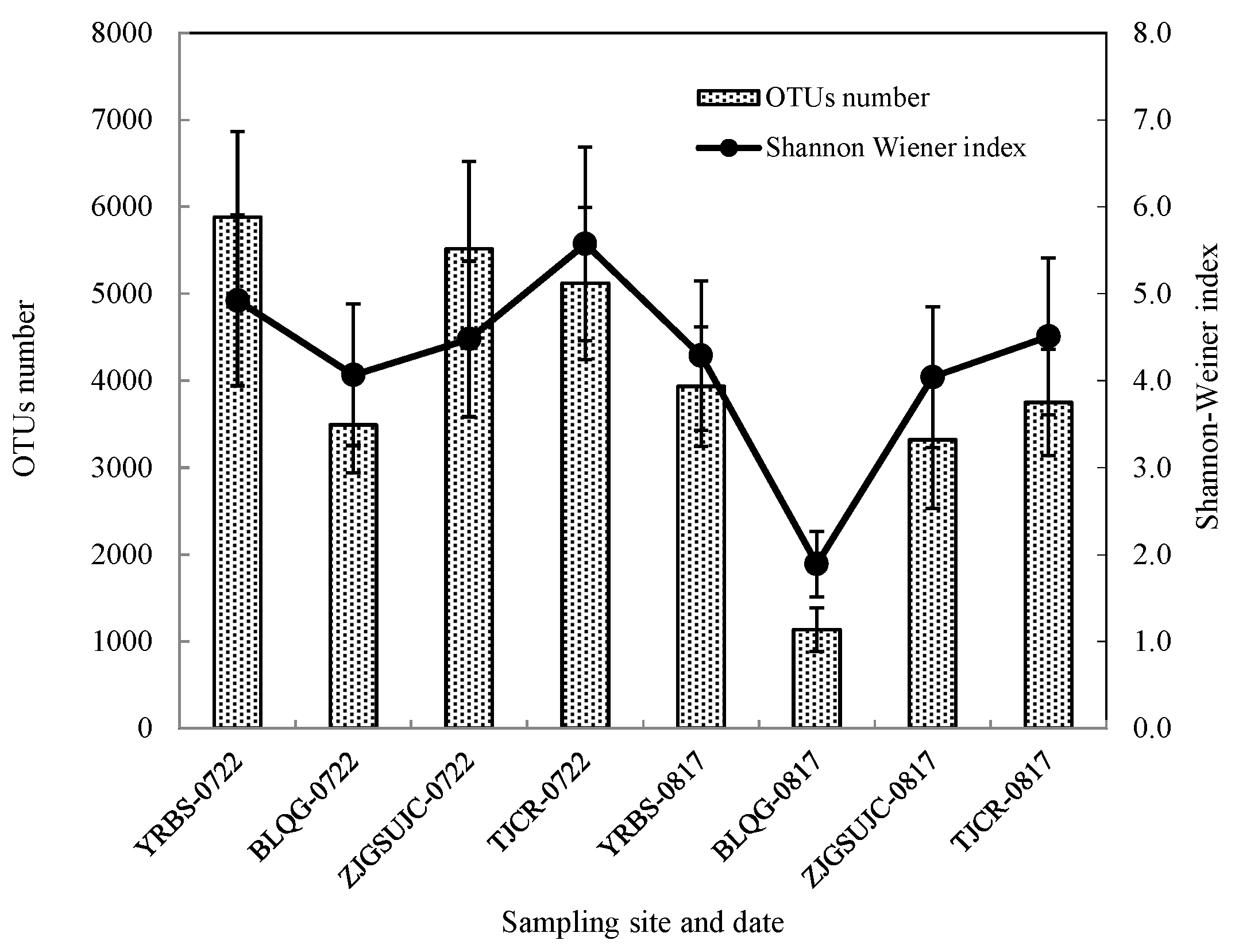
3.3. OTUs (Operational Taxonomic Units) and Shannon–Wiener Diversity Index
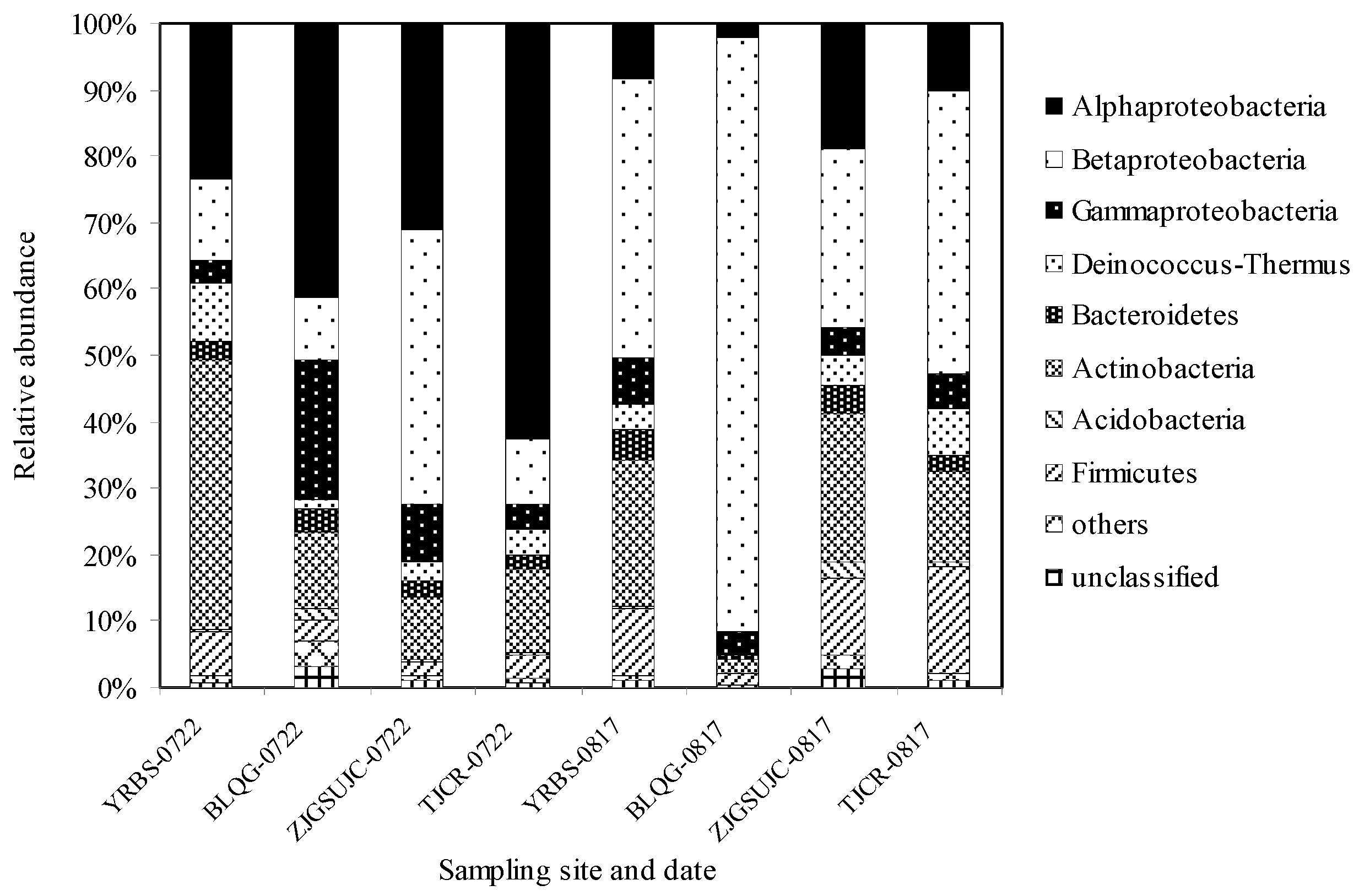
3.4. Abundance of Bacterial Phyla in Airborne Bacterial Communities
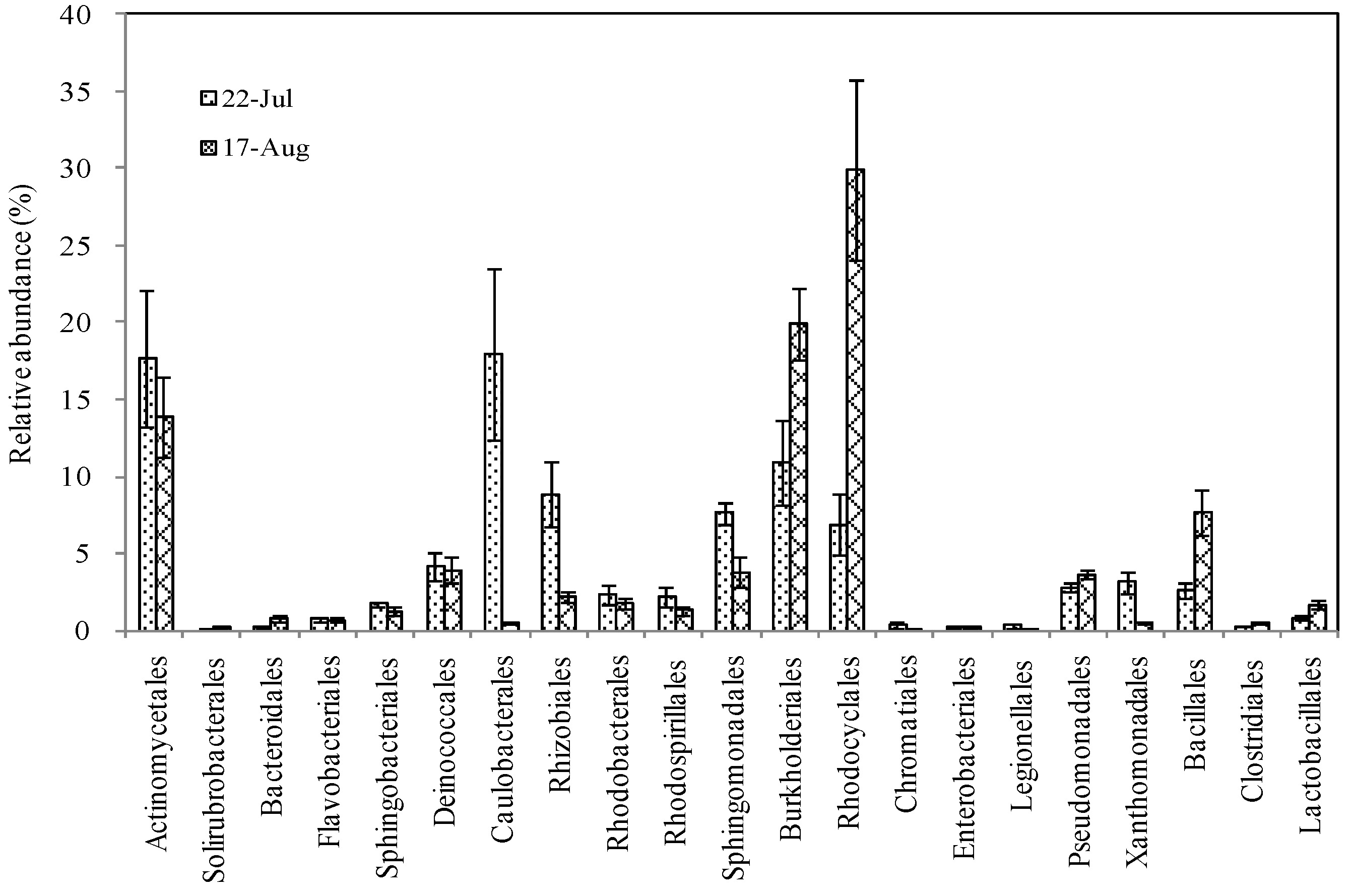
3.5. Dominant Bacterial Orders before and after the Heat Event
3.6. Bacterial Families Differentiate after a Period of Extreme Heat
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jaenicke, R. Abundance of cellular material and proteins in the atmosphere. Science 2005, 308, 73. [Google Scholar] [CrossRef] [PubMed]
- Peccia, J.; Hernandez, M. Incorporating polymerase chain reaction-based identification, population characterization, and quantification of microorganisms into aerosol science: A review. Atmos. Environ. 2006, 40, 3941–3961. [Google Scholar] [CrossRef]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Urbano, R.; Palenik, B.; Gaston, C.J.; Prather, K.A. Detection and phylogenetic analysis of coastal bioaerosols using culture dependent and independent techniques. Biogeosciences 2011, 7, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Dueker, M.E.; O’Mullan, G.D.; Juhl, A.R.; Weathers, K.C.; Uriarte, M. Local environmental pollution strongly influences culturable bacterial aerosols at an urban aquatic superfund site. Environ. Sci. Technol. 2012, 46, 10926–10933. [Google Scholar] [CrossRef] [PubMed]
- Dueker, M.E.; O’Mullan, G.D. Aeration remediation of a polluted waterway increases near-surface coarse and culturable microbial aerosols. Sci. Total Environ. 2014, 478, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Brandi, G.; Sisti, M.; Amagliani, G. Evaluation of the environmental impact of microbial aerosols generated by wastewater treatment plants utilizing different aeration systems. J. Appl. Microbiol. 2010, 88, 845–852. [Google Scholar]
- Carducci, A.; Arrighi, S.; Ruschi, A. Detection of coliphages and enteroviruses in sewage and aerosol from an activated sludge wastewater treatment plant. Lett. Appl. Microbiol. 2010, 21, 207–209. [Google Scholar]
