Effect of Phosphate-Solubilizing Bacteria on the Mobility of Insoluble Cadmium and Metabolic Analysis

Abstract
:1. Introduction
2. Material and Methods
2.1. PSB Isolation and Cd Solubility Tests
2.2. Effects of PSB on Cd Mobility in Contaminated Soil
2.3. Determination of PSB Secretions and Organic Acids
2.4. The Ability of PSB-Secreted Organic Acids to Solubilize Cd
2.5. Statistical Analysis
3. Results and Discussion
3.1. Effects of PSB on Solubilization of Cd Carbonate in Liquid Medium
3.2. Effects of Cd-Mobilizing PSB on Mobility of Soil Cd
3.3. Metabolite Analysis and Their Cd-Solubilizing Abilities
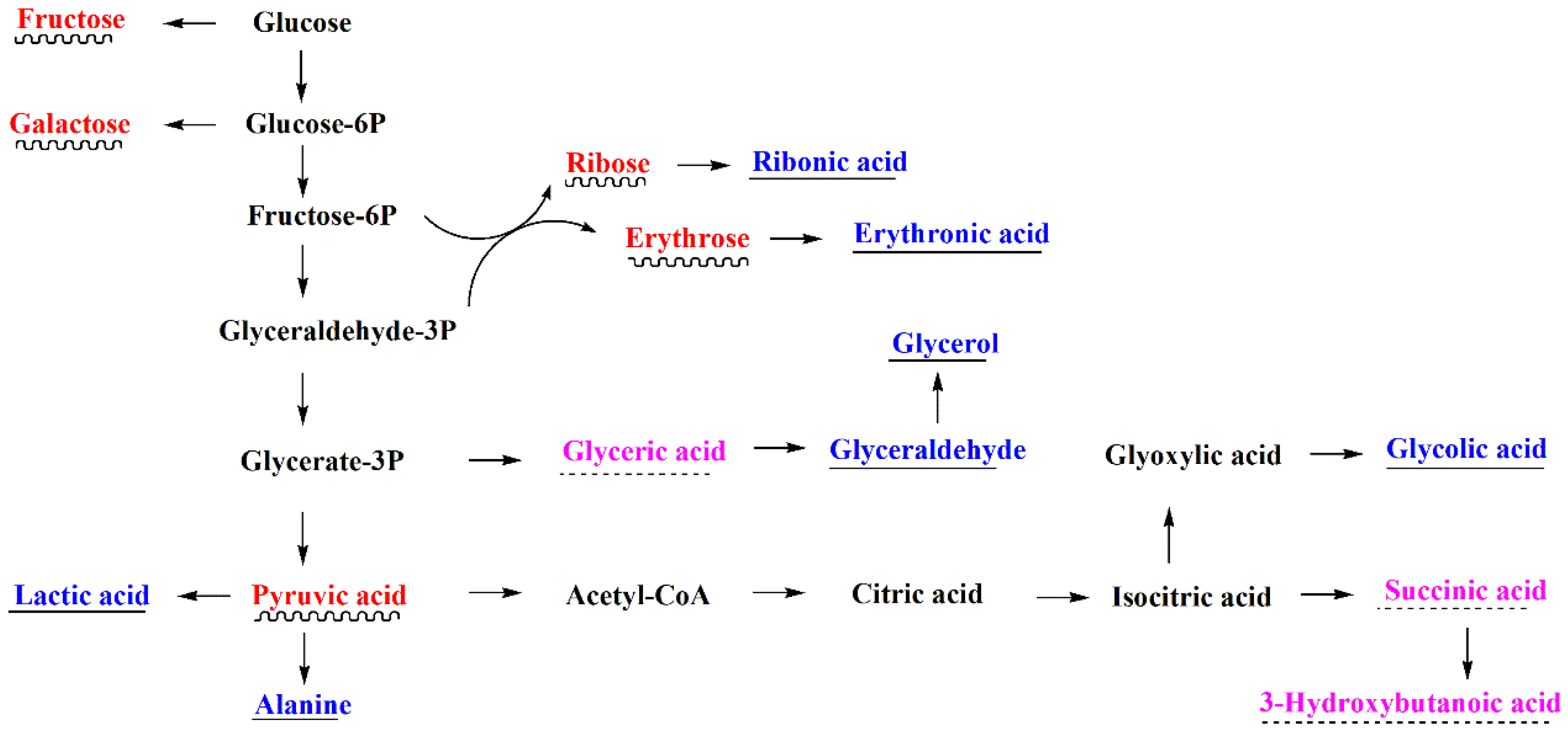
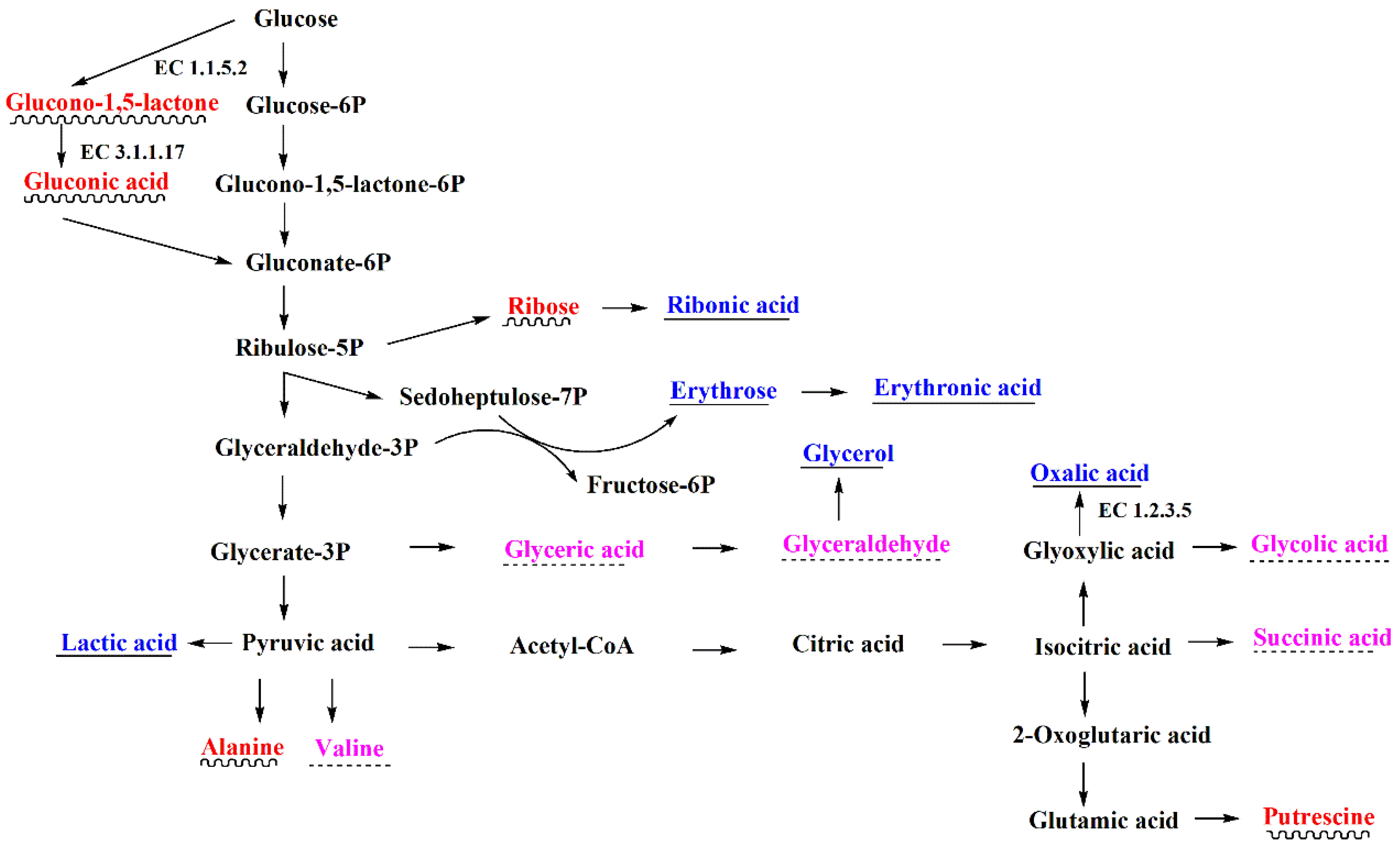
3.4. Metabolic Pathway Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Järup, L.; Åkesson, A. Current status of cadmium as an environmental health problem. Toxicol. Appl. Pharmacol. 2009, 238, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, T.; Plusquin, M.; Hogervorst, J.; Roels, H.A.; Celis, H.; Thijs, L.; Vangronsveld, J.; Van Hecke, E.; Staessen, J.A. Environmental exposure to cadmium and risk of cancer: A prospective population-based study. Lancet Oncol. 2006, 7, 119–126. [Google Scholar] [CrossRef]
- World Health Organization. Exposure to Cadmium: A Major Public Health Concern. Available online: http://www.who.int/ipcs/assessment/public_health/cadmium/en (accessed on 30 May 2018).
