Metabolomics Reveals Metabolic Changes Caused by Low-Dose 4-Tert-Octylphenol in Mice Liver

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals and Treatments
2.3. Analysis of Serum Levels of ALT and AST
2.4. Histological Evaluations
2.5. Sample Preparation for the Mass Spectrometry Analysis of Mouse Liver and Blood Serum
2.6. Metabolomic Profiling
2.7. Targeted S-Adenosylmethionine Analysis in Liver and 4-Tert-Octylphenol in Blood Serum
2.8. Real-Time PCR Analysis
2.9. Enzyme-Linked Immunosorbent Assay
2.10. Statistical Analysis
3. Results
3.1. Changes of Body Weight and Liver Organ Coefficient
3.2. Histological Results
3.3. Results of the Serum ALT and AST
3.4. Metabolomic Profiles
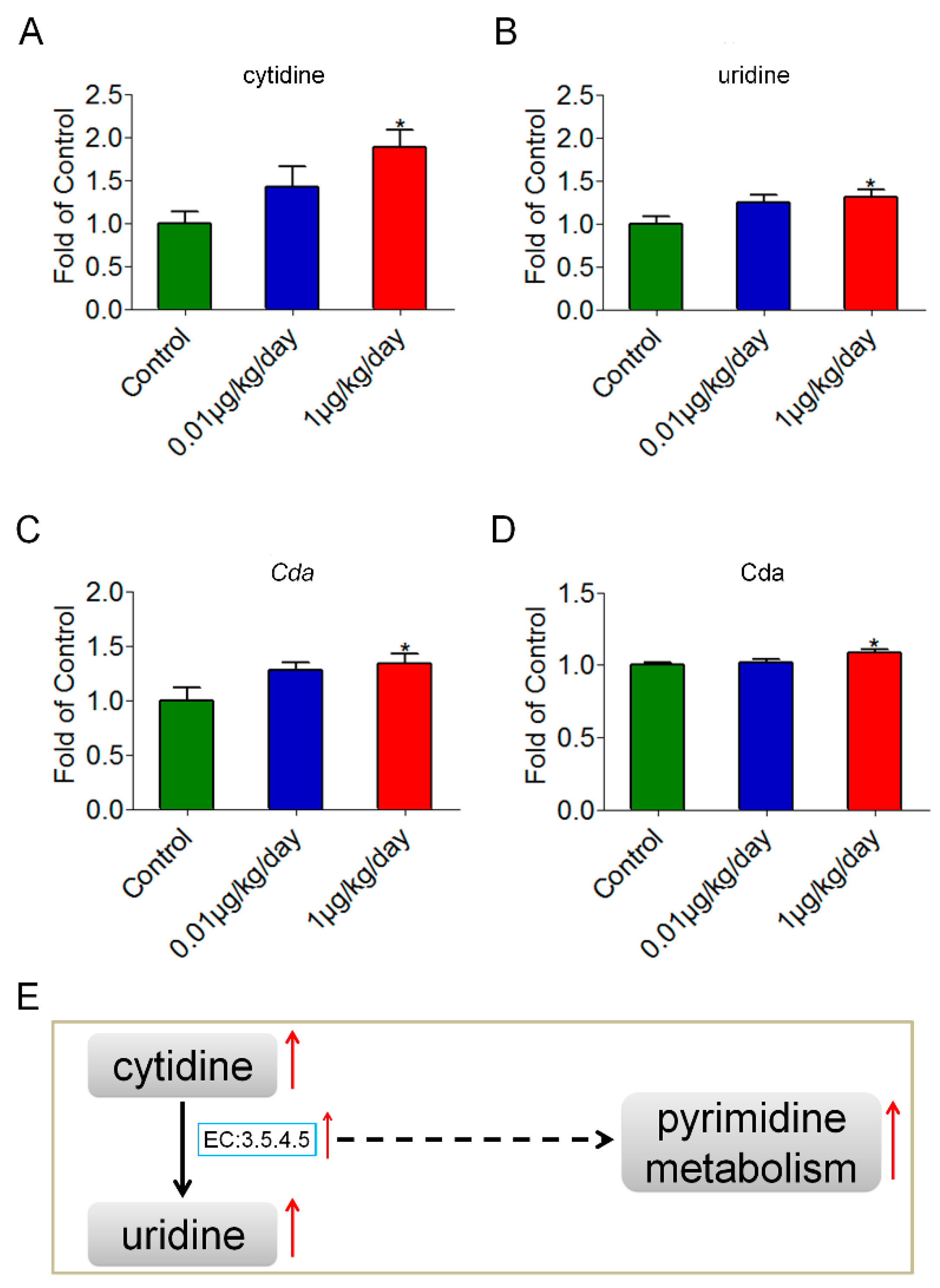
3.5. Low-Dose OP Promoted Pyrimidine Synthesis through Cytidine Deaminase
3.6. Purine Synthesis Was Increased through Pyridoxal-5′-Phosphate (PLP)-Dependent Enzyme-Shmt1 after Low-Dose OP Treatment
3.7. Amino Acid Derivative Was Increased after Low-Dose OP Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nimrod, A.C.; Benson, W.H. Environmental estrogenic effects of alkylphenol ethoxylates. Crit. Rev. Toxicol. 1996, 26, 335–364. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M.; Falkowska, L.; Grabowski, P.; Kwasniak, J.; Mudrak-Cegiolka, S.; Reindl, A.R.; Sokolowski, A.; Szumilo, E.; Zgrundo, A. Bisphenol a, 4-tert-octylphenol, and 4-nonylphenol in the gulf of gdansk (southern baltic). Arch. Environ. Contam. Toxicol. 2014, 67, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.K.; Ding, W.H. Determination of alkylphenolic residues in fresh fruits and vegetables by extractive steam distillation and gas chromatography-mass spectrometry. J. Chromatogr. A 2005, 1088, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Barber, L.B.; Loyo-Rosales, J.E.; Rice, C.P.; Minarik, T.A.; Oskouie, A.K. Endocrine disrupting alkylphenolic chemicals and other contaminants in wastewater treatment plant effluents, urban streams, and fish in the great lakes and upper mississippi river regions. Sci. Total Environ. 2015, 517, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Calafat, A.M.; Ye, X.; Wong, L.Y.; Reidy, J.A.; Needham, L.L. Exposure of the U.S. Population to bisphenol a and 4-tertiary-octylphenol: 2003–2004. Environ. Health Perspect. 2008, 116, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhu, P.; Xu, B.; Zhao, R.; Qiao, S.; Chen, X.; Tang, R.; Wu, D.; Song, L.; Wang, S.; et al. Determination of nine environmental phenols in urine by ultra-high-performance liquid chromatography-tandem mass spectrometry. J. Anal. Toxicol. 2012, 36, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.N.; Christiansen, L.B.; Pedersen, K.L.; Korsgaard, B.; Bjerregaard, P. In vivo estrogenic activity of branched and linear alkylphenols in rainbow trout (oncorhynchus mykiss). Sci. Total Environ. 1999, 233, 89–96. [Google Scholar] [CrossRef]