- Albrecht, A.; Witzenberger, R.; Bernzen, U.; Jackel, U. Detection of airborne microbes in a composting facility by cultivation based and cultivation independent methods. Ann. Agric. Environ. Med. 2007, 14, 81–85. [Google Scholar] [PubMed]
- Gangamma, S. Characteristics of airborne bacteria in Mumbai urban environment. Sci. Total Environ. 2014, 488–489, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Smets, W.; Moretti, S.; Denys, S.; Lebeer, S. Airborne bacteria in the atmosphere: Presence, purpose, and potential. Atmos. Environ. 2016, 139, 214–221. [Google Scholar] [CrossRef]
- Martinez, K.; Rao, C.; Burton, N. Exposure assessment and analysis for biological agents. Grana 2004, 43, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Heutte, N.; André, V.; Arvis, C.D.; Bouchart, V.; Lemarié, F.; Legendre, P.; Votier, E.; Louis, M.Y.; Madelaine, S.; Séguin, V.; et al. Assessment of multi-contaminant exposure in a cancer treatment center: A 2-year monitoring of molds, mycotoxins, endotoxins, and glucans in bioaerosols. Environ. Monit. Assess. 2017, 189, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Yao, W.; Lou, X.; Hao, C.; Gong, C.; Ouyang, Z. Profile and characteristics of culturable bacteria in Hangzhou, southeast of China. Aerosol Air Qual. Res. 2016, 16, 1690–1700. [Google Scholar] [CrossRef]
- Fang, Z.G.; Ouyang, Z.Y.; Zheng, H.; Wang, X.K.; Hu, L.F. Culturable airborne bacteria in outdoor environments in Beijing, China. Microb. Ecol. 2007, 54, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.; Hwang, C.Y.; Cho, B.C. Effects of heavy rainfall on the composition of airborne bacterial communities. Front. Environ. Sci. Eng. 2018, 12, 1–12. [Google Scholar] [CrossRef]
- Bertolini, V.; Gandolfi, I.; Ambrosini, R.; Bestetti, G.; Innocente, E.; Rampazzo, G.; Franzetti, A. Temporal variability and effect of environmental variables on airborne bacterial communities in an urban area of Northern Italy. Appl. Microbiol. Biotechnol. 2013, 97, 6561–6570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, R.M.; Clements, N.; Emerson, J.B.; Wiedinmyer, C.; Hannigan, M.P.; Fierer, N. Seasonal variability in bacterial and fungal diversity of the near-surface atmosphere. Environ. Sci. Technol. 2013, 47, 12097–12106. [Google Scholar] [CrossRef] [PubMed]