- Satarug, S.; Garrett, S.H.; Sens, M.A.; Sens, D.A. Cadmium, environmental exposure, and health outcomes. Environ. Health Perspect. 2010, 118, 182–190. [Google Scholar] [CrossRef] [PubMed]
- He, S.; He, Z.; Yang, X.; Stoffella, P.J.; Baligar, V.C. Chapter 4—Soil biogeochemistry, plant physiology, and phytoremediation of cadmium-contaminated soils. In Advances in Agronomy; Donald, L.S., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 134, pp. 135–225. [Google Scholar]
- Prasad, M.N.V.; Freitas, H.; Fraenzle, S.; Wuenschmann, S.; Markert, B. Knowledge explosion in phytotechnologies for environmental solutions. Environ. Pollut. 2010, 158, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Suresh, B.; Ravishankar, G.A. Phytoremediation—A novel and promising approach for environmental clean-up. Crit. Rev. Biotechnol. 2004, 24, 97–124. [Google Scholar] [CrossRef] [PubMed]
- Surriya, O.; Sarah Saleem, S.; Waqar, K.; Gul Kazi, A. Chapter 1—Phytoremediation of soils: Prospects and challenges. In Soil Remediation and Plants; Hakeem, K.R., Sabir, M., Öztürk, M., Mermut, A.R., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 1–36. [Google Scholar]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—Concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. JKSUS 2014, 26, 1–20. [Google Scholar] [CrossRef]
- Glick, B.R. Using soil bacteria to facilitate phytoremediation. Biotechnol. Adv. 2010, 28, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Prasad, M.N.V.; Rajkumar, M.; Freitas, H. Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol. Adv. 2011, 29, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Ae, N.; Prasad, M.N.V.; Freitas, H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exper. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Ma, Y.; Oliveira, R.S.; Nai, F.; Rajkumar, M.; Luo, Y.; Rocha, I.; Freitas, H. The hyperaccumulator Sedum plumbizincicola harbors metal-resistant endophytic bacteria that improve its phytoextraction capacity in multi-metal contaminated soil. J. Environ. Manag. 2015, 156, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Luo, S.; Li, X.; Wan, Y.; Chen, J.; Liu, C. Interaction of cd-hyperaccumulator Solanum nigrum L. and functional endophyte Pseudomonas sp. Lk9 on soil heavy metals uptake. Soil Biol. Biochem. 2014, 68, 300–308. [Google Scholar] [CrossRef]
- Luo, S.; Xu, T.; Chen, L.; Chen, J.; Rao, C.; Xiao, X.; Wan, Y.; Zeng, G.; Long, F.; Liu, C.; et al. Endophyte-assisted promotion of biomass production and metal-uptake of energy crop sweet sorghum by plant-growth-promoting endophyte Bacillus sp. Sls18. Appl. Microbiol. Biotechnol. 2012, 93, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Archana, G. Cadmium minimization in food crops by cadmium resistant plant growth promoting rhizobacteria. Appl. Soil Ecol. 2016, 107, 66–78. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; Merten, D.; Svatoš, A.; Büchel, G.; Kothe, E. Siderophores mediate reduced and increased uptake of cadmium by Streptomyces tendae f4 and sunflower (Helianthus annuus), respectively. J. Appl. Microbiol. 2009, 107, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- He, L.-Y.; Chen, Z.-J.; Ren, G.-D.; Zhang, Y.-F.; Qian, M.; Sheng, X.-F. Increased cadmium and lead uptake of a cadmium hyperaccumulator tomato by cadmium-resistant bacteria. Ecotoxicol. Environ. Saf. 2009, 72, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.-F.; Xia, J.