Quorum Sensing versus Quenching Bacterial Isolates Obtained from MBR Plants Treating Leachates from Municipal Solid Waste

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wastewater Treatment Plants
2.2. Sampling and Strain Isolation
2.3. Identification of Isolates
2.4. Detection of AHL Production by Agar Plate Assay
2.5. Surface Adherence Capability of AHL Producers
2.6. Detection of Quorum Quenching Activity by the Agar Plate Assay
2.7. QQ Strains Addition to Mixed Liquors and Monospecific Bacterial Biofilms
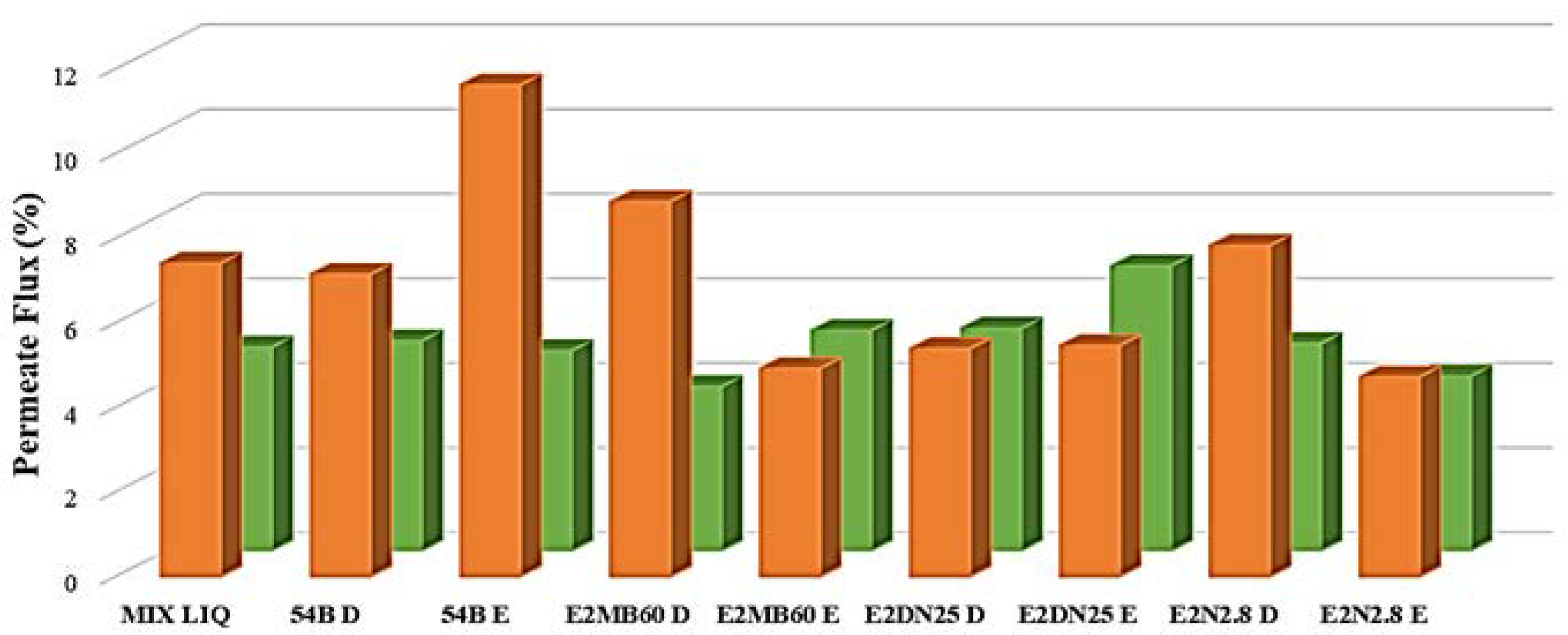
2.8. Effect of QQ Strains on Ultrafiltration Fluxes
2.9. Quantification of Enzymatic Activities
3. Results
3.1. Identification and Characterization of Bacterial Isolates
3.2. AHL Producers and Potential Adherence to Surfaces
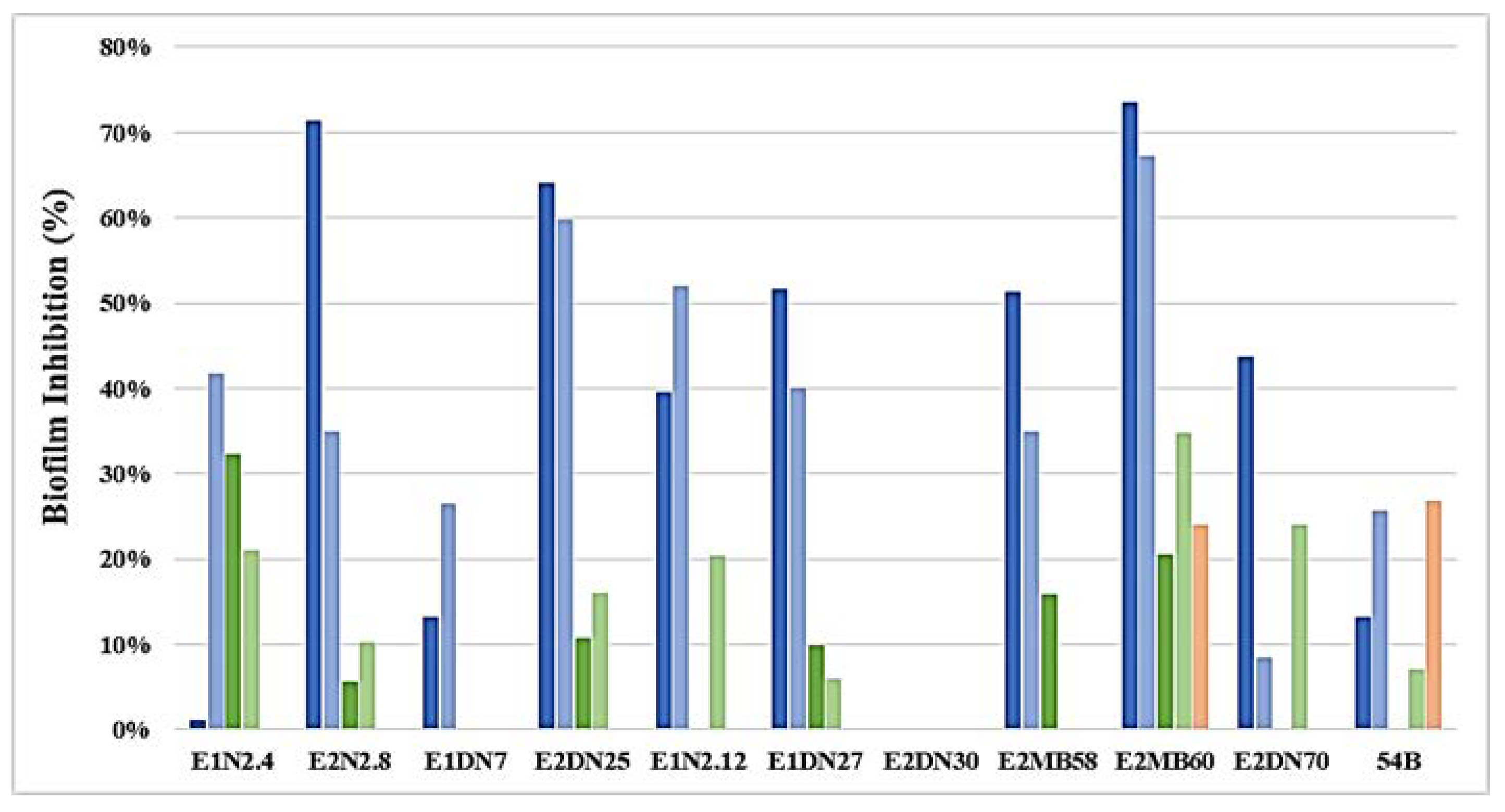
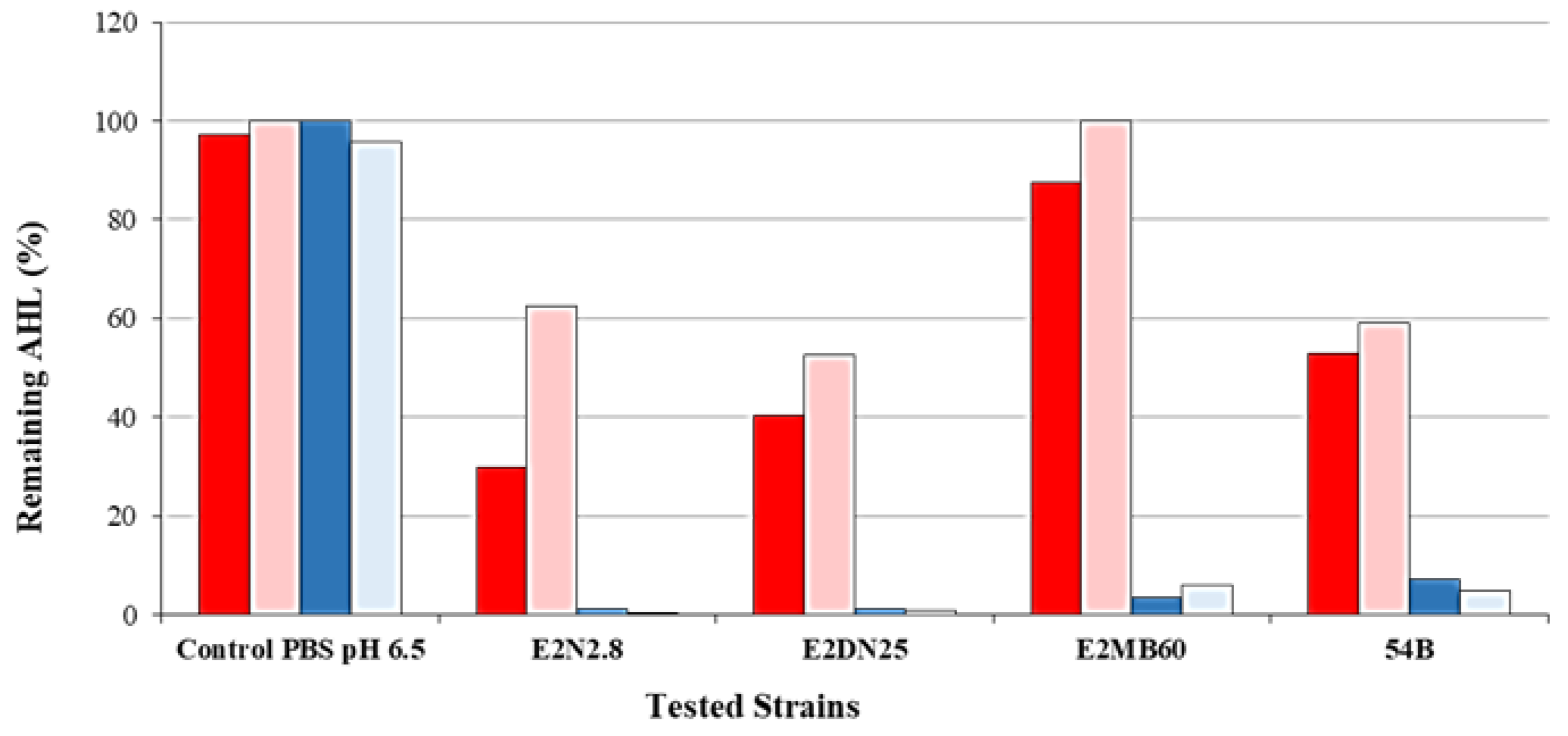
3.3. Inhibition of AHL and QQ Assays
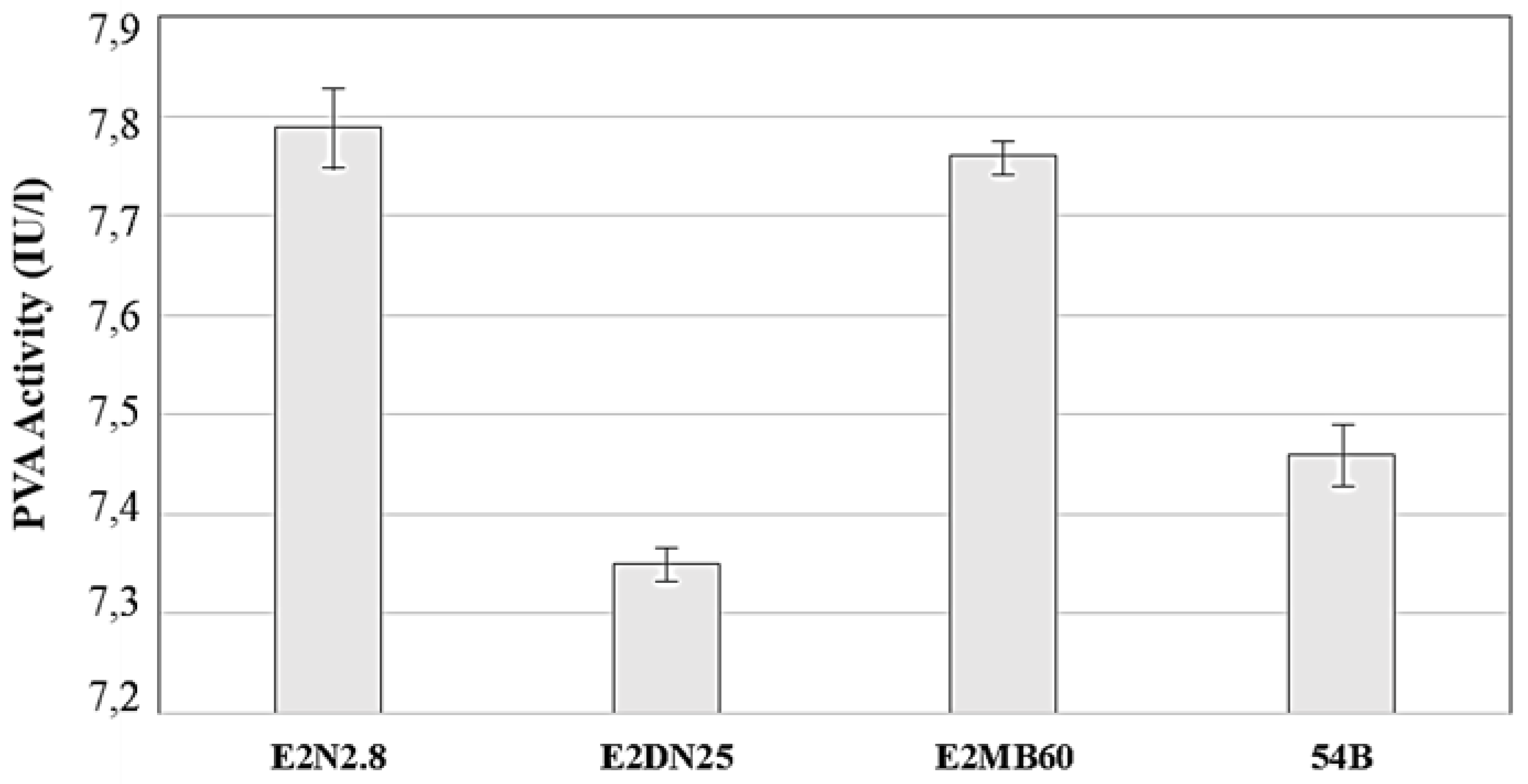
3.4. Enzymatic Activities of Selected QQ Strains
3.4.1. Lactonase Activity
3.4.2. Acylase Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Brown, S.P.; Johnstone, R.A. Cooperation in the dark: Signalling and collective action in quorum-sensing bacteria. Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Hogan, D.A. Quorum sensing: Alcohols in a social situation. Curr. Biol. 2006, 16, 457–458. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Parkinson, K.; Thompson, C.R.L. What can microbial genetics teach sociobiology? Trends Genet. 2007, 23, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria—The Luxr-Luxi family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, K.; Paul, D.; Kim, Y.S.; Kweon, J.H. 2(5H)-Furanone: A prospective strategy for biofouling-control in membrane biofilm bacteria by quorum sensing inhibition. Braz. J. Microbiol. 2010, 41, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, C. Exploiting quorum sensing interfering strategies in Gram-negative bacteria for the enhancement of environmental applications. Front. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- LaSarre, B.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef] [PubMed]
- Christiaen, S.E.A.; Brackman, G.; Nelis, H.J.; Coenye, T. Isolation and identification of quorum quenching bacteria from environmental samples. J. Microbiol. Methods 2011, 87, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Biswa, P.; Doble, M. Production of acylated homoserine lactone by Gram-positive bacteria isolated from marine water. FEMS Microbiol. Lett. 2013, 343, 34–41. [Google Scholar] [CrossRef] [PubMed]