- Woo, A.C.; Brar, M.S.; Chan, Y.; Lau, M.C.Y.; Leung, F.C.C.; Scott, J.A.; Vrijmoed, L.L.P.; Zawar-Reza, P.; Pointing, S.B. Temporal variation in airborne microbial populations and microbially-derived allergens in a tropical urban landscape. Atmos. Environ. 2013, 74, 291–300. [Google Scholar] [CrossRef]
- Gao, M.; Yan, Y.; Qiu, T.L.; Han, M.L.; Wang, X.M. Variation of correlations between factors and culturable airborne bacteria and fungi. Atmos. Environ. 2016, 128, 10–19. [Google Scholar] [CrossRef]
- Maki, T.; Puspitasari, F.; Hara, K.; Yamada, M.; Kobayashi, F.; Hasegawa, H.; Iwasaka, Y. Variations in the structure of airborne bacterial communities in a down wind area during an Asian dust (Kosa) event. Sci. Total Environ. 2014, 488, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Lighthart, B. Effect of simulated solar radiation on mixed outdoor atmospheric bacterial populations. FEMS Microbiol. Ecol. 1998, 26, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Jeon, E.M.; Kim, H.J.; Jung, K.; Kim, J.H.; Kim, M.Y.; Kim, Y.P.; Ka, J.O. Impact of Asian dust events on airborne bacterial community assessed by molecular analyses. Atmos. Environ. 2011, 45, 4313–4321. [Google Scholar] [CrossRef]
- Cao, C.; Jiang, W.; Wang, B.; Fang, J.; Lang, J.; Tian, G.; Jiang, J.; Zhu, T.F. Inhalable microorganisms in Beijing’s PM2.5 and PM10 pollutants during a severe smog event. Environ. Sci. Technol. 2014, 48, 1499–1507. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Fu, H.L.; Wang, W.; Liu, J.; Meng, Q.L.; Wang, W.K. Characteristics of bacterial and fungal aerosols during the autumn haze days in Xi’an, China. Atmos. Environ. 2015, 122, 439–447. [Google Scholar] [CrossRef]
- Gao, M.; Jia, R.Z.; Qiu, T.L.; Han, M.L.; Song, Y.; Wang, X.M. Seasonal size distribution of airborne culturable bacteria and fungi and preliminary estimation of their deposition in human lungs during non-haze and haze days. Atmos. Environ. 2015, 118, 203–210. [Google Scholar] [CrossRef]
