Preharvest Transmission Routes of Fresh Produce Associated Bacterial Pathogens with Outbreak Potentials: A Review


Abstract
:1. Introduction
2. Global Burden of Fresh Produce-Related Foodborne Diseases
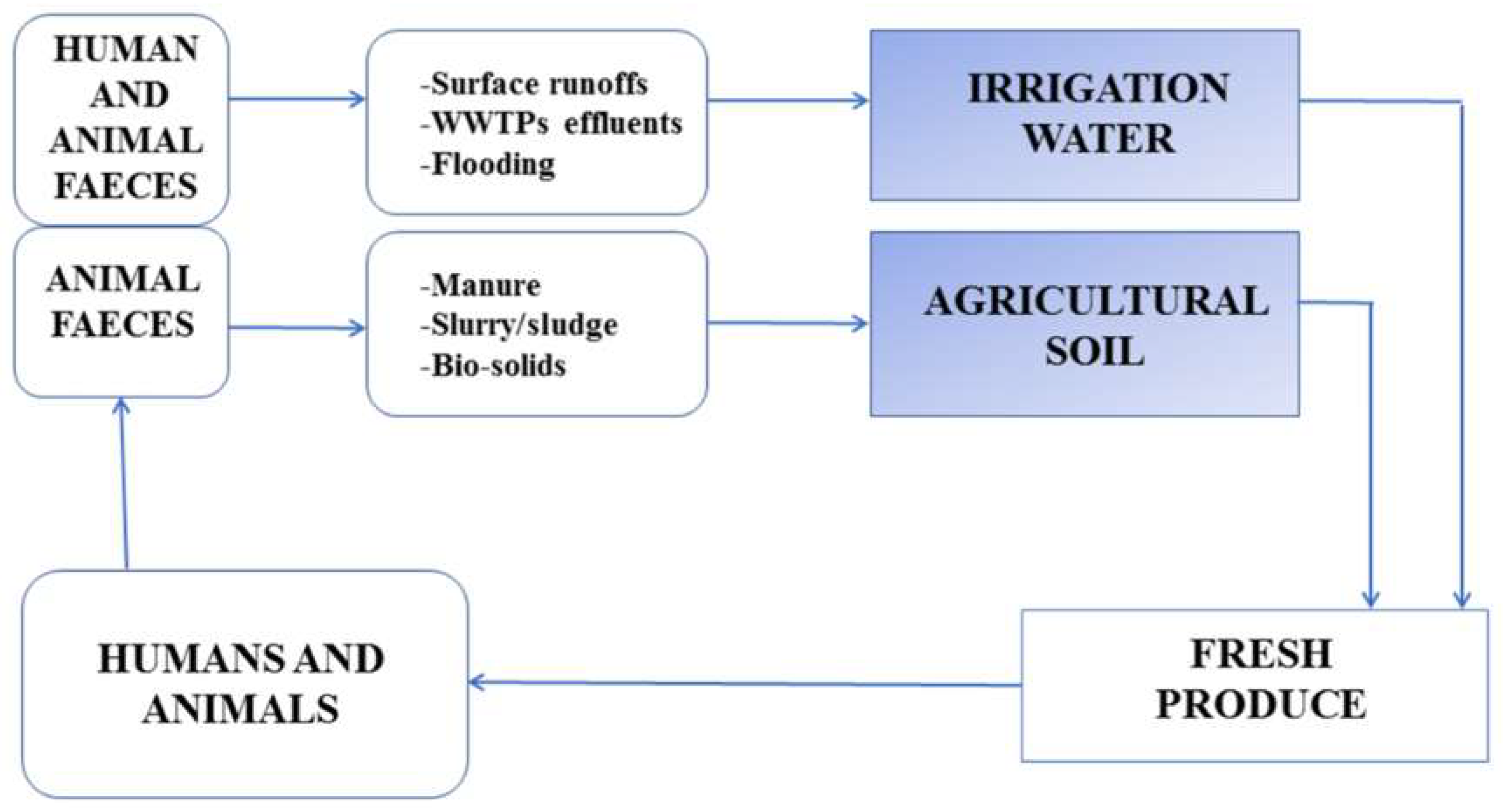
3. Fresh Produce Contamination between the Continuum of Farm to Table
4. Preharvest Transmission Routes of Fresh Produce Associated Foodborne Pathogens
4.1. Irrigation Water
4.1.1. Sources of Irrigation Water
Wastewater
Surface Water
Groundwater
Harvested Rainwater
4.1.2. The Microbiology of Irrigation Water
4.1.3. The Effect of Irrigation Application Methods and Timing on the Microbial Contamination of Fresh Produce
4.2. Agricultural Soil
4.3. Manure
4.4. Animal Intrusion
5. Some Bacterial Pathogens with Outbreak Potentials and Their Antimicrobial Resistance Trends
5.1. Escherichia coli
E. coli O157: H7
5.2. Salmonella spp.
5.3. Shigella spp.
5.4. Klebsiella spp.
5.5. Citrobacter spp.
5.6. Enterobacter spp.
5.7. ESBL-Producing Enterobacteriaceae
5.8. Listeria Monocytogenes
6. The Impact of Antibiotic Resistance
7. Interventions to Prevent Fresh Produce Contamination Prior to Harvesting