- De Decker, S.; Reynaud, Y.; Saulnier, D. First molecular evidence of cross-species induction of metalloprotease gene expression in Vibrio strains pathogenic for Pacific oyster Crassostrea gigas involving a quorum sensing system. Aquaculture 2013, 392, 1–7. [Google Scholar] [CrossRef]
- Pande, G.S.J.; Scheie, A.A.; Benneche, T.; Wille, M.; Sorgeloos, P.; Bossier, P.; Defoirdt, T. Quorum sensing-disrupting compounds protect larvae of the giant freshwater prawn Macrobrachium rosenbergii from Vibrio harveyi infection. Aquaculture 2013, 406, 121–124. [Google Scholar] [CrossRef]
- Swift, S.; Downie, J.A.; Whitehead, N.A.; Barnard, A.M.L.; Salmond, G.P.C.; Williams, P. Quorum sensing as a population-density-dependent determinant of bacterial physiology. Adv. Microb. Physiol. 2001, 45, 199–270. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.; Winzer, K.; Chan, W.C.; Camara, M. Look who’s talking: Communication and quorum sensing in the bacterial world. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1119–1134. [Google Scholar] [CrossRef] [PubMed]
- Judd, S. The MBR Book: Principles and Applications of Membrane Bioreactors for Water and Wastewater Treatmen, 1st ed.; Elsevier Science: New York, NY, USA, 2006; ISBN 978-1-85617-481-7. [Google Scholar]
- Lesjean, B.; Huisjes, E.H. Survey of the European MBR market: Trends and perspectives. Desalination 2008, 231, 71–81. [Google Scholar] [CrossRef]
- Jarusutthirak, C.; Amy, G. Role of soluble microbial products (SMP) in membrane fouling and flux decline. Environ. Sci. Technol. 2006, 40, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Le-Clech, P.; Chen, V.; Fane, T.A.G. Fouling in membrane bioreactors used in wastewater treatment. J. Membr. Sci. 2006, 284, 17–53. [Google Scholar] [CrossRef]
- Meng, F.; Chae, S.-R.; Drews, A.; Kraume, M.; Shin, H.-S.; Yang, F. Recent advances in membrane bioreactors (MBRs): Membrane fouling and membrane material. Water Res. 2009, 43, 1489–1512. [Google Scholar] [CrossRef] [PubMed]
- Lade, H.; Paul, D.; Kweon, J.H. Isolation and molecular characterization of biofouling bacteria and profiling of quorum sensing signal molecules from membrane bioreactor activated sludge. Int. J. Mol. Sci. 2014, 15, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 1998, 280, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Yeon, K.-M.; Cheong, W.-S.; Oh, H.-S.; Lee, W.-N.; Hwang, B.-K.; Lee, C.-H.; Beyenal, H.; Lewandowski, Z. Quorum sensing: A new biofouling control paradigm in a membrane bioreactor for advanced wastewater treatment. Environ. Sci. Technol. 2009, 43, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Drews, A. Membrane fouling in membrane bioreactors-Characterisation, contradictions, cause and cures. J. Membr. Sci. 2010, 363, 1–28. [Google Scholar] [CrossRef]
