Response of Ruderal Species Diversity to an Urban Environment: Implications for Conservation and Management


Abstract
:1. Introduction
2. Study Site and Methods
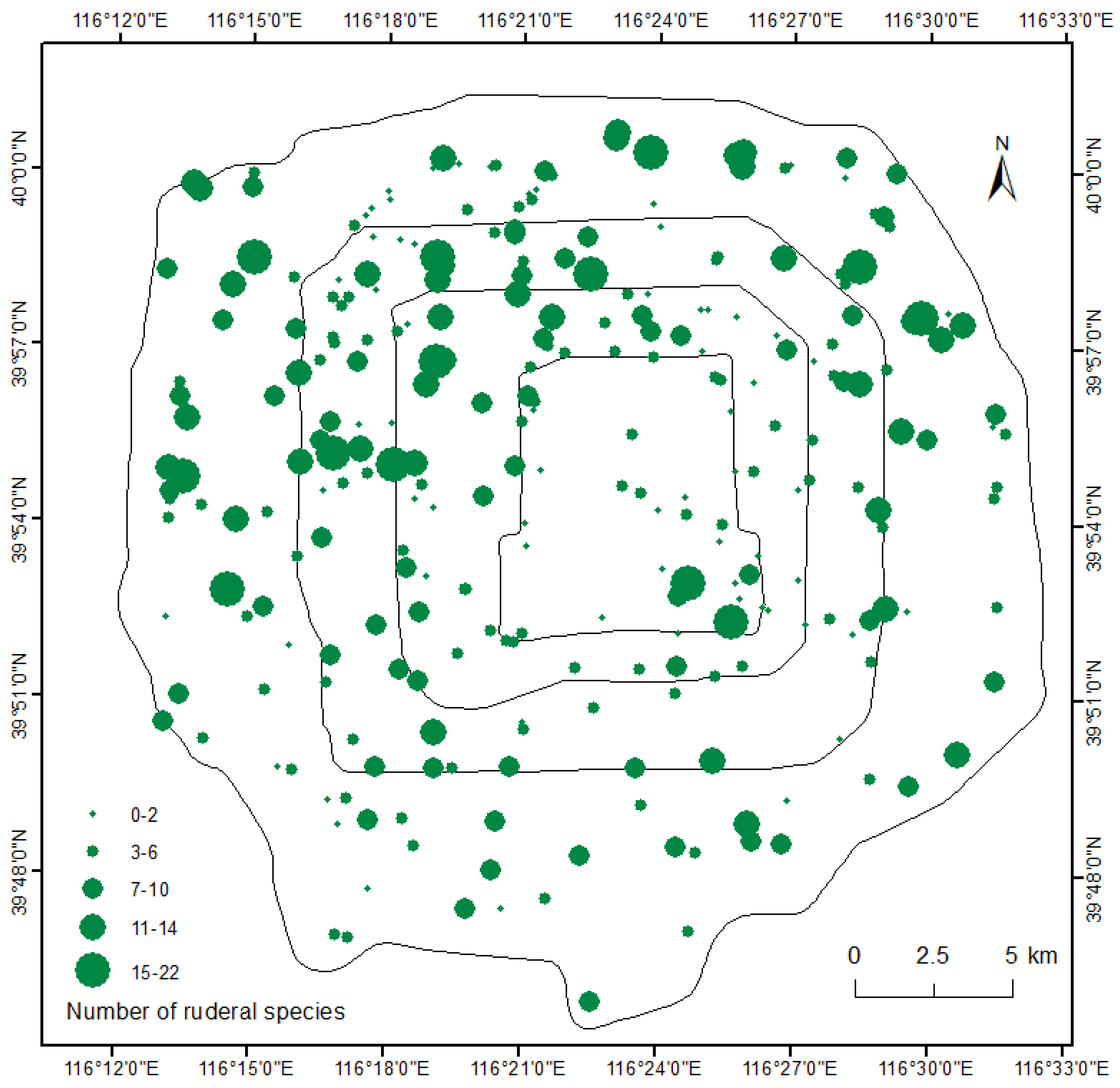
2.1. Study Site
2.2. Sampling Design and Field Investigation
2.3. Species Origin
2.4. Chorological Types
2.5. Invasive Species
2.6. Pollen-Allergenic Species
2.7. Socioeconomic Variables
2.8. Data Analysis
3. Results
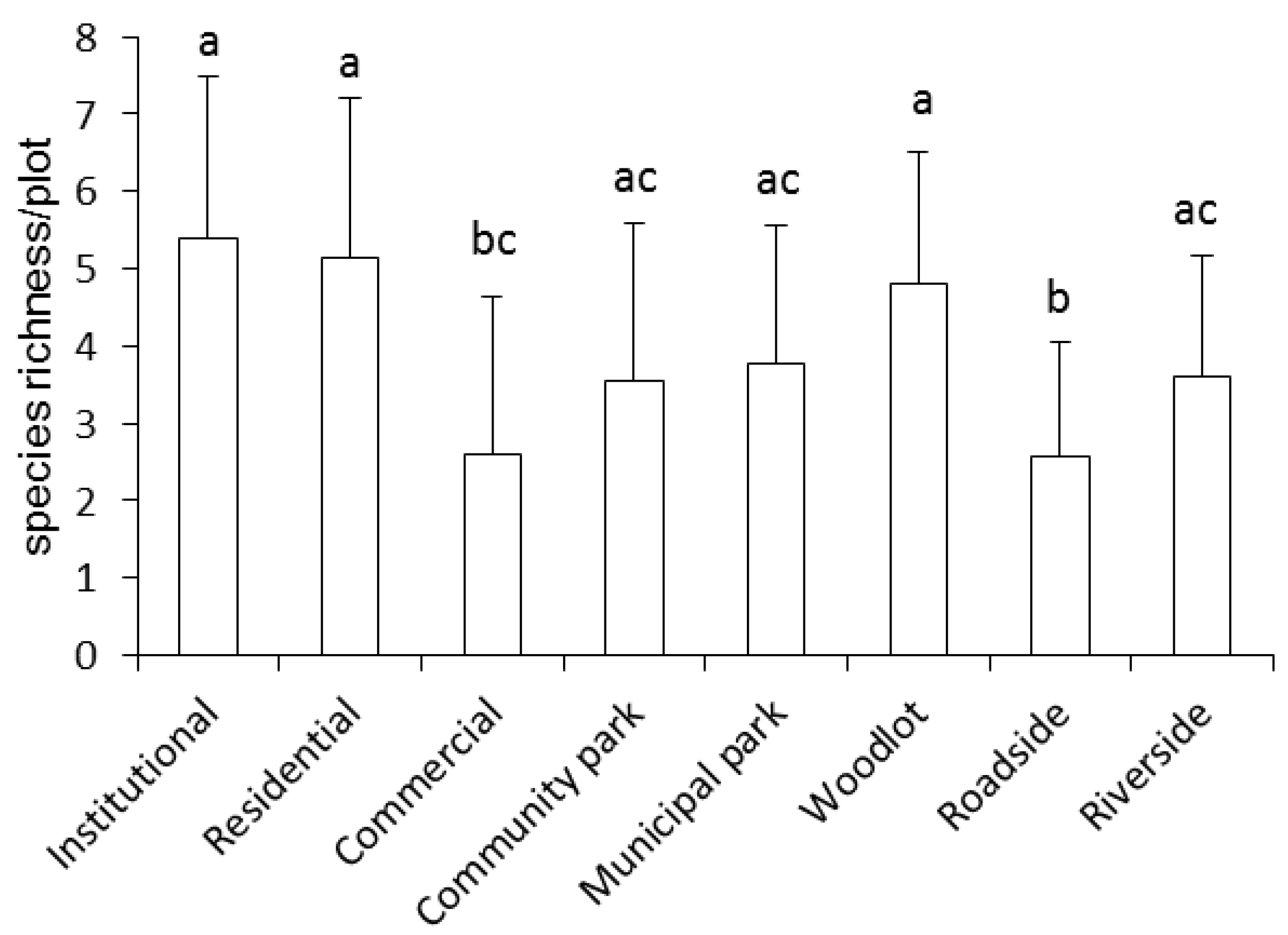
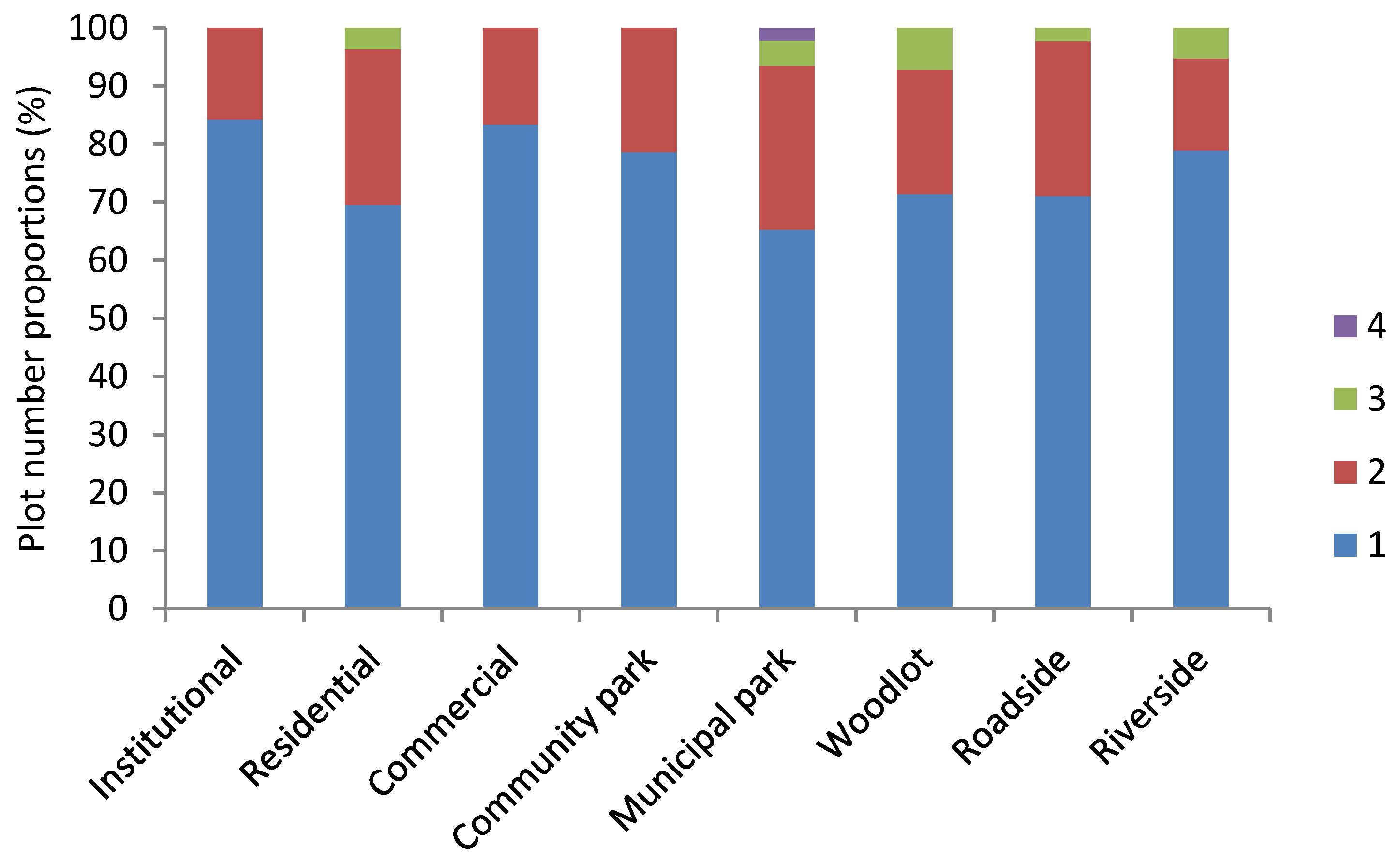
3.1. Ruderal Species Diversity
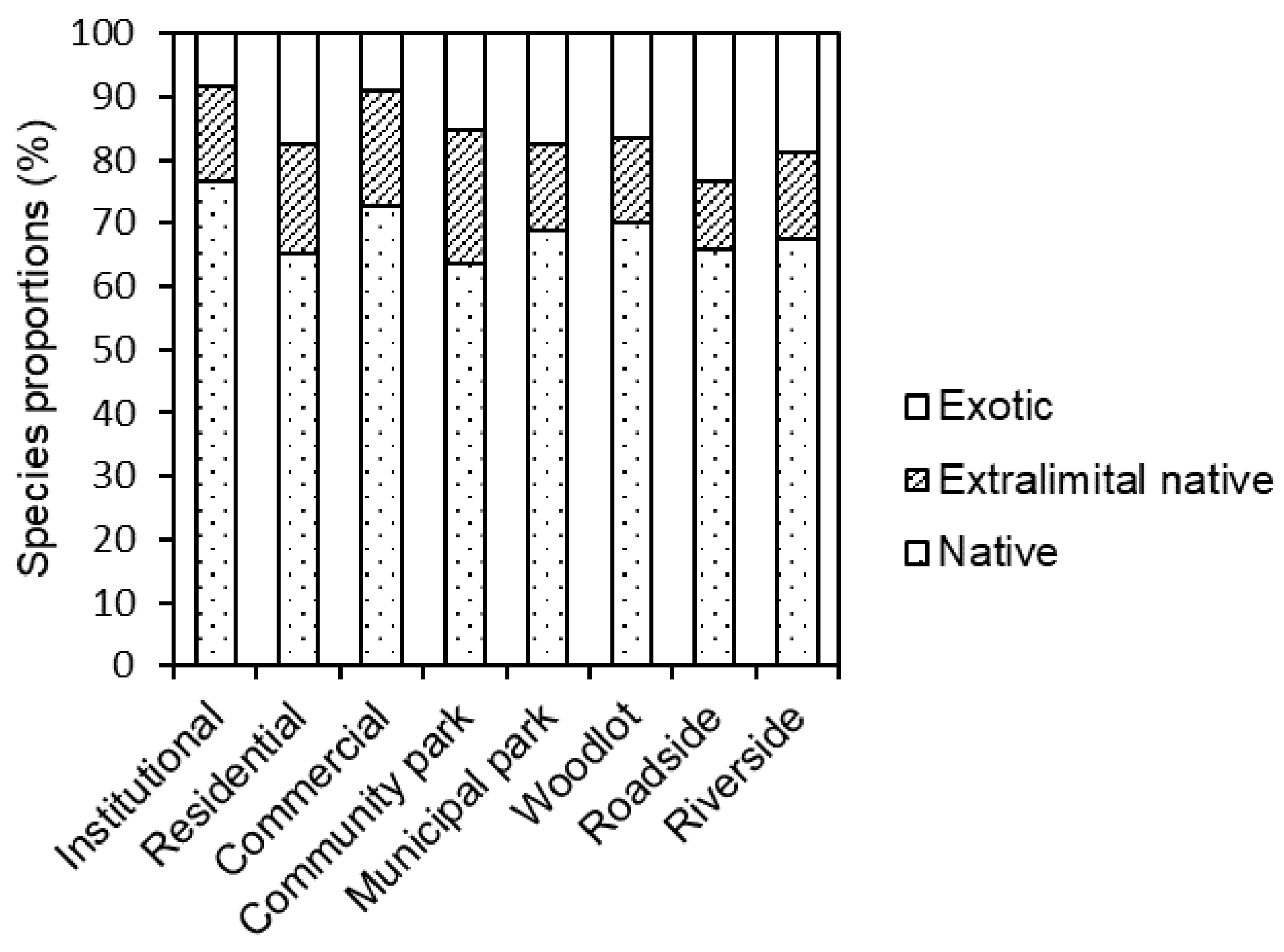
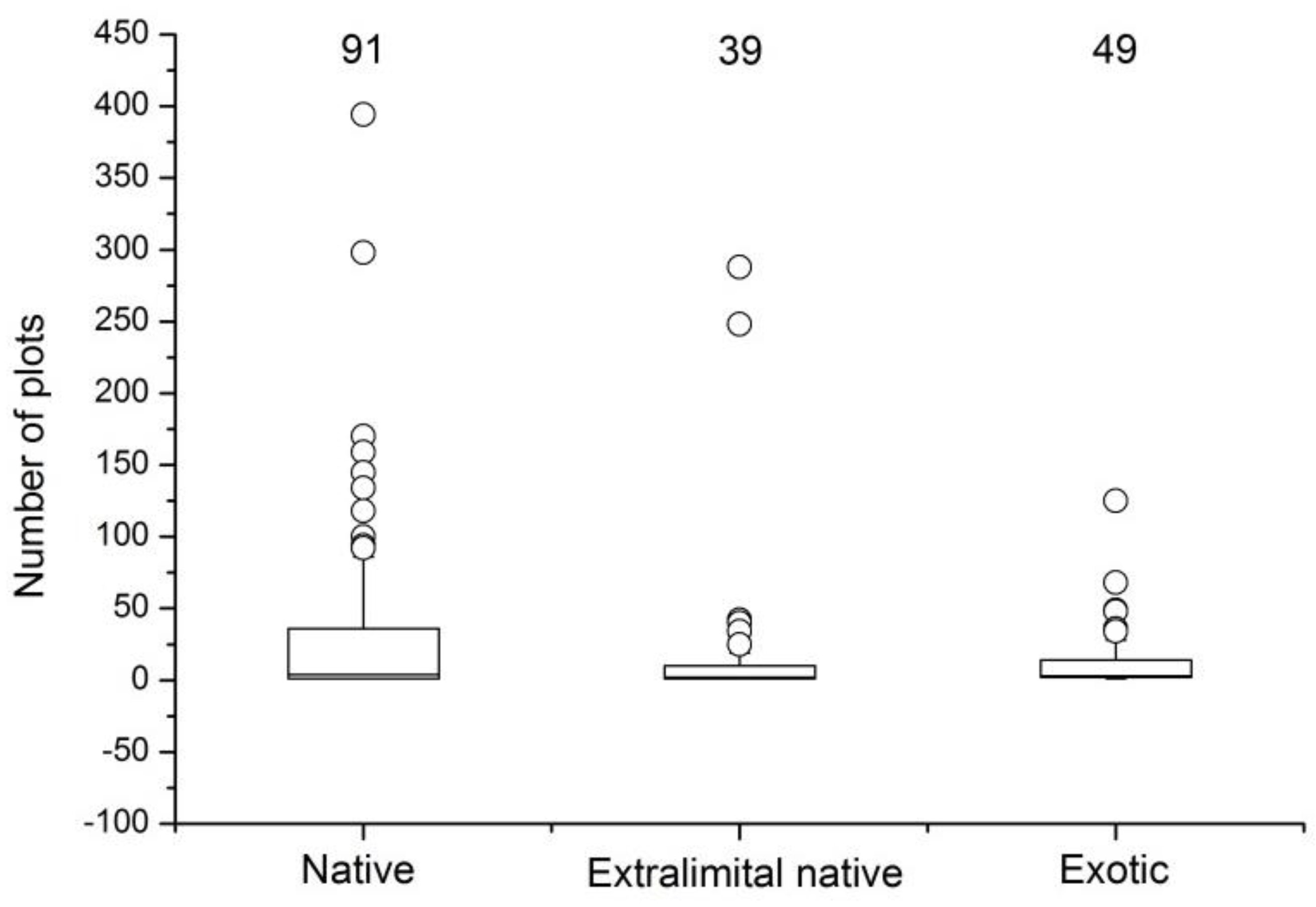
3.2. Species Origin
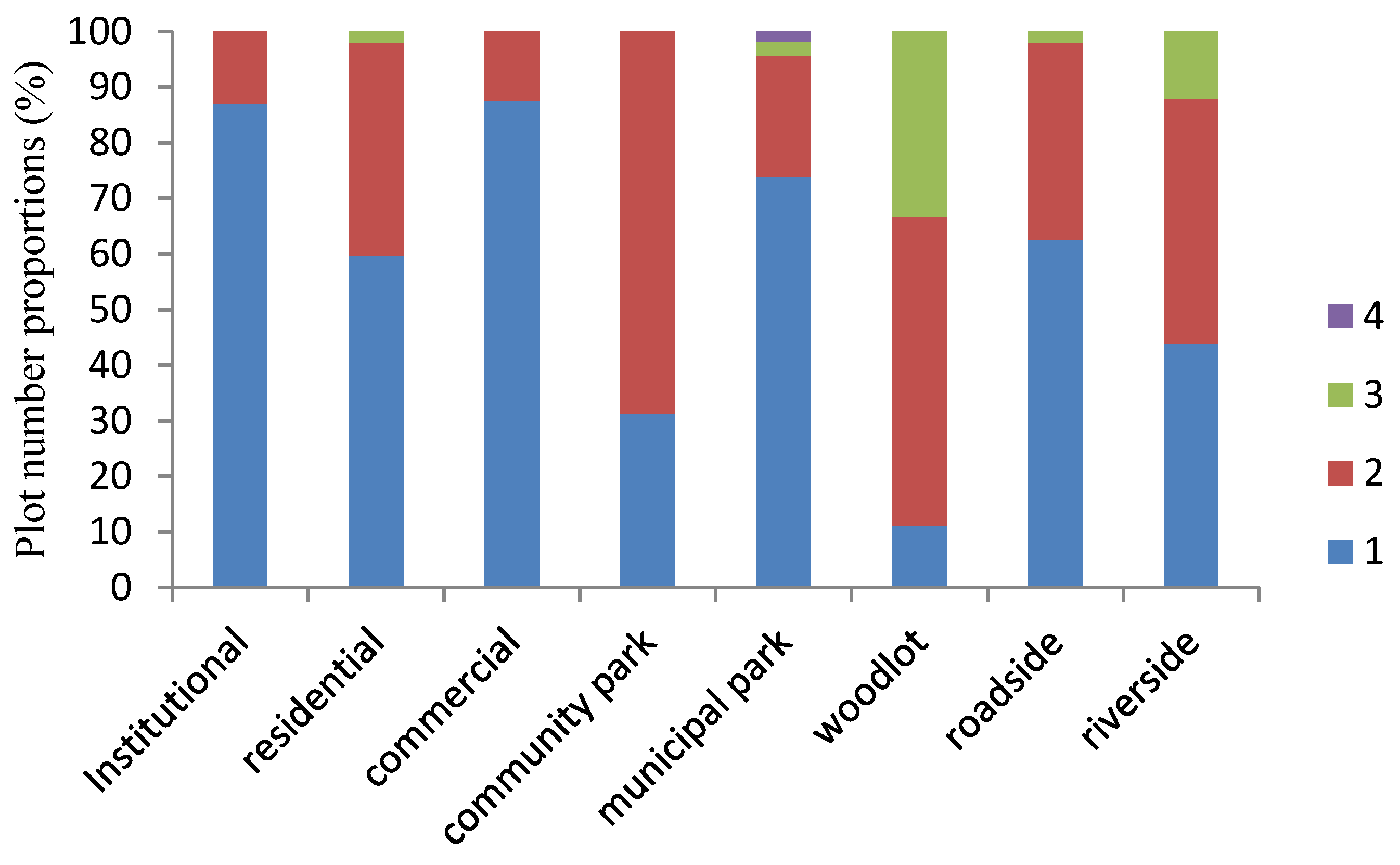
3.3. Pollen Allergenic Species
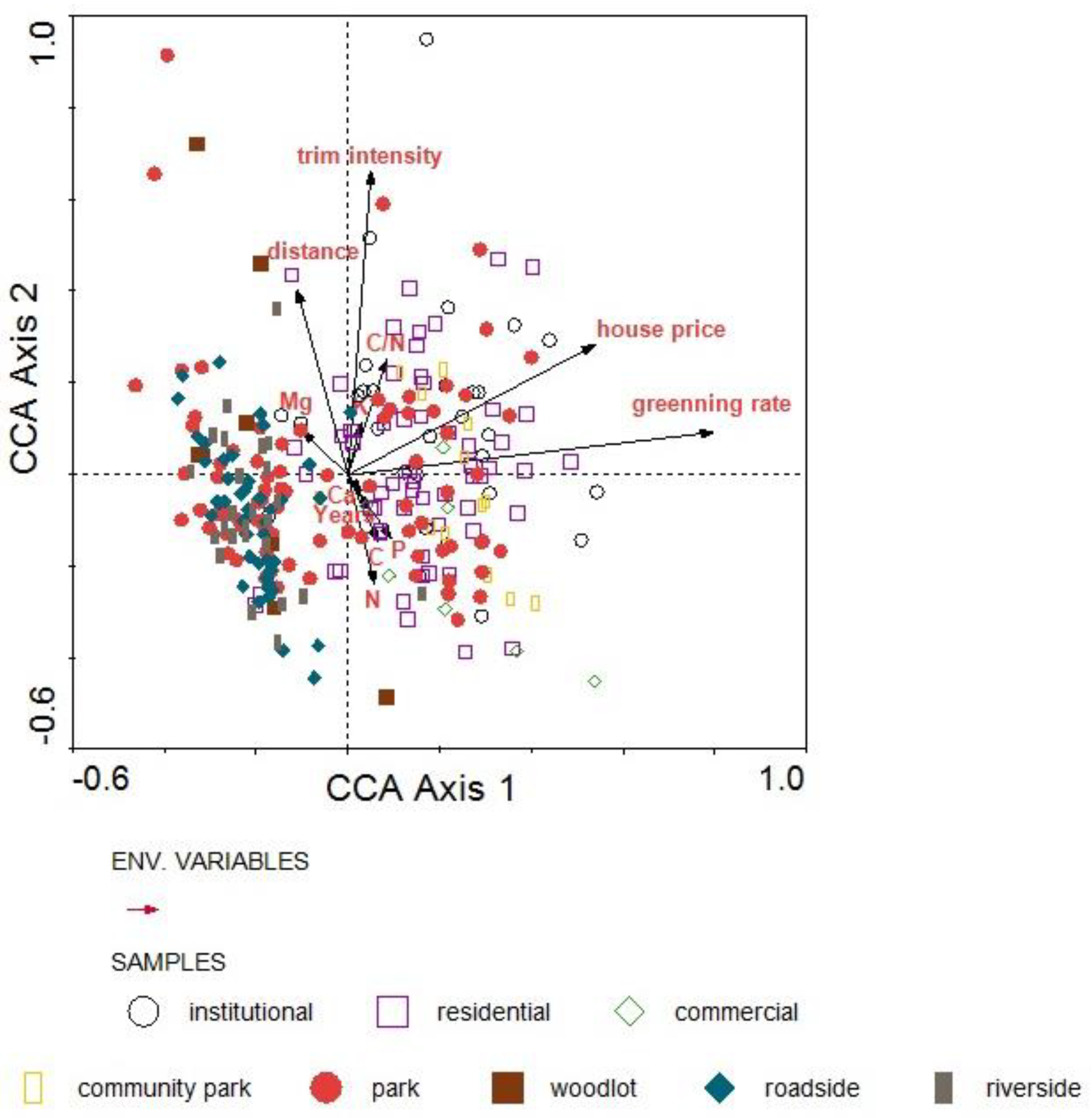
3.4. Impact Factors
4. Discussion
4.1. Composition and Diversity of Ruderal Species
4.2. Recommendations on Management
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ohsawa, M.; Da, L.J.; Otuka, T. Urban vegetation-its structure and dynamics. In Integrated Studies in Urban Ecosystems as the Basis of Urban Planning; Obara, H., Ed.; Kagawa Nutrition College: Kagawa, Japan, 1988. [Google Scholar]
- Iuliana, P.P.U.A.; Adelina, D.; Valentin, S.; Doina, C. Ecological and aesthetic role of spontaneous flora in urban sustainable landscapes development. J. Plant Dev. 2011, 18, 169–177. [Google Scholar]
- Robinson, S.L.; Lundholm, J.T. Ecosystem services provided by urban spontaneous vegetation. Urban Ecosyst. 2012, 15, 545–557. [Google Scholar] [CrossRef]
- Tredici, P.D. Spontaneous Urban Vegetation: Reflections of Change in a Globalized World. Nat. Cult. 2010, 5, 299–315. [Google Scholar] [CrossRef]