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Garedew, L.; Hagos, Z.; Zegeye, B.; Addis, Z. The detection and antimicrobial susceptibility profile of Shigella isolates from meat and swab samples at butchers’ shops in Gondar town, Northwest Ethiopia. J. Infect. Public Health 2016, 9, 348–355. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization, Geneva, Switzerland. Food Safety and Food Borne Illness. WHO Media Center Fact Sheet, Food Safety Department. Available online: https://foodhygiene2010.files.wordpress.com/2010/06/who-food_safety_fact-sheet.pdf (accessed on 6 October 2018).
- Yeni, F.; Yavaş, S.; Alpas, H.; Soyer, Y. Most Common Foodborne Pathogens and Mycotoxins on Fresh Produce: A Review of Recent Outbreaks. Crit. Rev. Food Sci. Nutr. 2016, 56, 1532–1544. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Jang, H.; Matthews, K.R. Effect of the food production chain from farm practices to vegetable processing on outbreak incidence. Microb. Biotechnol. 2014, 7, 517–527. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority); ECDC (European Centre for Disease Prevention and Control). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 3429. [Google Scholar] [CrossRef]
- CDC Centers for Disease Control and Prevention. Surveillance for Foodborne Disease Outbreaks, United States. Annual Report: US Department of Health and Human Services, Atlanta, Georgia. 2012. Available online: https://www.cdc.gov/foodsafety/pdfs/foodborne-disease-outbreaks-annual-report-2012-508c.pdf (accessed on 6 October 2018).