- Kim, S.-R.; Oh, H.-S.; Jo, S.-J.; Yeon, K.-M.; Lee, C.-H.; Lim, D.-J.; Lee, C.-H.; Lee, J.-K. Biofouling control with bead-entrapped quorum quenching bacteria in membrane bioreactors: Physical and biological effects. Environ. Sci. Technol. 2013, 47, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, S.-K.; Kwon, H.; Lee, S.H.; Lee, K.; Chang Hyun, N.; Jo, S.J.; Oh, H.-S.; Park, P.-K.; Choo, K.-H.; et al. Crossing the border between laboratory and field: Bacterial quorum quenching for anti-biofouling strategy in an MBR. Environ. Sci. Technol. 2016, 50, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Oger, P.M.; Uroz, S. Screening for N-AHSL-based-signaling interfering enzymes. In Metagenomics: Methods and Protocols; Streit, W.R., Daniel, R., Eds.; Humana Press: New York, NY, USA, 2010; Volume 668, pp. 203–217. ISBN 978-1-60761-822-5. [Google Scholar]
- Oh, H.-S.; Yeon, K.-M.; Yang, C.-S.; Kim, S.-R.; Lee, C.-H.; Park, S.Y.; Han, J.Y.; Lee, J.K. Control of membrane biofouling in MBR for wastewater treatment by quorum quenching bacteria encapsulated in microporous membrane. Environ. Sci. Technol. 2012, 46, 4877–4884. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.-H.; Wang, L.-H.; Zhang, L.-H. Quorum-quenching microbial infections: Mechanisms and implications. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Mayer, C.; Muras, A.; Otero, A. Silencing bacterial communication through enzymatic quorum sensing inhibition. In Quorum Sensing vs Quorum Quenching: A Battle with No End in Sight; Kalia, V.C., Ed.; Springer: New Delhi, India, 2015; ISBN 978-81-322-1981-1. [Google Scholar]
- Leadbetter, J.R.; Greenberg, E.P. Metabolism of acyl-homoserine lactone quorum-sensing signals by Variovorax paradoxus. J. Bacteriol. 2000, 182, 6921–6926. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Diggle, S.P.; Heeb, S.; Camara, M.; Otero, A. Quorum quenching activity in Anabaena sp. PCC 7120: Identification of AiiC, a novel AHL-acylase. FEMS Microbiol. Lett. 2008, 280, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kang, H.O.; Jang, H.S.; Lee, J.K.; Koo, B.T.; Yum, D.Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl. Environ. Microbiol. 2005, 71, 2632–2641. [Google Scholar] [CrossRef] [PubMed]
- Sio, C.F.; Otten, L.G.; Cool, R.H.; Diggle, S.P.; Braun, P.G.; Bos, R.; Daykin, M.; Camara, M.; Williams, P.; Quax, W.J. Quorum quenching by an N-acyl-homoserine lactone acylase from Pseudomonas aeruginosa PAO1. Infect. Immun. 2006, 74, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Sunder, A.V.; Kumar, A.; Naik, N.; Pundle, A.V. Characterization of a new Bacillus cereus ATUAVP1846 strain producing penicillin V acylase, and optimization of fermentation parameters. Ann. Microbiol. 2012, 62, 1287–1293. [Google Scholar] [CrossRef]