- Dong, L.J.; Qi, J.H.; Shao, C.C.; Zhong, X.; Gao, D.M.; Cao, W.W.; Gao, J.W.; Bai, R.; Long, G.Y.; Chu, C.C. Concentration and size distribution of total airborne microbes in hazy and foggy weather. Sci. Total Environ. 2015, 541, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Baccini, M.; Michelozzi, P. Heat effects on mortality in 15 European cities. Epidemiology 2008, 19, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.D. Lethal effects of heat on bacterial physiology and structure. Sci. Prog. 2003, 86, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Lee, J.E.; Kim, S.S. Thermal effects on bacterial bioaerosols in continuous air flow. Sci. Total Environ. 2009, 407, 4723–4730. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.B.; Li, X.; Wang, T.F.; Wang, B.; Li, C.T.; Zeng, G.M. A review on airborne microorganisms in particulate matters: Composition, characteristics and influence factors. Environ. Int. 2018, 113, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Q.; Deng, Y.; Wang, Y.Q.; Wang, X.K.; Zhang, H.X.; Sun, X.; Ouyang, Z.Y. Meteorological factors had more impact on airborne bacterial communities than air pollutants. Sci. Total Environ. 2017, 601–602, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Brągoszewska, E.; Pastuszka, J.S. Influence of meteorological factors on the level and characteristics of culturable bacteria in the air in Gliwice, Upper Silesia (Poland). Aerobiologia 2018, 34, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaffer, B.T.; Lighthart, B. Survey of culturable airborne bacteria at four diverse locations in Oregon: Urban, rural, forest, and coastal. Microb. Ecol. 1997, 34, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Hoeksma, P.; Aarnink, A.; Ogink, N. Effects of temperature and relative humidity on the survival of airborne bacteria. Livest. Res. Rep. 2015, 864, 1–27. [Google Scholar]
- Mouli, P.C.; Mohan, S.V.; Reddy, S.J. Assessment of microbial (bacteria) concentrations of ambient air at semi-arid urban region: Influence of meteorological factors. Appl. Ecol. Environ. Res. 2005, 3, 139–149. [Google Scholar] [CrossRef]