- Fischer, L.K.; Lippe, M.V.D.; Rillig, M.C.; Kowarik, I. Creating novel urban grasslands by reintroducing native species in wasteland vegetation. Biol. Conserv. 2013, 159, 119–126. [Google Scholar] [CrossRef]
- Mckinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Cervelli, E.W.; Lundholm, J.T.; Du, X. Spontaneous urban vegetation and habitat heterogeneity in Xi’an, China. Landsc. Urban Plan. 2013, 120, 25–33. [Google Scholar] [CrossRef]
- Leung, G.P.C.; Hau, B.C.H.; Corlett, R.T. Exotic plant invasion in the highly degraded upland landscape of Hong Kong, China. Biodivers. Conserv. 2008, 18, 191. [Google Scholar] [CrossRef]
- Wesche, K.; Krause, B.; Culmsee, H.; Leuschner, C. Fifty years of change in Central European grassland vegetation: Large losses in species richness and animal-pollinated plants. Biol. Conserv. 2012, 150, 76–85. [Google Scholar] [CrossRef]
- Kowarik, I. On the Role of Alien Species in Urban Flora and Vegetation. In Urban Ecology: An International Perspective on the Interaction Between Humans and Nature; Marzluff, J.M., Shulenberger, E., Endlicher, W., Alberti, M., Bradley, G., Ryan, C., Simon, U., ZumBrunnen, C., Eds.; Springer: Boston, MA, USA, 2008; pp. 321–338. [Google Scholar]
- Wania, A.; Kühn, I.; Klotz, S. Plant richness patterns in agricultural and urban landscapes in Central Germany—Spatial gradients of species richness. Landsc. Urban Plan. 2006, 75, 97–110. [Google Scholar] [CrossRef]