- Kang, B.R.; Lee, J.H.; Ko, S.J.; Lee, Y.H.; Cha, J.S.; Cho, B.H.; Kim, Y.C. Degradation of acyl-homoserine lactone molecules by Acinetobacter sp. strain C1010. Can. J. Microbiol. 2004, 50, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Hwang, B.J.; Shin, M.H.; Kim, J.A.; Kim, H.K.; Lee, J.K. N-acylhomoserine lactonase producing Rhodococcus spp. with different AHL-degrading activities. FEMS Microbiol. Lett. 2006, 261, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Oger, P.M.; Chapelle, E.; Adeline, M.-T.; Faure, D.; Dessaux, Y. A Rhodococcus qsdA-encoded enzyme defines a novel class of large-spectrum quorum-quenching lactonases. Appl. Environ. Microbiol. 2008, 74, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Thomas, P.W.; Momb, J.; Hoang, Q.Q.; Petsko, G.A.; Ringe, D.; Fast, W. Structure and specificity of a quorum-quenching lactonase (AiiB) from Agrobacterium tumefaciens. Biochemistry 2007, 46, 11789–11799. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Xu, J.L.; Li, X.Z.; Zhang, L.H. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc. Natl. Acad. Sci. USA 2000, 97, 3526–3531. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Gusti, A.R.; Zhang, Q.; Xu, J.L.; Zhang, L.H. Identification of quorum-quenching N-acyl homoserine lactonases from Bacillus species. Appl. Environ. Microbiol. 2002, 68, 1754–1759. [Google Scholar] [CrossRef] [PubMed]
- d’Angelo-Picard, C.; Faure, D.; Penot, I.; Dessaux, Y. Diversity of N-acyl homoserine lactone-producing and -degrading bacteria in soil and tobacco rhizosphere. Environ. Microbiol. 2005, 7, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.H.; Zhang, L.H. Quorum sensing and quorum-quenching enzymes. J. Microbiol. 2005, 43, 101–109. [Google Scholar] [PubMed]
- Pana, J.; Huang, T.; Yao, F.; Huang, Z.; Powell, C.A.; Qiu, S.; Guan, X. Expression and characterization of aiiA gene from Bacillus subtilis BS-1. Microbiol. Res. 2008, 163, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Momb, J.; Thomas, P.W.; Moulin, A.; Petsko, G.A.; Fast, W.; Ringe, D. Mechanism of the quorum-quenching lactonase (AiiA) from Bacillus thuringiensis. 1. Product-bound structures. Biochemistry 2008, 47, 7706–7714. [Google Scholar] [CrossRef] [PubMed]
- Lade, H.; Paul, D.; Kweon, J.H. N-acyl homoserine lactone-mediated quorum sensing with special reference to use of quorum quenching bacteria in membrane biofouling control. BioMed Res. Int. 2014, 2014, 162584. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, A.; Russo, L.; Belgiorno, V. Combined anaerobic/aerobic treatment of OFMSW: Performance evaluation using mass balances. Chem. Eng. J. 2015, 267, 16–24. [Google Scholar] [CrossRef]