-J. Improvement of rape (Brassica napus) plant growth and cadmium uptake by cadmium-resistant bacteria. Chemosphere 2006, 64, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Sinha, S.; Mukherjee, S.K. Cadmium–induced siderophore production by a high cd-resistant bacterial strain relieved cd toxicity in plants through root colonization. Curr. Microbiol. 2008, 56, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Lei, M.; Yang, J.; Yang, J.; Wan, X.-M.; Chen, T.-B.; Zhou, X.-Y.; Gu, S.-P.; Guo, G.-H. Effect of fertilizers on the Cd uptake of two sedum species (Sedum spectabile Boreau and Sedum aizoon L.) as potential Cd accumulators. Ecol. Eng. 2017, 106, 409–414. [Google Scholar] [CrossRef]
- Ahemad, M. Phosphate-solubilizing bacteria-assisted phytoremediation of metalliferous soils: A review. 3 Biotech 2015, 5, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Rekha, P.D.; Arun, A.B.; Shen, F.T.; Lai, W.A.; Young, C.C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Gulati, A.; Sharma, N.; Vyas, P.; Sood, S.; Rahi, P.; Pathania, V.; Prasad, R. Organic acid production and plant growth promotion as a function of phosphate solubilization by Acinetobacter rhizosphaerae strain BIHB 723 isolated from the cold deserts of the trans-Himalayas. Arch. Microbiol. 2010, 192, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Stefanoni Rubio, P.J.; Godoy, M.S.; Della Mónica, I.F.; Pettinari, M.J.; Godeas, A.M.; Scervino, J.M. Carbon and nitrogen sources influence tricalcium phosphate solubilization and extracellular phosphatase activity by talaromyces flavus. Curr. Microbiol. 2016, 72, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Tang, S.; Ju, X.; Ding, Y.; Liao, S.; Song, N. Effects of inoculation of a plant growth promoting rhizobacterium Burkholderia sp. D54 on plant growth and metal uptake by a hyperaccumulator Sedum alfredii Hance grown on multiple metal contaminated soil. World J. Microbiol. Biotechnol. 2011, 27, 2835–2844. [Google Scholar] [CrossRef]
- Jeong, S.; Moon, H.S.; Nam, K.; Kim, J.Y.; Kim, T.S. Application of phosphate-solubilizing bacteria for enhancing bioavailability and phytoextraction of cadmium (Cd) from polluted soil. Chemosphere 2012, 88, 204–210. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Ye, Z.; Yang, D.; Yan, J.; Xiao, L.; Zhong, T.; Yuan, M.; Cai, X.; Fang, Z.; Jing, Y. Characterization of endophytic Rahnella sp. Jn6 from Polygonum pubescens and its potential in promoting growth and Cd, Pb, Zn uptake by Brassica napus. Chemosphere 2013, 90, 1960–1965. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.-Y.; Sheng, X.-F.; Qian, M.; Wang, Q.-Y. Isolation and characterization of a heavy metal-resistant Burkholderia sp. From heavy metal-contaminated paddy field soil and its potential in promoting plant growth and heavy metal accumulation in metal-polluted soil. Chemosphere 2008, 72, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 587. [Google Scholar] [CrossRef] [PubMed]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 174. [Google Scholar] [CrossRef] [PubMed]
- Tessier, A.; Campbell, P.G.C.; Bisson, M. Sequential extraction procedure for the speciation of particulate trace metals. Anal. Chem. 1979, 51, 844–851. [Google Scholar] [CrossRef]
- Fiehn, O.; Kopka, J.; Trethewey, R.N.; Willmitzer, L. Identification of Uncommon Plant Metabolites Based on Calculation of Elemental Compositions Using Gas Chromatography and Quadrupole Mass Spectrometry. Anal. Chem. 2000, 72, 3573–3580. [Google Scholar] [CrossRef] [PubMed]