- Beuchat, L.R. Ecological factors influencing survival and growth of human pathogens on raw fruits and vegetables. Microbes Infect. 2002, 4, 413–423. [Google Scholar] [CrossRef]
- Olaimat, A.N.; Holley, R.A. Factors influencing the microbial safety of fresh produce: A review. Food Microbiol. 2012, 32, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.N.; Sodha, S.V.; Shaw, R.K.; Griffin, P.M.; Pink, D.; Hand, P.; Frankel, G. Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol. 2010, 12, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Navratil, S.; Gregory, A.; Bauer, A.; Srinath, I.; Szonyi, B.; Nightingale, K.; Anciso, J.; Jun, M.; Han, D.; et al. Multifactorial Effects of Ambient Temperature, Precipitation, Farm Management, and Environmental Factors Determine the Level of Generic Escherichia coli Contamination on Preharvested Spinach. Appl. Environ. Microbiol. 2015, 81, 2635–2650. [Google Scholar] [CrossRef] [PubMed]
- Marine, S.C.; Pagadala, S.; Wang, F.; Pahl, D.M.; Melendez, M.V.; Kline, W.L.; Oni, R.A.; Walsh, C.S.; Everts, K.L.; Buchanan, R.L.; et al. The growing season, but not the farming system, is a food safety risk determinant for leafy greens in the mid-Atlantic region of the United States. Appl. Environ. Microbiol. 2015, 81, 2395–2407. [Google Scholar] [CrossRef] [PubMed]
- Rajwar, A.; Srivastava, P.; Sahgal, M. Microbiology of Fresh Produce: Route of Contamination, Detection Methods, and Remedy. Crit. Rev. Food Sci. Nutr. 2016, 56, 2383–2390. [Google Scholar] [CrossRef] [PubMed]
- Uyttendaele, M.; Jaykus, L.A.; Amoah, P.; Chiodini, A.; Cunliffe, D.; Jacxsens, L.; Holvoet, K.; Korsten, L.; Lau, M.; McClure, P.; et al. Microbial Hazards in Irrigation Water: Standards, Norms, and Testing to Manage Use of Water in Fresh Produce Primary Production. Compr. Rev. Food Sci. Food Saf. 2015, 14, 336–356. [Google Scholar] [CrossRef]
- Faour-klingbeil, D.; Murtada, M.; Kuri, V.; Todd, E.C.D. Understanding the routes of contamination of ready-to-eat vegetables in the Middle East. Food Control 2016, 62, 125–133. [Google Scholar] [CrossRef]
- Jongman, M.; Korsten, L. Irrigation water quality and microbial safety of leafy greens in different vegetable production systems: A review. Food Rev. Int. 2018, 34, 308–328. [Google Scholar] [CrossRef]
- Callejón, R.M.; Rodríguez-naranjo, I.M.; Ubeda, C.; Hornedo-ortega, R.; Garcia-parrilla, C.M.; Troncoso, A.M. Reported Foodborne Outbreaks Due to Fresh Produce in the United States and European Union: Trends and Causes. Foodborne Pathog. Dis. 2015, 12, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Kozak, G.K.; Macdonald, D.; Landry, L.; Farber, J.M. Foodborne Outbreaks in Canada Linked to Produce: 2001 through 2009. J. Food Prot. 2013, 76, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Barrett, D.M.; Beaulieu, J.C.; Shewfelt, R.; Barrett, D.M.; Beaulieu, J.C.; Color, R.S.; Barrett, D.M.; Beaulieu, J.C.; Shewfelt, R.O.B. Color, Flavor, Texture, and Nutritional Quality of Fresh-Cut Fruits and Vegetables: Desirable Levels, Instrumental and Sensory Measurement, and the Effects of Processing Nutritional Quality of Fresh-Cut Fruits and Vegetables: Desirable Levels, Instrumenta. Crit. Rev. Food Sci. Nutr. 2010, 50, 369–389. [Google Scholar] [CrossRef] [PubMed]
- Dietary Guidelines for Americans 2015–2020 U.S. Department of Health and Human Services and U.S. Department of Agriculture. 2015–2020. Available online: https://health.gov/dietaryguidelines/2015/ (accessed on 11 October 2018).
- Food-Based Dietary Guidelines in Europe. Scientific Opinion on establishing Food-Based Dietary Guidelines. EFSA J. 2010, 8, 1460. [Google Scholar] [CrossRef]
- Castro-Ibáñez, I.; Gil, M.I.; Allende, A. Ready-to-eat vegetables: Current problems and potential solutions to reduce microbial risk in the production chain. LWT Food Sci. Technol. 2017, 85, 284–292. [Google Scholar] [CrossRef]
- FAO/WHO—Food and Agricultural Organization of the United Nations/World Health Organization. Microbiological Hazards in Fresh Fruits and Vegetables. Available online: http://www.who.int/foodsafety/publications/micro/MRA_FruitVeges.pdf (accessed on 13 October 2018).