- Zuriaga-Agusti, E.; Mendoza-Roca, J.A.; Bes-Pia, A.; Alonso-Molina, J.L.; Fernandez-Gimenez, E.; Alvarez-Requena, C.; Munagorri-Manueco, F.; Ortiz-Villalobos, G. Comparison between mixed liquors of two side-stream membrane bioreactors treating wastewaters from waste management plants with high and low solids anaerobic digestion. Water Res. 2016, 100, 517–525. [Google Scholar] [CrossRef] [PubMed]
- Young, J.P.; Downer, H.L.; Eardly, B.D. Phylogeny of the phototrophic Rhizobium strain BTAi1 by polymerase chain reaction-based sequencing of a 16S rRNA gene segment. J. Bacteriol. 1991, 173, 2271–2277. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL-W—Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Ravn, L.; Christensen, A.B.; Molin, S.; Givskov, M.; Gram, L. Methods for detecting acylated homoserine lactones produced by Gram-negative bacteria and their application in studies of AHL-production kinetics. J. Microbiol. Methods 2001, 44, 239–251. [Google Scholar] [CrossRef]
- Watson, B.; Currier, T.C.; Gordon, M.P.; Chilton, M.D.; Nester, E.W. Plasmid required for virulence of Agrobacterium tumefaciens. J. Bacteriol. 1975, 123, 255–264. [Google Scholar] [PubMed]
- Steindler, L.; Venturi, V. Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 2007, 266, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005, 1, 1B. 1.1–1B. 1.17. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Romero, M.; Martin-Cuadrado, A.-B.; Roca-Rivada, A.; Maria Cabello, A.; Otero, A. Quorum quenching in cultivable bacteria from dense marine coastal microbial communities. FEMS Microbiol. Ecol. 2011, 75, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Muras, A.; Mayer, C.; Bujan, N.; Magarinos, B.; Otero, A. In vitro quenching of fish pathogen Edwardsiella tarda AHL production using marine bacterium Tenacibaculum sp. strain 20J cell extracts. Dis. Aquat. Org. 2014, 108, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Ortori, C.A.; Atkinson, S.; Chhabra, S.R.; Camara, M.; Williams, P.; Barrett, D.A. Comprehensive profiling of N-acylhomoserine lactones produced by Yersinia pseudotuberculosis using liquid chromatography coupled to hybrid quadrupole-linear ion trap mass spectrometry. Anal. Bioanal. Chem. 2007, 387, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Romero, M.; Prado, S.; Dubert, J.; Tahrioui, A.; Otero, A.; Llamas, I. N-acylhomoserine lactone-degrading bacteria isolated from hatchery bivalve larval cultures. Microbiol. Res. 2013, 168, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.L.; Chalker, V.J.; Kirke, D.; Hardman, A.; Camara, M.; Williams, P. The LuxM Homologue vanM from Vibrio anguillanrum directs the synthesis of N-(3-hydroxyhexanoyl) homoserine lactone and N-hexanoylhomoserine lactone. J. Bacteriol. 2001, 183, 3537–3547. [Google Scholar] [CrossRef] [PubMed]