- Muratet, A.; Porcher, E.; Devictor, V.; Arnal, G.; Moret, J.; Wright, S.; Machon, N. Evaluation of floristic diversity in urban areas as a basis for habitat management. Appl. Veg. Sci. 2008, 11, 451–460. [Google Scholar] [CrossRef]
- Audrey, M.; Nathalie, M.; Frédéric, J.; Jacques, M.; Emmanuelle, P. The Role of Urban Structures in the Distribution of Wasteland Flora in the Greater Paris Area, France. Ecosystems 2007, 10, 661–671. [Google Scholar]
- Rink, D. Wilderness: The Nature of Urban Shrinkage? The Debate on Urban Restructuring and Restoration in Eastern Germany. Nat. Cult. 2009, 4, 275–292. [Google Scholar] [CrossRef]
- Prentis, E.; Norton, G.A. “MEADOWS”—An expert system for the establishment of diverse wildflower grasslands on derelict land in urban areas in the UK. Ecol. Eng. 1992, 1, 213–228. [Google Scholar] [CrossRef]
- Kühn, N. Intentions for the Unintentional. J. Landsc. Archit. 2006, 1, 46–53. [Google Scholar] [CrossRef]
- Muratet, A.; Pellegrini, P.; Dufour, A.B.; Arrif, T.; Chiron, F. Perception and knowledge of plant diversity among urban park users. Landsc. Urban Plan. 2015, 137, 95–106. [Google Scholar] [CrossRef]
- Hope, D.; Gries, C.; Zhu, W.; Fagan, W.F.; Redman, C.L.; Grimm, N.B.; Nelson, A.L.; Martin, C.; Kinzig, A. Socioeconomics drive urban plant diversity. Proc. Natl. Acad. Sci. USA 2003, 100, 8788–8792. [Google Scholar] [CrossRef] [Green Version]
- Turner, K.; Lefler, L.; Freedman, B. Plant communities of selected urbanized areas of Halifax, Nova Scotia, Canada. Landsc. Urban Plan. 2005, 71, 191–206. [Google Scholar] [CrossRef]
- Lundholm, J.T.; Marlin, A. Habitat origins and microhabitat preferences of urban plant species. Urban Ecosyst. 2006, 9, 139–159. [Google Scholar] [CrossRef]