- Chen, M.; Tang, R.; Fu, G.; Xu, B.; Zhu, P.; Qiao, S.; Chen, X.; Xu, B.; Qin, Y.; Lu, C.; et al. Association of exposure to phenols and idiopathic male infertility. J. Hazard. Mater. 2013, 250–251, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Chen, M.; Wu, W.; Xu, B.; Tang, R.; Chen, X.; Du, G.; Lu, C.; Meeker, J.D.; Zhou, Z.; et al. Interactions between urinary 4-tert-octylphenol levels and metabolism enzyme gene variants on idiopathic male infertility. PLoS ONE 2013, 8, e59398. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Wu, C.; Lu, D.; Qi, X.; Xu, H.; Guo, J.; Liang, W.; Chang, X.; Wang, G.; Zhou, Z. Birth outcome measures and prenatal exposure to 4-tert-octylphenol. Environ. Pollut. 2016, 212, 65–70. [Google Scholar] [CrossRef] [PubMed]
- White, R.; Jobling, S.; Hoare, S.A.; Sumpter, J.P.; Parker, M.G. Environmentally persistent alkylphenolic compounds are estrogenic. Endocrinology 1994, 135, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Madsen, L.L.; Korsgaard, B.; Bjerregaard, P. 4-tert-octylphenol and 17βestradiol applied by feeding to flounder platichthys flesus: Induction of vitellogenin and accumulation in tissues. Mar. Environ. Res. 2002, 54, 729–733. [Google Scholar] [CrossRef]
- Barlas, N.; Aydogan, M. Histopathologic effects of maternal 4-tert-octylphenol exposure on liver, kidney and spleen of rats at adulthood. Arch. Toxicol. 2009, 83, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, E.J.; Park, M.N.; Kim, J.E.; Kim, S.C.; Jeung, E.B.; Lee, G.S.; Hwang, D.Y.; An, B.S. The adverse effect of 4-tert-octylphenol on fat metabolism in pregnant rats via regulation of lipogenic proteins. Environ. Toxicol. Pharmacol. 2015, 40, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Saggu, S.; Sakeran, M.I.; Zidan, N.; Tousson, E.; Mohan, A.; Rehman, H. Ameliorating effect of chicory (chichorium intybus L.) fruit extract against 4-tert-octylphenol induced liver injury and oxidative stress in male rats. Food Chem. Toxicol. 2014, 72, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Xiao, N.; Zhang, Y.H. Toxic effects of octylphenol on the expression of genes in liver identified by suppression subtractive hybridization of rana chensinensis. Ecotoxicology 2014, 23, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Wang, X. Power of metabolomics in biomarker discovery and mining mechanisms of obesity. Obes. Rev. 2013, 14, 344–349. [Google Scholar] [CrossRef] [PubMed]
- De Preter, V. Metabonomics and systems biology. Methods Mol. Boil. 2015, 1277, 245–255. [Google Scholar]
- Ratnasekhar, C.; Sonane, M.; Satish, A.; Mudiam, M.K. Metabolomics reveals the perturbations in the metabolome of caenorhabditis elegans exposed to titanium dioxide nanoparticles. Nanotoxicology 2015, 9, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Mu, X.; Zhang, J.; Huang, Q.; Alamdar, A.; Tian, M.; Liu, L.; Shen, H. Serum metabolomics reveals that arsenic exposure disrupted lipid and amino acid metabolism in rats: A step forward in understanding chronic arsenic toxicity. Metallomics 2015, 7, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Manna, S.K.; Thompson, M.D.; Gonzalez, F.J. Application of mass spectrometry-based metabolomics in identification of early noninvasive biomarkers of alcohol-induced liver disease using mouse model. Adv. Exp. Med. Boil. 2015, 815, 217–238. [Google Scholar]