- Hormigo, D.; De La Mata, I.; Castillon, M.P.; Acebal, C.; Arroyo, M. Kinetic and microstructural characterization of immobilized penicillin acylase from Streptomyces lavendulae on Sepabeads EC-EP. Biocatal. Biotransform. 2009, 27, 271–281. [Google Scholar] [CrossRef]
- Torres, R.; Ramon, F.; de la Mata, I.; Acebal, C.; Castillon, M.P. Enhanced production of penicillin V acylase from Streptomyces lavendulae. Appl. Microbiol. Biotechnol. 1999, 53, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.-S.; Kim, S.-R.; Cheong, W.-S.; Lee, C.-H.; Lee, J.-K. Biofouling inhibition in MBR by Rhodococcus sp. BH4 isolated from real MBR plant. Appl. Microbiol. Biotechnol. 2013, 97, 10223–10231. [Google Scholar] [CrossRef] [PubMed]
- Yates, E.A.; Philipp, B.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y.; Camara, M.; Smith, H.; et al. N-acylhomoserine Lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun. 2002, 70, 5635–5646. [Google Scholar] [CrossRef] [PubMed]
- Mukherji, R.; Varshney, N.K.; Panigrahi, P.; Suresh, C.G.; Prabhune, A. A new role for penicillin acylases: Degradation of acyl homoserine lactone quorum sensing signals by Kluyvera citrophila penicillin G acylase. Enzym. Microb. Technol. 2014, 56, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.; de la Mata, I.; Castillon, M.P.; Arroyo, M.; Torres, J.; Acebal, C. Purification and characterization of penicillin V acylase from Streptomyces lavendulae. In Stability and Stabilization of Biocatalysts; Ballesteros, A., Plou, F.J., Iborra, J.L., Halling, P.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; Volume 15, pp. 719–724. ISBN 0-444-82970-9. [Google Scholar]
- Torres-Bacete, J.; Hormigo, D.; Torres-Guzman, R.; Arroyo, M.; Pilar Castillon, M.; Luis Garcia, J.; Acebal, C.; de la Mata, I. Overexpression of Penicillin V Acylase from Streptomyces lavendulae and elucidation of its catalytic residues. Appl. Environ. Microbiol. 2015, 81, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Cheong, W.-S.; Lee, C.-H.; Moon, Y.-H.; Oh, H.-S.; Kim, S.-R.; Lee, S.H.; Lee, C.-H.; Lee, J.-K. Isolation and identification of indigenous quorum quenching bacteria, Pseudomonas sp. 1A1, for biofouling control in MBR. Ind. Eng. Chem. Res. 2013, 52, 10554–10560. [Google Scholar] [CrossRef]