- Denis, N.; Zhang, H.; Leroux, A.; Trudel, R.; Bietlot, H. Prevalence and trends of bacterial contamination in fresh fruits and vegetables sold at retail in Canada. Food Control 2016, 67, 225–234. [Google Scholar] [CrossRef]
- CDC Centers for Disease Control and Prevention. List of Selected Multistate Foodborne Outbreak Investigations. Available online: https://www.cdc.gov/foodsafety/outbreaks/multistate-outbreaks/outbreaks-list.html (accessed on 25 March 2019).
- Grant, J.; Wendelboe, A.M.; Wendel, A.; Jepson, B.; Torres, P.; Smelser, C.; Rolfs, R.T. Spinach-associated Escherichia coli O157:H7 outbreak, Utah and New Mexico, 2006. Emerg. Infect. Dis. 2008, 14, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Wendel, A.M.; Hoang Johnson, D.; Sharapov, U.; Grant, J.; Archer, J.R.; Monson, T.; Koschmann, C.; Davis, J.P. Multistate Outbreak of Escherichia coli O157:H7 Infection Associated with Consumption of Packaged Spinach, August–September 2006: The Wisconsin Investigation. Clin. Infect. Dis. 2009, 48, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Hanning, I.B.; Nutt, J.D.; Ricke, S.C. Salmonellosis Outbreaks in the United States Due to Fresh Produce: Sources and Potential Intervention Measures. Foodborne Pathog. Dis. 2009, 6, 635–648. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. List of Selected Multistate Foodborne Outbreak Investigations Foodborne Outbreaks Food Safety CDC. Available online: https://www.cdc.gov/foodsafety/outbreaks/multistate-outbreaks/outbreaks-list.html (accessed on 13 October 2018).
- EFSA European Food Safety Authority. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15. [Google Scholar] [CrossRef]
- Byrne, L.; Fisher, I.; Peters, T.; Mather, A.; Thomson, N.; Rosner, B.; Bernard, H.; McKeown, P.; Cormican, M.; Cowden, J.; et al. International Outbreak Control Team a multi-country outbreak of Salmonella Newport gastroenteritis in Europe associated with watermelon from Brazil, confirmed by whole genome sequencing: October 2011 to January 2012. Euro Surveill. 2014, 19, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Tauxe, R.; Hedberg, C. The growing burden of foodborne outbreaks due to contaminated fresh produce: Risks and opportunities. Epidemiol. Infect. 2009, 137, 307. [Google Scholar] [CrossRef] [PubMed]
- Elviss, N.C.; Little, C.L.; Hucklesby, L.; Sagoo, S.; Surman-Lee, S.; de Pinna, E.; Threlfall, E.J. Food, Water and Environmental Surveillance Network. Microbiological study of fresh herbs from retail premises uncovers an international outbreak of salmonellosis. Int. J. Food Microbiol. 2009, 134, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Pezzoli, L.; Elson, R.; Little, C.L.; Yip, H.; Fisher, I.; Yishai, R.; Anis, E.; Valinsky, L.; Biggerstaff, M.; Patel, N.; et al. Packed with Salmonella—Investigation of an International Outbreak of Salmonella Senftenberg Infection Linked to Contamination of Prepacked Basil in 2007. Foodborne Pathog. Dis. 2008, 5, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Sewell, A.M.; Farber, J.M. Foodborne outbreaks in Canada linked to produce. J. Food Prot. 2001, 64, 1863–1877. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Sarker, M.R.; Hossain, A. Microbiological food safety: A dilemma of developing societies. Crit. Rev. Microbiol. 2014, 40, 348–359. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. Outbreak of Salmonella Infections Linked to Cavi Brand Whole, Fresh Papayas. Available online: https://www.cdc.gov/salmonella/uganda-06-19/index.html (accessed on 16 July 2019).