- Tong, Y.; Lighthart, B. A study of the relationship between pigmented outdoor atmospheric bacteria and solar radiation. Photochem. Photobiol. 1997, 65, 103–106. [Google Scholar] [CrossRef]
- Ray, B. Reversible freeze-injury. In Repairable Lesions in Microorganisms; Hurst, A., Nasim, A., Eds.; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Neidhardt, F.C.; Ingraham, J.L.; Schaechter, M. Physiology of the Bacterial Cell: A Molecular Approach: Sinauer Associates Sunderland; Sinauer Associates Inc.: Sunderland, MA, USA, 1990. [Google Scholar]
- Tringe, S.G.; Zhang, T.; Liu, X.; Yu, Y.; Lee, W.H.; Yap, J.; Yao, F.; Suan, S.T.; Ing, S.K.; Haynes, M. The airborne metagenome in an indoor urban environment. PLoS ONE 2008, 3, e1862. [Google Scholar] [CrossRef] [PubMed]
- Delort, A.M.; Vaïtilingom, M.; Amato, P.; Sancelme, M.; Parazols, M.; Mailhot, G.; Laj, P.; Deguillaume, L. A short overview of the microbial population in clouds: Potential roles in atmospheric chemistry and nucleation processes. Atmos. Environ. 2010, 98, 249–260. [Google Scholar] [CrossRef]
- Kobayashi, F.; Maki, T.; Kakikawa, M.; Yamada, M.; Puspitasari, F.; Iwasaka, Y. Bioprocess of Kosa bioaerosols: Effect of ultraviolet radiation on airborne bacteria within Kosa (Asian dust). J. Biosci. Bioeng. 2015, 119, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Park, J.; Kodama, M.; Ichijo, T.; Baba, T. Changes in the airborne bacterial community in outdoor environments following Asian dust events. Microbes Environ. 2014, 29, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.W. The effects of environmental parameters on the survival of airborne infectious agents. J. R. Soc. Interface 2009, 6, 737–748. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Chan, C.C.; Chew, G.L.; Shih, P.W.; Lee, C.T.; Chao, H.J. Meteorological factors and ambient bacterial levels in a subtropical urban environment. Int. J. Biometeorol. 2012, 56, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, C.; Krempff, A.; Guiraud, H.; Binder, P.; Tiret, C.; Dumenil, G. Atmospheric pollution by airborne microorganisms in the city of Marseilles. Atmos. Environ. 1996, 30, 155–160. [Google Scholar] [CrossRef]