- Yu, J.; Jiang, Y.S.; Jiang, Y.; Peng, Y.F.; Sun, Z.; Dai, X.N.; Cao, Q.T.; Sun, Y.M.; Han, J.C.; Gao, Y.J. Targeted metabolomic study indicating glycyrrhizin’s protection against acetaminophen-induced liver damage through reversing fatty acid metabolism. Phytother. Res. 2014, 28, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Della Torre, S.; Ciana, P. Selective estrogen receptor modulators and the tissue-selective estrogen complex: Analysis of cell type-specific effects using in vivo imaging of a reporter mouse model. Methods Mol. Boil. 2016, 1366, 297–313. [Google Scholar]
- Horn, T.L.; Bhattacharjee, A.; Schook, L.B.; Rutherford, M.S. Altered hepatic mrna expression of apoptotic genes during dimethylnitrosamine exposure. Toxicol. Sci. 2000, 57, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Duysen, E.G.; Li, B.; Carlson, M.; Li, Y.F.; Wieseler, S.; Hinrichs, S.H.; Lockridge, O. Increased hepatotoxicity and cardiac fibrosis in cocaine-treated butyrylcholinesterase knockout mice. Basic Clin. Pharmacol. Toxicol. 2008, 103, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Kitanaka, J.; Kitanaka, N.; Takemura, M. Chronic methamphetamine administration reduces histamine-stimulated phosphoinositide hydrolysis in mouse frontal cortex. Biochem. Biophys. Res. Commun. 2003, 300, 932–937. [Google Scholar] [CrossRef]
- Sasaki, A.; Takasaki, I.; Andoh, T.; Nojima, H.; Shiraki, K.; Kuraishi, Y. Roles of alpha-adrenoceptors and sympathetic nerve in acute herpetic pain induced by herpes simplex virus inoculation in mice. J. Pharmacol. Sci. 2003, 92, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Urfi, M.K.; Mujahid, M.; Rahman, M.A.; Rahman, M.A. The role of Tamarix gallica leaves extract in liver injury induced by rifampicin plus isoniazid in sprague dawley rats. J. Diet. Suppl. 2018, 15, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Dong, T.; Wang, L.; Guan, Q.; Song, L.; Chen, D.; Zhou, Z.; Chen, M.; Xia, Y.; Wang, X. Obesity aggravates toxic effect of bpa on spermatogenesis. Environ. Int. 2017, 105, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.L.; Ferreira, M.; Silva, J.; Gomes, M.; Dias, F.; Santos, J.I.; Mauricio, J.; Lobo, F.; Medeiros, R. Higher circulating expression levels of miR-221 associated with poor overall survival in renal cell carcinoma patients. Tumour Boil. 2014, 35, 4057–4066. [Google Scholar] [CrossRef] [PubMed]
- Shyh-Chang, N.; Locasale, J.W.; Lyssiotis, C.A.; Zheng, Y.; Teo, R.Y.; Ratanasirintrawoot, S.; Zhang, J.; Onder, T.; Unternaehrer, J.J.; Zhu, H.; et al. Influence of threonine metabolism on S-adenosylmethionine and histone methylation. Science 2013, 339, 222–226. [Google Scholar] [CrossRef] [PubMed]
- da Silva, V.R.; Ralat, M.A.; Quinlivan, E.P.; DeRatt, B.N.; Garrett, T.J.; Chi, Y.Y.; Frederik Nijhout, H.; Reed, M.C.; Gregory, J.F., 3rd. Targeted metabolomics and mathematical modeling demonstrate that vitamin B-6 restriction alters one-carbon metabolism in cultured HepG2 cells. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E93–E101. [Google Scholar] [CrossRef] [PubMed]