- Jim, C.Y.; Chen, W.Y. Bioreceptivity of buildings for spontaneous arboreal flora in compact city environment. Urban For. Urban Green. 2011, 10, 19–28. [Google Scholar] [CrossRef]
- Chen, X.; Wang, W.; Liang, H.; Liu, X.; Da, L. Dynamics of ruderal species diversity under the rapid urbanization over the past half century in Harbin, Northeast China. Urban Ecosyst. 2014, 17, 455–472. [Google Scholar] [CrossRef]
- Merunková, K.; Chytrý, M. Environmental control of species richness and composition in upland grasslands of the southern Czech Republic. Plant Ecol. 2012, 213, 591–602. [Google Scholar] [CrossRef]
- Krasniqi, S.; Kopali, A.; Rota, E. Study of plant diversity in the river upstream of Prizreni Lumbardh. Alban. J. Agric. Sci. 2013, 12, 425. [Google Scholar]
- Budeanu, O. Management and assessment of medicinal plants from spontaneous flora in the Republic of Moldova. Bull. Transilv. Univ. Bras. 2013, 6, 81–86. [Google Scholar]
- Miller, J.R. Biodiversity conservation and the extinction of experience. Trends Ecol. Evol. 2005, 20, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Yuebo, S.U.; Wan, W.; Liu, W.; Zhang, H.; Sun, X.; Ouyang, Z.; Wang, X. Urban Plant Diversity in Relation to Land Use Types in Built-up Areas of Beijing. Chin. Geogr. Sci. 2018, 100–110. [Google Scholar] [CrossRef]
- He, J.Q. Exotic Plants in China; Shanghai Scientific & Technical Publishers: Shanghai, China, 2011. (In Chinese) [Google Scholar]
- He, S.Y.; Xing, Q.H.; Yin, Z.T.; Jiang, X.F. Flora of Beijing; Beijing Publishing House: Beijing, China, 1984; Volume 1. [Google Scholar]