- Macher, J.; Huang, F.; Flores, M. A two-year study of microbiological indoor air quality in a new apartment. Arch. Occup. Environ. Health 1991, 46, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, X.; Zhang, H.; Yao, X.W.; Zhou, M.; Wang, J.Q.; He, Z.F.; Zhang, H.H.; Lou, L.P.; Mao, W.H.; et al. Effect of air pollution on the total bacteria and pathogenic bacteria in different sizes of particulate matter. Environ. Pollut. 2018, 233, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Fedorak, P.M.; Westlake, D.W.S. Effect of sunlight on bacterial survival in transparent air samplers. Can. J. Microbiol. 1978, 24, 618–619. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Zhang, D. Transport of bacterial cells toward the Pacific in northern hemisphere westerly winds. Atmos. Environ. 2014, 87, 138–145. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Functional Type | Architecture Type | Vehicle and Personnel Flow | Vegetation Coverage |
|---|---|---|---|---|
| TJCR | Heavy traffic intersection | High and low office buildings, hotels, and, main traffic road | Approximately 180 vehicles per minute, and 30 persons per minute | Less than 5 percent |
| ZJGSUJC | Cultural and educational area | Experimental buildings, classrooms, student dormitory, and office buildings around | Few vehicles and approximately 10 persons per minute and the number of about 100 persons per minute off class | About 50 percent |
| YRBS | Commercial area and business district | Mall and many shopping buildings around | Approximately 60 vehicles per minute and 80 persons per minute | Less than 5 percent |
| BLQG | Scenic tourist area | No buildings around | Few vehicles and personnel | More than 95 percent |
| Bacterial Groups | YRBS | BLQG | ZJGSUJC | TJCR | ||||
|---|---|---|---|---|---|---|---|---|
| -0722 (%) | -0817 (%) | -0722 (%) | -0817 (%) | -0722 (%) | -0817 (%) | -0722 (%) | -0817 (%) | |
| Acetobacteraceae | 0.90 | 0.80 | 0.62 | 0.10 | 0.77 | 1.79 | 0.76 | 1.29 |