- CDC Centers for Disease Control and Prevention. Outbreak of Salmonella Infections Linked to Pre-Cut Melons. Available online: https://www.cdc.gov/salmonella/carrau-04-19/index.html (accessed on 24 April 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of E. coli O157:H7 Infections Linked to Romaine Lettuce (Final Update). Available online: https://www.cdc.gov/ecoli/2018/o157h7-04-18/index.html (accessed on 25 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Shiga Toxin-Producing Escherichia coli O157:H7 Infections Linked to Leafy Greens (Final Update). Available online: https://www.cdc.gov/ecoli/2017/o157h7-12-17/index.html (accessed on 25 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Adelaide Infections Linked to Pre-Cut Melon (Final Update). Available online: https://www.cdc.gov/salmonella/adelaide-06-18/index.html (accessed on 25 March 2019).
- Wadamori, Y.; Gooneratne, R.; Hussain, M.A. Outbreaks and factors influencing microbiological contamination of fresh produce. J. Sci. Food Agric. 2017, 97, 1396–1403. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Shiga Toxin-Producing Escherichia coli O157 Infections Linked to Alfalfa Sprouts Produced by Jack & The Green Sprouts (Final Update). Available online: https://www.cdc.gov/ecoli/2016/o157-02-16/index.html (accessed on 25 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Frozen Vegetables (Final Update). Available online: https://www.cdc.gov/listeria/outbreaks/frozen-vegetables-05-16/index.html (accessed on 25 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Infections Linked to Alfalfa Sprouts from One Contaminated Seed Lot (Final Update). Available online: https://www.cdc.gov/salmonella/muenchen-02-16/index.html (accessed on 25 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Packaged Salads Produced at Springfield, Ohio Dole Processing Facility (Final Update). Available online: https://www.cdc.gov/listeria/outbreaks/bagged-salads-01-16/index.html (accessed on 25 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Poona Infections Linked to Imported Cucumbers (Final Update). Available online: https://www.cdc.gov/salmonella/poona-09-15/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Commercially Produced, Prepackaged Caramel Apples Made from Bidart Bros. Apples (Final Update). Available online: https://www.cdc.gov/listeria/outbreaks/caramel-apples-12-14/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Enteritidis Infections Linked to Bean Sprouts (Final Update). Available online: https://www.cdc.gov/salmonella/enteritidis-11-14/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Shiga Toxin-Producing Escherichia coli O121 Infections Linked to Raw Clover Sprouts (Final Update). Available online: https://www.cdc.gov/ecoli/2014/O121-05-14/index.html (accessed on 26 March 2019).
- PHE Public Health England. Foodborne Illness in Humans: General Outbreaks in England and Wales in 2014. Available online: https://www.gov.uk/government/publications/foodborne-illness-in-humans-general-outbreaks-in-england-and-wales-in-2014 (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Shiga Toxin-Producing Escherichia coli O157:H7 Infections Linked to Ready-to-Eat Salads (Final Update). Available online: https://www.cdc.gov/ecoli/2013/O157H7-11-13/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Saintpaul Infections Linked to Imported Cucumbers (Final Update). Available online: https://www.cdc.gov/salmonella/saintpaul-04-13/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Shiga Toxin-Producing Escherichia coli O157:H7 Infections Linked to Organic Spinach and Spring Mix Blend (Final Update). Available online: https://www.cdc.gov/ecoli/2012/O157H7-11-12/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Braenderup Infections Associated with Mangoes (Final Update). Available online: https://www.cdc.gov/salmonella/braenderup-08-12/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Typhimurium and Salmonella Newport Infections Linked to Cantaloupe (Final Update). Available online: https://www.cdc.gov/salmonella/typhimurium-cantaloupe-08-12/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Shiga Toxin-Producing Escherichia coli O26 Infections Linked to Raw Clover Sprouts at Jimmy John’s Restaurants (Final Update). Available online: https://www.cdc.gov/ecoli/2012/O26-02-12/index.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of E. coli O157:H7 Infections Linked to Romaine Lettuce (Final Update). Available online: https://www.cdc.gov/ecoli/2011/romaine-lettace-3-23-12.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Listeriosis Linked to Whole Cantaloupes from Jensen Farms, Colorado (Final Update). Available online: https://www.cdc.gov/listeria/outbreaks/cantaloupes-jensen-farms/index.html (accessed on 12 March 2018).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Human Salmonella Agona Infections Linked to Whole, Fresh Imported Papayas (Final Update). Available online: https://www.cdc.gov/salmonella/2011/papayas-8-29-2011.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Human Salmonella Enteritidis Infections Linked to Alfalfa Sprouts and Spicy Sprouts (Final Update). Available online: https://www.cdc.gov/salmonella/2011/alfalfa-spicy-sprouts-7-6-2011.html (accessed on 26 March 2019).