- Giardina, G.; Brunotti, P.; Fiascarelli, A.; Cicalini, A.; Costa, M.G.; Buckle, A.M.; di Salvo, M.L.; Giorgi, A.; Marani, M.; Paone, A.; et al. How pyridoxal 5′-phosphate differentially regulates human cytosolic and mitochondrial serine hydroxymethyltransferase oligomeric state. FEBS J. 2015, 282, 1225–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amornwatcharapong, W.; Maenpuen, S.; Chitnumsub, P.; Leartsakulpanich, U.; Chaiyen, P. Human and plasmodium serine hydroxymethyltransferases differ in rate-limiting steps and ph-dependent substrate inhibition behavior. Arch. Biochem. Biophys. 2017, 630, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Ducker, G.S.; Ghergurovich, J.M.; Mainolfi, N.; Suri, V.; Jeong, S.K.; Hsin-Jung Li, S.; Friedman, A.; Manfredi, M.G.; Gitai, Z.; Kim, H.; et al. Human SHMT inhibitors reveal defective glycine import as a targetable metabolic vulnerability of diffuse large B-cell lymphoma. Proc. Natl. Acad. Sci. USA 2017, 114, 11404–11409. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Giovannucci, E.; Wolk, A. Vitamin B6 intake, alcohol consumption, and colorectal cancer: A longitudinal population-based cohort of women. Gastroenterology 2005, 128, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Mooney, S.; Leuendorf, J.E.; Hendrickson, C.; Hellmann, H. Vitamin B6: A long known compound of surprising complexity. Molecules 2009, 14, 329–351. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Garg, P.; Lee, S.; Choung, H.W.; Choung, Y.H.; Choung, P.H.; Chung, J.H. Nucleotide biosynthesis arrest by silencing SHMT1 function via vitamin B6-coupled vector and effects on tumor growth inhibition. Biomaterials 2014, 35, 9332–9342. [Google Scholar] [CrossRef] [PubMed]
- Niculescu, M.D.; Zeisel, S.H. Diet, methyl donors and DNA methylation: Interactions between dietary folate, methionine and choline. J. Nutr. 2002, 132, 2333S–2335S. [Google Scholar] [CrossRef] [PubMed]
- Medici, V.; Schroeder, D.I.; Woods, R.; LaSalle, J.M.; Geng, Y.; Shibata, N.M.; Peerson, J.; Hodzic, E.; Dayal, S.; Tsukamoto, H.; et al. Methylation and gene expression responses to ethanol feeding and betaine supplementation in the cystathionine beta synthase-deficient mouse. Alcohol. Clin. Exp. Res. 2014, 38, 1540–1549. [Google Scholar] [CrossRef] [PubMed]
- Racine, S.X.; Le Toumelin, P.; Adnet, F.; Cohen, Y.; Cupa, M.; Hantz, E.; Le Moyec, L. N-acetyl functions and acetate detected by nuclear magnetic resonance spectroscopy of urine to detect renal dysfunction following aminoglycoside and/or glycopeptide antibiotic therapy. Nephron Physiol. 2004, 97, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, E.J.; Park, M.N.; Lee, J.E.; Hong, S.H.; An, S.M.; Kim, S.C.; Hwang, D.Y.; An, B.S. Adverse effects of 4-tert-octylphenol on the production of oxytocin and hcg in pregnant rats. Lab. Anim. Res. 2014, 30, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.R.; Hwang, K.A.; Nam, K.H.; Kim, H.C.; Choi, K.C. Progression of breast cancer cells was enhanced by endocrine-disrupting chemicals, triclosan and octylphenol, via an estrogen receptor-dependent signaling pathway in cellular and mouse xenograft models. Chem. Res. Toxicol. 2014, 27, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Ahbab, M.A.; Kolankaya, D.; Barlas, N. Influence of vitamin C on bisphenol A, nonylphenol and octylphenol induced oxidative damages in liver of male rats. Food Chem. Toxicol. 2010, 48, 2865–2871. [Google Scholar] [CrossRef] [PubMed]