- Compilation Committee of the Flora of China. Flora of China; Science Press: Beijing, China, 2004. (In Chinese) [Google Scholar]
- Wu, Z.Y. The areal-types of Chinese genera of seed plants. Acta Bot. Yunnanica 1991, 4, 1–139. [Google Scholar]
- Wan, F.H.; Liu, Q.R.; Xie, M. Blological Invasions: Color Illustrations of Invasive Alien Plants in China; Science Press: Beijing, China, 2012. (In Chinese) [Google Scholar]
- Liu, Q.R.; Yu, M.; Zhou, Y.L. Study of alien plants in Beijing. J. Beijing Norm. Univ. (Nat. Sci.) 2002, 38, 399–404. [Google Scholar]
- Investigational Team on Airborne and Allergenic Pollen Grains in China. An Investigation on Airborne and Allergenic Pollen Grains in China; China People Press: Beijing, China, 1991. (In Chinese) [Google Scholar]
- Beijing Municipal Bureau of Statistics. Beijing Statistical Yearbook; China Statistics Press: Beijing, China, 2014. (In Chinese) [Google Scholar]
- Braak, C.J.F.T.; Smilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); Wageningen University: Ithaca, NY, USA, 2002. [Google Scholar]
- Chen, K.X. Study on Types, Distribution Pattern and Seasonal Change of Weed Communities in Urban Ecosystem of Shanghai; East China Normal Univesrity: Shanghai, China, 2005. (In Chinese) [Google Scholar]
- Li, J.H. The Medieinal Plants of Landscape Greening in Northeast Cities. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2007. [Google Scholar]
- Stewart, G.H.; Ignatieva, M.E.; Meurk, C.D.; Buckley, H.; Horne, B.; Braddick, T. URban Biotopes of Aotearoa New Zealand (URBANZ) (I): Composition and diversity of temperate urban lawns in Christchurch. Urban Ecosyst. 2009, 12, 233–248. [Google Scholar] [CrossRef]