- ECDC European Centre for Disease Prevention and Control. Outbreak of Shiga Toxin-Producing E. coli (STEC) in Germany, May 2011. Available online: https://ecdc.europa.eu/en/publications-data/outbreak-shiga-toxin-producing-e-coli-stec-germany-may-2011 (accessed on 26 March 2019).
- EFSA European Food Safety Authority. Shiga Toxin-Producing E. coli (STEC) O104:H4 2011 Outbreaks in Europe: Taking Stock. EFSA J. 2011, 9, 2390. [Google Scholar] [CrossRef]
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Human Salmonella I 4,[5],12:i: -Infections Linked to Alfalfa Sprouts (Final Update). Available online: https://www.cdc.gov/salmonella/2010/alfalfa-sprouts-2-10-11.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Human Salmonella Newport Infections Linked to Raw Alfalfa Sprouts (Final Update). Available online: https://www.cdc.gov/salmonella/2010/newport-alfalfa-sprout-6-29-10.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Human E. coli O145 Infections Linked to Shredded Romaine Lettuce from a Single Processing Facility (Final Update). Available online: https://www.cdc.gov/ecoli/2010/shredded-romaine-5-21-10.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Saintpaul Infections Linked to Raw Alfalfa Sprouts (Final Update). Available online: https://www.cdc.gov/salmonella/2009/raw-alfalfa-sprouts-5-8-2009.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Saintpaul Infections Linked to Raw Produce (Final Update). Available online: http://www.nejm.org/doi/abs/10.1056/NEJMoa1005741 (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Outbreak of Salmonella Serotype Saintpaul Infections Associated with Multiple Raw Produce Items-United States. 2008. Available online: http://www.ncbi.nlm.nih.gov/pubmed/18756191 (accessed on 26 March 2019).
- Mody, R.K.; Greene, S.A.; Gaul, L.; Sever, A.; Pichette, S.; Zambrana, I.; Dang, T.; Gass, A.; Wood, R.; Herman, K.; et al. National outbreak of Salmonella serotype saintpaul infections: Importance of Texas restaurant investigations in implicating jalapeño peppers. PLoS ONE 2011, 6, e16579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Litchfield Infections Linked to Cantaloupes (Final Update). Available online: https://www.cdc.gov/salmonella/2008/cantaloupes-4-2-2008.html (accessed on 26 March 2019).
- Warriner, K.; Namvar, A. The tricks learnt by human enteric pathogens from phytopathogens to persist within the plant environment. Curr. Opin. Biotechnol. 2010, 21, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Denny, J.; Threlfall, J.; Takkinen, J.; Löfdahl, S.; Westrell, T.; Varela, C.; Adak, B.; Boxall, N.; Ethelberg, S.; Torpdahl, M.; et al. Multinational Salmonella Paratyphi B variant Java (Salmonella Java) outbreak, August–December 2007. Euro Surveill. 2007, 12, 3332. [Google Scholar] [CrossRef] [PubMed]
- Pezzoli, L.; Elson, R.; Little, C.; Fisher, I.; Yip, H.; Peters, T.M.; Hampton, M.; de Pinna, E.; Coia, J.E.; Mather, H.; et al. International outbreak of Salmonella Senftenberg in 2007. Euro Surveill. 2007, 12, 3218. [Google Scholar] [CrossRef] [PubMed]
- Lewis, H.C.; Kirk, M.; Ethelberg, S.; Stafford, R.; Olsen, K.E.; Nielsen, E.M.; Lisby, M.; Madsen, S.B.; Mølbak, K. Outbreaks of shigellosis in Denmark and Australia associated with imported baby corn, August 2007—Final summary. Euro Surveill. 2007, 12, 3279. [Google Scholar] [CrossRef] [PubMed]
- Emberland, K.E.; Ethelberg, S.; Kuusi, M.; Vold, L.; Jensvoll, L.; Lindstedt, B.A.; Nygård, K.; Kjelsø, C.; Torpdahl, M.; Sørensen, G.; et al. Outbreak of Salmonella Weltevreden infections in Norway, Denmark and Finland associated with alfalfa sprouts, July–October 2007. Euro Surveill. 2007, 12, 3321. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of Salmonella Typhimurium Infections Linked to Tomatoes (Final Update). Available online: https://www.cdc.gov/salmonella/2006/tomatoes-11-2006.html (accessed on 26 March 2019).