- Chong, G.; Kimyon, O.; Rice, S.A.; Kjelleberg, S.; Manefield, M. The presence and role of bacterial quorum sensing in activated sludge. Microb. Biotechnol. 2012, 5, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.L.; Park, S.-Y.; Lee, C.-H.; Lee, C.-H.; Lee, J.-K. Quorum quenching bacteria isolated from the sludge of a wastewater treatment plant and their application for controlling biofilm formation. J. Microbiol. Biotechnol. 2014, 24, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Muras, A.; Lopez-Perez, M.; Mayer, C.; Parga, A.; Amaro-Blanco, J.; Otero, A. High prevalence of quorum-sensing and quorum-quenching activity among cultivable bacteria and metagenomic sequences in the Mediterranean Sea. Genes 2018, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-S.; Cao, J.-S.; Li, B.-B.; Li, W.-W.; Fang, F.; Tong, Z.-H.; Yu, H.-Q. Outcompeting presence of Acyl-Homoserine-Lactone (AHL)-quenching bacteria over AHL-producing bacteria in aerobic granules. Environ. Sci. Technol. Lett. 2016, 3, 36–40. [Google Scholar] [CrossRef]
- Feugeas, J.-P.; Tourret, J.; Launay, A.; Bouvet, O.; Hoede, C.; Denamur, E.; Tenaillon, O. Links between transcription, environmental adaptation and gene variability in Escherichia coli: Correlations between gene expression and gene variability reflect growth efficiencies. Mol. Biol. Evol. 2016, 33, 2515–2529. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, S.J.; Oh, T.K.; Oh, J.W.; Koo, B.T.; Yum, D.Y.; Lee, J.K. AhlD, an N-acylhomoserine lactonase in Arthrobacter sp., and predicted homologues in other bacteria. Microbiology-SGM 2003, 149, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; d’Angelo-Picard, C.; Carlier, A.; Elasri, M.; Sicot, C.; Petit, A.; Oger, P.; Faure, D.; Dessaux, Y. Novel bacteria degrading N-acylhomoserine lactones and their use as quenchers of quorum-sensing-regulated functions of plant-pathogenic bacteria. Microbiology-SGM 2003, 149, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Chhabra, S.R.; Camara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology-SGM 2005, 151, 3313–3322. [Google Scholar] [CrossRef] [PubMed]
- Rolinson, G.; Richards, M.; Batchelor, F. Production of 6-Aminopenicillanic Acid. U.S. Patent 3014845, 3014846, 26 December 1961. [Google Scholar]
- Sunder, A.V.; Utari, P.D.; Ramasamy, S.; van Merkerk, R.; Quax, W.; Pundle, A. Penicillin V acylases from Gram-negative bacteria degrade N-acylhomoserine lactones and attenuate virulence in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 2017, 101, 2383–2395. [Google Scholar] [CrossRef] [PubMed]