| Acidobacteria | 0.30 | 0.42 | 2.01 | 0.14 | 0.50 | 2.30 | 0.41 | 0.65 |
| Aerococcaceae | 0.19 | 0.15 | 0.08 | 0.01 | 0.01 | 0.10 | 0.24 | 0.26 |
| Alcaligenaceae | 0.02 | 0.03 | 0.81 | 0.00 | 0.04 | 0.01 | 0.04 | 0.06 |
| Alcanivoracaceae | 0.00 | 0.00 | 1.94 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 |
| Alteromonadaceae | 0.02 | 0.03 | 0.07 | 0.01 | 0.00 | 0.01 | 0.02 | 0.05 |
| Bacillaceae | 2.86 | 4.09 | 1.20 | 1.10 | 0.50 | 6.71 | 1.38 | 11.51 |
| Bradyrhizobiaceae | 0.25 | 0.10 | 1.12 | 0.05 | 1.42 | 0.24 | 2.32 | 0.10 |
| Burkholderiaceae | 4.95 | 13.76 | 3.24 | 26.79 | 19.96 | 8.23 | 4.11 | 12.03 |
| Caulobacteraceae | 7.37 | 0.41 | 3.50 | 0.06 | 16.11 | 0.96 | 44.69 | 0.51 |
| Chitinophagaceae | 0.34 | 0.56 | 1.04 | 0.07 | 0.85 | 1.28 | 0.50 | 0.52 |
| Comamonadaceae | 0.83 | 2.87 | 1.19 | 3.38 | 2.88 | 2.21 | 1.21 | 2.40 |
| Corynebacteriaceae | 1.24 | 1.37 | 0.25 | 0.12 | 0.65 | 0.81 | 0.51 | 0.76 |
| Cytophagaceae | 1.31 | 0.41 | 0.33 | 0.09 | 0.55 | 0.82 | 0.48 | 0.36 |
| Deinococcaceae | 8.50 | 4.07 | 1.22 | 0.18 | 2.83 | 4.19 | 3.58 | 7.00 |
| Dermabacteraceae | 1.26 | 0.82 | 0.20 | 0.02 | 0.04 | 0.24 | 0.20 | 0.16 |
| Dietziaceae | 0.30 | 0.20 | 0.18 | 0.04 | 0.03 | 0.13 | 0.08 | 0.34 |
| Enterobacteriaceae | 0.12 | 0.48 | 0.44 | 0.08 | 0.21 | 0.26 | 0.13 | 0.20 |
| Erythrobacteraceae | 0.89 | 0.75 | 0.17 | 0.01 | 0.40 | 0.50 | 0.16 | 0.37 |
| Flavobacteriaceae | 0.65 | 1.11 | 1.04 | 0.12 | 0.40 | 0.88 | 0.64 | 0.48 |
| Geodermatophilaceae | 1.18 | 0.20 | 0.07 | 0.00 | 0.32 | 0.50 | 0.16 | 0.19 |
| Hyphomicrobiaceae | 0.39 | 0.15 | 0.47 | 0.02 | 0.92 | 0.27 | 1.96 | 0.14 |
| Intrasporangiaceae | 1.09 | 0.75 | 0.19 | 0.00 | 0.03 | 0.27 | 0.23 | 0.58 |
| Kineosporiaceae | 0.17 | 0.17 | 0.09 | 0.00 | 0.20 | 0.75 | 0.18 | 0.15 |
| Lactobacillaceae | 0.54 | 0.85 | 0.15 | 0.08 | 0.15 | 0.38 | 0.29 | 0.91 |
| Legionellaceae | 0.00 | 0.03 | 1.17 | 0.00 | 0.01 | 0.01 | 0.02 | 0.02 |
| Methylobacteriaceae | 1.92 | 0.92 | 0.91 | 0.87 | 1.24 | 1.53 | 0.84 | 1.04 |
| Microbacteriaceae | 3.17 | 1.26 | 1.09 | 0.15 | 1.10 | 2.34 | 2.19 | 1.03 |