- Ute, R.; Cornelia, W.; Joachim, K.; Richard, N.T.R.; Lothar, W. Simultaneous analysis of metabolites in potato tuber by gas chromatography–mass spectrometry. Plant J. 2000, 23, 131–142. [Google Scholar]
- Halket, J.M.; Przyborowska, A.; Stein, S.E.; Mallard, W.G.; Down, S.; Chalmers, R.A. Deconvolution gas chromatography/mass spectrometry of urinary organic acids—Potential for pattern recognition and automated identification of metabolic disorders. Rapid Commun. Mass Spectrom. 1999, 13, 279–284. [Google Scholar] [CrossRef]
- Stein, S.E. An integrated method for spectrum extraction and compound identification from gas chromatography/mass spectrometry data. J. Am. Soc. Mass Spectrom. 1999, 10, 770–781. [Google Scholar] [CrossRef]
- Katsumasa, S.; Keiki, O.; Keitaro, T.; Mitsuru, O.; Takuro, S. Gas chromatography–mass spectrometry associated global analysis of rice root exudates under aseptical conditions. Soil Sci. Plant Nutr. 2009, 55, 505–513. [Google Scholar] [Green Version]
- Okazaki, K.; Oka, N.; Shinano, T.; Osaki, M.; Takebe, M. Differences in the Metabolite Profiles of Spinach (Spinacia oleracea L.) Leaf in Different Concentrations of Nitrate in the Culture Solution. Plant and Cell Physiol. 2008, 49, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Erban, A.; Schauer, N.; Fernie, A.R.; Kopka, J. Nonsupervised Construction and Application of Mass Spectral and Retention Time Index Libraries from Time-of-Flight Gas Chromatography-Mass Spectrometry Metabolite Profiles. In Metabolomics: Methods and Protocols; Weckwerth, W., Ed.; Humana Press: Totowa, NJ, USA, 2007; Volume 358, pp. 19–38. [Google Scholar]
- Kumari, A.; Parida, A.K. Metabolomics and network analysis reveal the potential metabolites and biological pathways involved in salinity tolerance of the halophyte Salvadora persica. Environ. Exper. Bot. 2018, 148, 85–99. [Google Scholar] [CrossRef]
- Li, K.; Ramakrishna, W. Effect of multiple metal resistant bacteria from contaminated lake sediments on metal accumulation and plant growth. J. Hazard. Mater. 2011, 189, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Rajkumar, M.; Freitas, H. Isolation and characterization of Ni mobilizing PGPB from serpentine soils and their potential in promoting plant growth and Ni accumulation by Brassica spp. Chemosphere 2009, 75, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Rajkumar, M.; Freitas, H. Improvement of plant growth and nickel uptake by nickel resistant-plant-growth promoting bacteria. J. Hazard. Mater. 2009, 166, 1154–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajkumar, M.; Freitas, H. Influence of metal resistant-plant growth-promoting bacteria on the growth of ricinus communis in soil contaminated with heavy metals. Chemosphere 2008, 71, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.; Ma, Y.; Freitas, H. Characterization of metal-resistant plant-growth promoting Bacillus weihenstephanensis isolated from serpentine soil in Portugal. J. Basic Microbiol. 2008, 48, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Buch, A.; Archana, G.; Naresh Kumar, G. Metabolic channeling of glucose towards gluconate in phosphate-solubilizing Pseudomonas aeruginosa p4 under phosphorus deficiency. Res. Microbiol. 2008, 159, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Fuhrer, T.; Fischer, E.; Sauer, U. Experimental identification and quantification of glucose metabolism in seven bacterial species. J. Bacteriol. 2005, 187, 1581–1590. [Google Scholar] [CrossRef] [PubMed]
- KEGG PATHWAY Database. Available online: http://www.genome.jp/kegg/pathway.html (accessed on 30 May 2018).