- Zhao, J.; Ouyang, Z.; Hua, Z.; Zhou, W.; Wang, X.; Xu, W.; Ni, Y. Erratum to: Plant species composition in green spaces within the built-up areas of Beijing, China. Plant Ecol. 2010, 209, 189–204. [Google Scholar] [CrossRef]
- Meurk, C.D.; Sullivan, J.; Mcwilliam, W. Vegetation History and Dynamics in New Zealand: Future Scenarios and Improved Trajectories Towards Restoring Natural Patterns. In Vegetation Structure and Function at Multiple Spatial, Temporal and Conceptual Scales; Springer: Berlin, Germany, 2016; pp. 517–528. [Google Scholar]
- Batten, L.A. Breeding Bird Species Diversity in Relation to Increasing Urbanisation. Bird Study 1972, 19, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Livingston, M.; Shaw, W.W.; Harris, L.K. A model for assessing wildlife habitats in urban landscapes of eastern Pima County, Arizona (USA). Landsc. Urban Plan. 2003, 64, 131–144. [Google Scholar] [CrossRef]
- Flory, S.; Clay, K. Non-native grass invasion alters native plant composition in experimental communities. Biol. Invasions 2010, 12, 1285–1294. [Google Scholar] [CrossRef]
- Davies, K.W. Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass. Oecologia 2011, 167, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Bormann, F.H.; Balmori, D.; Geballe, G.T.; Vernegaard, L. Redesigning the American Lawn: A Search for Environmental Harmony; Yale University Press: New Haven, CT, USA, 2001. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Institutional (%) | Residential (%) | Commercial (%) | Community Park (%) | Municipal Park (%) | Woodlot (%) | Roadside (%) | Riverside (%) |