| Micrococcaceae | 9.76 | 2.79 | 0.71 | 0.11 | 0.29 | 1.09 | 0.53 | 0.99 |
| Moraxellaceae | 1.75 | 3.61 | 0.84 | 1.68 | 2.07 | 1.57 | 1.18 | 2.51 |
| Nocardiaceae | 0.10 | 0.09 | 1.09 | 0.02 | 0.13 | 0.07 | 0.21 | 0.20 |
| Nocardioidaceae | 1.36 | 0.89 | 1.65 | 0.13 | 1.32 | 2.84 | 1.97 | 0.89 |
| Oxalobacteraceae | 0.76 | 0.92 | 0.48 | 0.11 | 0.75 | 1.04 | 0.42 | 1.31 |
| Propionibacteriaceae | 1.99 | 1.25 | 0.51 | 0.23 | 0.24 | 1.42 | 0.52 | 1.15 |
| Pseudomonadaceae | 0.77 | 1.46 | 2.81 | 1.13 | 1.09 | 1.28 | 0.49 | 1.29 |
| Rhodobacteraceae | 5.41 | 2.33 | 1.03 | 0.11 | 1.29 | 2.68 | 1.69 | 1.97 |
| Rhodocyclaceae | 5.16 | 23.40 | 2.38 | 58.34 | 16.49 | 13.30 | 3.45 | 24.62 |
| Rhodospirillaceae | 0.32 | 0.23 | 4.61 | 0.02 | 0.37 | 0.56 | 0.27 | 0.25 |
| Sphingomonadaceae | 4.93 | 1.80 | 10.90 | 0.64 | 6.19 | 7.53 | 6.71 | 3.36 |
| Staphylococcaceae | 1.36 | 2.13 | 0.28 | 0.16 | 0.36 | 0.91 | 0.57 | 0.69 |
| Streptococcaceae | 0.47 | 0.73 | 0.16 | 0.03 | 0.12 | 0.27 | 0.14 | 0.33 |
| Streptomycetaceae | 0.77 | 0.08 | 0.05 | 0.02 | 0.04 | 0.62 | 0.24 | 0.17 |
| Xanthomonadaceae | 0.49 | 0.62 | 5.43 | 0.24 | 4.47 | 0.48 | 1.72 | 0.58 |
| Others | 23.85 | 20.94 | 42.28 | 3.53 | 12.65 | 26.59 | 12.55 | 16.60 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Z.; Guo, W.; Zhang, J.; Lou, X. Influence of Heat Events on the Composition of Airborne Bacterial Communities in Urban Ecosystems. Int. J. Environ. Res. Public Health 2018, 15, 2295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15102295
Fang Z, Guo W, Zhang J, Lou X. Influence of Heat Events on the Composition of Airborne Bacterial Communities in Urban Ecosystems. International Journal of Environmental Research and Public Health. 2018; 15(10):2295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15102295
Chicago/Turabian StyleFang, Zhiguo, Weijun Guo, Junwen Zhang, and Xiuqin Lou. 2018. "Influence of Heat Events on the Composition of Airborne Bacterial Communities in Urban Ecosystems" International Journal of Environmental Research and Public Health 15, no. 10: 2295. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15102295