- Vineis, P.; Bartsch, H.; Caporaso, N.; Harrington, A.M.; Kadlubar, F.F.; Landi, M.T.; Malaveille, C.; Shields, P.G.; Skipper, P.; Talaska, G.; et al. Genetically based n-acetyltransferase metabolic polymorphism and low-level environmental exposure to carcinogens. Nature 1994, 369, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Vineis, P.; Khan, A.E.; Vlaanderen, J.; Vermeulen, R. The impact of new research technologies on our understanding of environmental causes of disease: The concept of clinical vulnerability. Environ. Health 2009, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Garavito, M.F.; Narvaez-Ortiz, H.Y.; Zimmermann, B.H. Pyrimidine metabolism: Dynamic and versatile pathways in pathogens and cellular development. J. Genet. Genom. 2015, 42, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, R. Prebiotic cytosine synthesis: A critical analysis and implications for the origin of life. Proc. Natl. Acad. Sci. USA 1999, 96, 4396–4401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darzynkiewicz, Z.; Traganos, F.; Zhao, H.; Halicka, H.D.; Li, J. Cytometry of DNA replication and rna synthesis: Historical perspective and recent advances based on “click chemistry”. Cytom. Part A 2011, 79, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Fromm, H.; Hargrove, M. Coenzymes and Vitamins. In Essentials of Biochemistry; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Noh, S.; Kim, D.H.; Jung, W.H.; Koo, J.S. Expression levels of serine/glycine metabolism-related proteins in triple negative breast cancer tissues. Tumour Boil. 2014, 35, 4457–4468. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.R.; Herbig, A.K.; Stover, P.J. New perspectives on folate catabolism. Annu. Rev. Nutr. 2001, 21, 255–282. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Si, Z.H.; Li, M.H.; Zhao, H.; Fu, Y.H.; Xing, Y.X.; Hong, W.; Ruan, L.Y.; Li, P.M.; Wang, J.S. 1H NMR-based metabolomics study of liver damage induced by ginkgolic acid (15:1) in mice. J. Pharm. Biomed. Anal. 2017, 136, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, X.; Du, W.; Feng, Y.; Kong, X.; Li, Y.; Xiao, L.; Zhang, P. Antitumor effects of ginkgolic acid in human cancer cell occur via cell cycle arrest and decrease the Bcl-2/Bax ratio to induce apoptosis. Chemotherapy 2010, 56, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Sarma, S.N.; Saleem, A.; Lee, J.Y.; Tokumoto, M.; Hwang, G.W.; Man Chan, H.; Satoh, M. Effects of long-term cadmium exposure on urinary metabolite profiles in mice. J. Toxicol. Sci. 2018, 43, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Wu, X.; Ke, C.; Yin, M.; Li, Z.; Fan, L.; Zhang, W.; Zhang, H.; Zhao, F.; Zhou, X.; et al. Identification of