|---|---|---|---|---|---|---|---|---|---|
| Oxalidaceae | Oxalis corniculata | 36.3 | 28.3 | 24.3 | 28.6 | 32.5 | 6.5 | 20.3 | 13.4 |
| Gramineae | Setaria viridis | 17.6 | 43.8 | 27.0 | 33.3 | 37.3 | 64.5 | 25.0 | 53.6 |
| Violaceae | Viola philippica | 14.7 | 17.2 | — | 14.3 | 16.6 | — | 6.3 | 14.4 |
| Violaceae | Viola pekinensis | 13.7 | 19.3 | 10.8 | — | 20.5 | 25.8 | 11.5 | 8.2 |
| Chenopodiaceae | Chenopodium glaucum | 8.8 | 27.5 | 18.9 | 21.4 | 25.3 | 67.7 | 13.0 | 26.8 |
| Asteraceae | Taraxacum mongolicum | 7.8 | 15.0 | 13.5 | 40.5 | 14.0 | 12.9 | 4.7 | 4.1 |
| Asteraceae | Inula japonica | 7.8 | 11.6 | 2.7 | 4.8 | 11.7 | 16.1 | 5.2 | — |
| Gramineae | Poa pratensis | 9.8 | 11.2 | 18.9 | 14.3 | 10.1 | 12.9 | 7.3 | 2.1 |
| Gramineae | Digitaria sanguinalis | 5.9 | 13.7 | 8.1 | 28.6 | 13.6 | 9.7 | 23.4 | 26.8 |
| Asteraceae | Conyza canadensis | 4.9 | 6.4 | 10.8 | 9.5 | 2.9 | 12.9 | 2.1 | 2.1 |
| Euphorbiaceae | Acalypha australis | 4.9 | 5.2 | 5.4 | 9.5 | 8.1 | 3.2 | 6.8 | 5.2 |
| Amaranthaceae | Amaranthus retroflexus | — | 10.3 | 2.7 | 7.1 | 3.6 | 9.7 | 6.8 | 9.3 |
| Asteraceae | Ixeridium sonchifolium | 2.9 | 11.2 | 13.5 | 4.8 | 7.1 | 16.1 | 3.1 | 1.0 |
| Plantaginaceae | Plantago depressa | 3.9 | 7.3 | 5.4 | 11.9 | 14.6 | 16.1 | 3.1 | 4.1 |
| Portulacaceae | Portulaca oleracea | — | 10.3 | 10.8 | 9.5 | 14.9 | — | 13.5 | 7.2 |
| Solanaceae | Solanum nigrum | 2.9 | 10.7 | 8.1 | 16.7 | 7.1 | 19.4 | 3.6 | 7.2 |
| Euphorbiaceae | Euphorbia humifusa | 2.9 | 4.3 | 2.7 | 4.8 | 5.2 | 3.2 | 8.9 | 9.3 |