- Algburi, A.; Zehm, S.; Netrebov, V.; Bren, A.B.; Chistyakov, V.A.; Chikindas, M.L. Subtilosin prevents biofilm formation by inhibiting bacterial quorum sensing. Probiotics Antimicrob. Proteins 2017, 9, 81–90. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | C. violaceum CV026 | A. tumefaciens NT1 | Adherence Capability |
|---|---|---|---|
| Aeromonas hydrophila E2MB52 | Strong | Moderate | Moderate |
| Pseudomonas pseudoalcaligenes E2MB82 | Non-detected | Strong | Strong |
| Thauera butanivorans E2DN83 | Non-detected | Moderate | Strong |
| Pseudomonas aeruginosa E2MBUPVDN12 | Non-detected | Moderate | Strong |
| Pseudomonas aeruginosa E2MBUPVDN20 | Non-detected | Moderate | Strong |
| WWTP | Bacterial Strains | C6-HSL | C8-HSL | C12-HSL |
|---|---|---|---|---|
| WWTPA | Bacillus cereus E1DN3 | + 1 | + | - |
| WWTPA | Bacillus subtilis E1DN6 | - 2 | + | + |
| WWTPA | Bacillus sp. E1DN7 | + | + | + |
| WWTPA | Gordonia paraffinivorans E1N2.12 | - | + | + |
| WWTPA | Bacillus cereus E1DN27 | - | + | + |
| WWTP B | Bacillus cereus E2N2.8 | + | + | + |
| WWTP B | Bacillus thuringiensis E2DN25 | + | + | + |
| WWTP B | Bacillus sp. E2DN30 | - | + | + |
| WWTP B | Bacillus cereus E2DN35 | + | + | + |
| WWTP B | Bacillus cereus E2DN36 | + | + | + |
| WWTP B | Bacillus cereus E2AL40 | + | + | - |
| WWTP B | Bacillus cereus E2AL41 | + | + | + |
| WWTP B | Bacillus thuringiensis E2MB58 | - | + | + |
| WWTP B | Bacillus subtilis E2MB60 | - | + | + |
| WWTP B | Bacillus cereus E2DN70 | + | + | + |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soler, A.; Arregui, L.; Arroyo, M.; Mendoza, J.A.; Muras, A.; Álvarez, C.; García-Vera, C.; Marquina, D.; Santos, A.; Serrano, S. Quorum Sensing versus Quenching Bacterial Isolates Obtained from MBR Plants Treating Leachates from Municipal Solid Waste. Int. J. Environ. Res. Public Health 2018, 15, 1019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15051019
Soler A, Arregui L, Arroyo M, Mendoza JA, Muras A, Álvarez C, García-Vera C, Marquina D, Santos A, Serrano S. Quorum Sensing versus Quenching Bacterial Isolates Obtained from MBR Plants Treating Leachates from Municipal Solid Waste. International Journal of Environmental Research and Public Health. 2018; 15(5):1019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15051019
Chicago/Turabian StyleSoler, Albert, Lucía Arregui, Miguel Arroyo, José Antonio Mendoza, Andrea Muras, Cristina Álvarez, Cristina García-Vera, Domingo Marquina, Antonio Santos, and Susana Serrano. 2018. "Quorum Sensing versus Quenching Bacterial Isolates Obtained from MBR Plants Treating Leachates from Municipal Solid Waste" International Journal of Environmental Research and Public Health 15, no. 5: 1019. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15051019