- CDC Centers for Disease Control and Prevention. Multistate Outbreak of E. coli O157:H7 Infections Linked to Fresh Spinach (Final Update). Available online: https://www.cdc.gov/ecoli/2006/spinach-10-2006.html (accessed on 26 March 2019).
- Compton, J.A.; Baney, J.A.; Donaldson, S.C.; Houser, B.A.; San Julian, G.J.; Yahner, R.H.; Chmielecki, W.; Reynolds, S.; Jayarao, B.M. Salmonella infections in the common raccoon (Procyon lotor) in Western Pennsylvania. J. Clin. Microbiol. 2008, 46, 3084–3086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munnoch, S.A.; Ward, K.; Sheridan, S.; Fitzsimmons, G.J.; Shadbolt, C.T.; Piispanen, J.P.; Wang, Q.; Ward, T.J.; Worgan, T.L.; Oxenford, C.; et al. A multi-state outbreak of Salmonella Saintpaul in Australia associated with cantaloupe consumption. Epidemiol. Infect. 2009, 137, 367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohekar, S.; Tsui, F.W.; Tsui, H.W.; Xi, N.; Riarh, R.; Bilotta, R.; Inman, R.D. Symptomatic Acute Reactive Arthritis After An Outbreak of Salmonella. J. Rheumatol. 2008, 35, 1599–1602. [Google Scholar] [PubMed]
- CDC Centers for Disease Control and Prevention. Multistate Outbreaks of Salmonella Infections Associated with Raw Tomatoes Eaten in Restaurants—United States. 2005–2006. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17805221 (accessed on 26 March 2019).
- Datex Food Safety from Farm to Fork. Available online: https://www.slideshare.net/acoxdatex/food-safety-from-farm-to-fork (accessed on 18 October 2018).
- Microbiological Hazards in Fresh Leafy Vegetables and Herbs; Italy, 2008. Available online: https://apps.who.int/iris/handle/10665/44031 (accessed on 8 November 2019).
- USDA U.S. Department of Agriculture. USDA Good Agricultural Practices and Good Handling Practices Audit Verification Checklist, Version 1.2. Available online: https://www.ams.usda.gov/sites/default/files/media/mGAP Excel Checklist Version 1.3.pdf (accessed on 22 March 2019).