potential biomarkers for ovarian cancer by urinary metabolomic profiling. J. Proteome Res. 2013, 12, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Ng, D.P.; Salim, A.; Liu, Y.; Zou, L.; Xu, F.G.; Huang, S.; Leong, H.; Ong, C.N. A metabolomic study of low estimated GFR in non-proteinuric type 2 diabetes mellitus. Diabetologia 2012, 55, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Bradford, B.U.; O’Connell, T.M.; Han, J.; Kosyk, O.; Shymonyak, S.; Ross, P.K.; Winnike, J.; Kono, H.; Rusyn, I. Metabolomic profiling of a modified alcohol liquid diet model for liver injury in the mouse uncovers new markers of disease. Toxicol. Appl. Pharmacol. 2008, 232, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, N.; Wei, S.; Li, M.; Yang, J.; Li, K.; Jin, L.; Xie, Y.; Giesy, J.P.; Zhang, X.; Yu, H. Effects of perfluorooctanoic acid on metabolic profiles in brain and liver of mouse revealed by a high-throughput targeted metabolomics approach. Sci. Rep. 2016, 6, 23963. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, M.; Li, H.; Zhao, H.; Wang, F.; He, Q.; Zhang, T.; Wang, S. Serum metabonomics study of the hepatoprotective effect of amarogentin on CCL4-induced liver fibrosis in mice by GC-TOF-MS analysis. J. Pharm. Biomed. Anal. 2018, 149, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Casati, L.; Sendra, R.; Colciago, A.; Negri-Cesi, P.; Berdasco, M.; Esteller, M.; Celotti, F. Polychlorinated biphenyls affect histone modification pattern in early development of rats: A role for androgen receptor-dependent modulation? Epigenomics 2012, 4, 101–112. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | GenBank Accession no. | Product Length (bp) | Primer Sequences |
|---|---|---|---|
| Cda | NM_028176.1 | 78 | Sense: 5′-ATGAGAGAGTTTGGCACCGAC-3′ |
| Anti-sense:5′-CTCCTGGACCGTCCTGACTA-3′ | |||
| Shmt1 | NM_009171.2 | 94 | Sense: 5′-CCCGAAACCAAGTGAACTGGA-3′ |
| Anti-sense:5′-ACTGGTTCAGAGTTGCCTTGTA-3′ | |||
| Gapdh | NM_001289726.1 | 124 | Sense: 5′-CCCTTAAGAGGGATGCTGCC-3′ |
| Anti-sense:5′-TACGGCCAAATCCGTTCACA-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, K.; Ding, X.; Yang, J.; Hu, Y.; Song, Y.; Chen, M.; Sun, R.; Dong, T.; Xu, B.; Han, X.; et al. Metabolomics Reveals Metabolic Changes Caused by Low-Dose 4-Tert-Octylphenol in Mice Liver. Int. J. Environ. Res. Public Health 2018, 15, 2686. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122686
Zhou K, Ding X, Yang J, Hu Y, Song Y, Chen M, Sun R, Dong T, Xu B, Han X, et al. Metabolomics Reveals Metabolic Changes Caused by Low-Dose 4-Tert-Octylphenol in Mice Liver. International Journal of Environmental Research and Public Health. 2018; 15(12):2686. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122686
Chicago/Turabian StyleZhou, Kun, Xingwang Ding, Jing Yang, Yanhui Hu, Yun Song, Minjian Chen, Rongli Sun, Tianyu Dong, Bo Xu, Xiumei Han, and et al. 2018. "Metabolomics Reveals Metabolic Changes Caused by Low-Dose 4-Tert-Octylphenol in Mice Liver" International Journal of Environmental Research and Public Health 15, no. 12: 2686. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122686