| Gramineae | Eleusine indica | 1.0 | 15.5 | 8.1 | 14.3 | 13.0 | 12.9 | 13.0 | — |
| Rubiaceae | Rubia cordifolia | 2.9 | 6.0 | — | 8.1 | 8.1 | 35.5 | 3.1 | 14.4 |
| Labiatae | Leonurus artemisia | 1.0 | 6.0 | — | 14.3 | 5.5 | — | 4.7 | 9.3 |
| Asclepiadaceae | Cynanchum chinense | 1.0 | 6.9 | 2.7 | — | 9.0 | 16.1 | 2.6 | 9.3 |
| Gramineae | Buchloe dactyloides | — | 4.7 | 2.7 | — | 3.6 | — | 9.9 | 6.2 |
| Convolvulaceae | Pharbitis nil | — | 2.6 | — | 2.4 | 1.3 | — | 2.6 | 11.3 |
| Convolvulaceae | Calystegia hederacea | 2.0 | 1.7 | 5.4 | — | 4.9 | 6.5 | 3.1 | 10.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.; Yu, F.; Ren, Y.; Liu, D.; Li, J.; Ouyang, Z.; Wang, X. Response of Ruderal Species Diversity to an Urban Environment: Implications for Conservation and Management. Int. J. Environ. Res. Public Health 2018, 15, 2832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122832
Guo P, Yu F, Ren Y, Liu D, Li J, Ouyang Z, Wang X. Response of Ruderal Species Diversity to an Urban Environment: Implications for Conservation and Management. International Journal of Environmental Research and Public Health. 2018; 15(12):2832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122832
Chicago/Turabian StyleGuo, Peipei, Fang Yu, Yuan Ren, Dong Liu, Jia Li, Zhiyun Ouyang, and Xiaoke Wang. 2018. "Response of Ruderal Species Diversity to an Urban Environment: Implications for Conservation and Management" International Journal of Environmental Research and Public Health 15, no. 12: 2832. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15122832