- Buch, A.; Archana, G.; Naresh Kumar, G. Heterologous expression of phosphoenolpyruvate carboxylase enhances the phosphate solubilizing ability of fluorescent pseudomonads by altering the glucose catabolism to improve biomass yield. Bioresour. Technol. 2010, 101, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Nikel, P.I.; Chavarría, M.; Fuhrer, T.; Sauer, U.; de Lorenzo, V. Pseudomonas putida kt2440 Strain Metabolizes Glucose through a Cycle Formed by Enzymes of the Entner-Doudoroff, Embden-Meyerhof-Parnas, and Pentose Phosphate Pathways. J. Biol. Chem. 2015, 290, 25920–25932. [Google Scholar] [CrossRef] [PubMed]
- Buch, A.D.; Archana, G.; Kumar, G.N. Enhanced citric acid biosynthesis in Pseudomonas fluorescens ATCC 13525 by overexpression of the Escherichia coli citrate synthase gene. Microbiology 2009, 155, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Ludueña, L.M.; Anzuay, M.S.; Magallanes-Noguera, C.; Tonelli, M.L.; Ibañez, F.J.; Angelini, J.G.; Fabra, A.; McIntosh, M.; Taurian, T. Effects of P limitation and molecules from peanut root exudates on pqqE gene expression and pqq promoter activity in the phosphate-solubilizing strain Serratia sp. S119. Res. Microbiol. 2017, 168, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, G.; Ishikawa, Y.; Kobayashi, K.; Migita, C.T.; Elias, M.D.; Nakamura, S.; Tagawa, S.; Yamada, M. Amino acid residues interacting with both the bound quinone and coenzyme, pyrroloquinoline quinone, in Escherichia coli membrane-bound glucose dehydrogenase. J. Biol. Chem. 2008, 283, 22215–22221. [Google Scholar] [CrossRef] [PubMed]
- Rodrı́guez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | 1 Day | 7 Days | ||||||
|---|---|---|---|---|---|---|---|---|
| PO43– (mg/L) | Cd2+ (mg/L) | Cd2+ Dissolution Rate (%) | pH | PO43– (mg/L) | Cd2+ (mg/L) | Cd2+ Dissolution Rate (%) | pH | |
| B. cereus1 | 6.16 ± 0.17 bA | 16.94 ± 0.80 bA | 43.32 | 6.24 ± 0.05 bB | 10.06 ± 0.90 bB | 20.03 ± 1.42 dB | 51.23 | 5.38 ± 0.24 bA |
| P. fluorescens1 | 31.82 ± 2.13 cA | 29.96 ± 2.84 cB | 76.62 | 5.00 ± 0.11 aA | 30.55 ± 1.34 cA | 12.00 ± 0.77 cA | 30.69 | 6.36 ± 0.05 cB |
| B. cereus2 | 4.31 ± 0.19 abA | 4.40 ± 0.15 aA | 11.26 | 6.49 ± 0.03 cA | 4.04 ± 0.20 aA | 5.34 ± 0.11 bB | 13.66 | 6.43 ± 0.06 cA |
| P. fluorescens2 | 4.66 ± 0.23 abA | 5.18 ± 0.04 aA | 13.26 | 6.28 ± 0.03 bB | 64.27 ± 2.45 dB | 35.98 ± 0.56 eB | 92.03 | 4.09 ± 0.33 aA |
| control | 3.69 ± 0.30 aA | 2.87 ± 0.24 aA | 7.34 | 6.71 ± 0.04 dB | 3.76 ± 0.32 aA | 3.68 ± 0.35 aB | 9.41 | 6.54 ± 0.04 cA |
| Treatment | 1 Day | 7 Days | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Dissolved Cd2+ (mg/kg Dry Soil) | pH | Net Dissolved Cd2+ (mg/kg Dry Soil) | Ratio of Net Dissolved Cd2+ to Total Cadmium in Soil | Ratio of Net Dissolved Cd2+ to the Exchangeable and Carbonate-Bound Cadmium | Dissolved Cd2+ (mg/kg Dry Soil) | pH | Net Dissolved Cd2+ (mg/kg Dry Soil) | Ratio of Net Dissolved Cd2+ to Total Cadmium in Soil | Ratio of Net Dissolved Cd2+ to the Exchangeable and Carbonate-Bound Cadmium | |
| B. cereus | 0.08 ± 0.03 a | 6.55 ± 0.07 b | 0.04 | 2.0% | 7.6% | 0.03 ± 0.01 a | 6.66 ± 0.09 b | - | - | - |
| P. fluorescens | 0.22 ± 0.03 b | 4.42 ± 0.05 a | 0.18 | 9.2% | 34.3% | 0.43 ± 0.08 b | 4.46 ± 0.18 a | 0.39 | 20.0% | 74.3% |
| control | 0.04 ± 0.02 a | 6.57 ± 0.20 b | 0.04 ± 0.02 a | 6.48 ± 0.20 b | ||||||
| Organic acid | Concentration in B. cereus (mg/L) | Cd-Solubilizing Ability in B. cereus (mg/L) | Concentration in P. fluorescens (mg/L) | Cd-Solubilizing Ability in P. fluorescens (mg/L) |
|---|---|---|---|---|
| Pyruvic acid | 9.738 | 4.52 | n. d. | n. d. |
| Lactic acid | 1.988 | 2.11 | 1.193 | 1.80 |
| Glycolic acid | 5.382 | 2.95 | 0.339 | - |
| Oxalic acid | n. d. | n. d. | 6.962 | 7.52 |
| 3-Hydroxy butyric acid | 0.783 | - | n. d. | n. d. |
| Succinic acid | 1.098 | 2.28 | 0.854 | - |
| Glyceric acid | 0.998 | - | 0.431 | - |
| Gluconic acid | n. d. | n. d. | 75.315 | 13.08 |
| Erythronic acid | 2.120 | 1.51 | 5.260 | 2.22 |
| Ribonic acid | 3.242 | 1.77 | 3.554 | 1.84 |
| 3-Hydroxy propionic acid | 0.087 | - | n. d. | n. d. |
| 2,4-Dihydroxy butanoic acid | 1.346 | 1.34 | 2.203 | 1.53 |
| 3,4-Dihydroxy butanoic acid | 1.255 | 1.32 | 2.084 | 1.57 |
| Hexadecanoic acid | 1.466 | 1.38 | 1.079 | 1.26 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Zhou, X.-F.; Wang, L.-L.; Li, Q.-S.; Zhou, T.; Chen, Y.-K.; Zhao, Z.-Y.; He, B.-Y. Effect of Phosphate-Solubilizing Bacteria on the Mobility of Insoluble Cadmium and Metabolic Analysis. Int. J. Environ. Res. Public Health 2018, 15, 1330. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15071330
Yang P, Zhou X-F, Wang L-L, Li Q-S, Zhou T, Chen Y-K, Zhao Z-Y, He B-Y. Effect of Phosphate-Solubilizing Bacteria on the Mobility of Insoluble Cadmium and Metabolic Analysis. International Journal of Environmental Research and Public Health. 2018; 15(7):1330. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15071330
Chicago/Turabian StyleYang, Ping, Xue-Fang Zhou, Li-Li Wang, Qu-Sheng Li, Ting Zhou, Yu-Kun Chen, Zi-Yi Zhao, and Bao-Yan He. 2018. "Effect of Phosphate-Solubilizing Bacteria on the Mobility of Insoluble Cadmium and Metabolic Analysis" International Journal of Environmental Research and Public Health 15, no. 7: 1330. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph15071330