- Murray, K.; Wu, F.; Shi, J.; Xue, S.J.; Warriner, K. Challenges in the microbiological food safety of fresh produce: Limitations of post-harvest washing and the need for alternative interventions. Food Qual. Saf. 2017, 1, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Alum, E.A.; Urom, S.M.O.C.; Ben, C.M.A. Microbiological Contamination of Food: The Mechanisms, Impacts And Prevention. Int. J. Sci. Technol. Res. 2016, 5, 66–78. [Google Scholar]
- Whipps, J.M.; Hand, P.; Pink, D.; Bending, G.D. Phyllosphere microbiology with special reference to diversity and plant genotype. J. Appl. Microbiol. 2008, 105, 1744–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alegbeleye, O.O.; Singleton, I.; Sant’Ana, A.S. Sources and contamination routes of microbial pathogens to fresh produce during field cultivation: A review. Food Microbiol. 2018, 73, 177–208. [Google Scholar] [CrossRef] [PubMed]
- Jay, M.T.; Cooley, M.; Carychao, D.; Wiscomb, G.W.; Sweitzer, R.A.; Crawford-Miksza, L.; Farrar, J.A.; Lau, D.K.; O’Connell, J.; Millington, A.; et al. Escherichia coli O157:H7 in feral swine near spinach fields and cattle, central California coast. Emerg. Infect. Dis. 2007, 13, 1908–1911. [Google Scholar] [CrossRef] [PubMed]
- Akinde, S.B.; Sunday, A.A.; Adeyemi, F.M.; Fakayode, I.B.; Oluwajide, O.O.; Adebunmi, A.A.; Oloke, J.K.; Adebooye, C.O. Microbes in Irrigation Water and Fresh Vegetables: Potential Pathogenic Bacteria Assessment and Implications for Food Safety. Appl. Biosaf. J. ABSA Int. 2016, 21, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Kirby, R.M.; Bartram, J.; Carr, R. Water in food production and processing: Quantity and quality concerns. Food Control 2003, 14, 283–299. [Google Scholar] [CrossRef]
- Zamxaka, M.; Pironcheva, G.; Muyima, N. Microbiological and physico-chemical assessment of the quality of domestic water sources in selected rural communities of the Eastern Cape Province, South Africa. Water SA 2004, 30, 333–340. [Google Scholar] [CrossRef] [Green Version]
- WBCSD World Business Council for Sustainable Development. Facts and Trends: Water; Earthprint Ltd.: Conches-Geneva, Switzerland, 2015. [Google Scholar]
- Viljoen, M.; Kundhlande, G.; Baiphethi, M.N.; Esterhuyse, P.; Botha, J.J.; Anderson, J.J.; Minkley, G.W. An Assessment of the Social and Economic Acceptability of Rainwater Harvesting and Conservation Practices in Selected Peri-Urban and Rural Communities; WRC Report No. 1648/1/12; Water Research Commission: Pretoria, South Africa, 2012. [Google Scholar]
- James, J. Overview of Microbial Hazards in Fresh Fruit and Vegetables Operations. In Microbial Hazard Identification in Fresh Fruit and Vegetables; John Wiley & Sons: Hoboken, NJ, USA, 2006; pp. 1–36. [Google Scholar]
- Leifert, C.; Ball, K.; Volakakis, N.; Cooper, J.M. Control of enteric pathogens in ready-to-eat vegetable crops in organic and ‘low input’ production systems: A HACCP-based approach. J. Appl. Microbiol. 2008, 105, 931–950. [Google Scholar] [CrossRef] [PubMed]
- Pachepsky, Y.; Shelton, D.R.; McLain, J.E.T.; Patel, J.; Mandrell, R.E. Irrigation Waters as a Source of Pathogenic Microorganisms in Produce: A Review. Adv. Agron. 2011, 113, 75–141. [Google Scholar] [CrossRef]
- Gatto D’Andrea, M.L.; Barboza, A.G.S.; Garcés, V.; Alvarez, M.S.R.; Iribarnegaray, M.A.; Liberal, V.I.; Fasciolo, E.G.; van Lier, J.B.; Seghezzo, L. The Use of (Treated) Domestic Wastewater for Irrigation: Current Situation and Future Challenges. Int. J. Water Wastewater Treat. 2015, 1. [Google Scholar] [CrossRef]
- Adefisoye, M.A.; Okoh, A.I. Ecological and Public Health Implications of the Discharge of Multidrug-Resistant Bacteria and Physicochemical Contaminants from Treated Wastewater Effluents in the Eastern Cape, South Africa. Water 2017, 9, 562. [Google Scholar] [CrossRef] [Green Version]
- Steele, M.; Odumeru, J. Irrigation water as source of foodborne pathogens on fruit and vegetables. J. Food Prot. 2004, 67, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- Feachem, R.G.; Bradley, D.J.; Garelick, H.; Mara, D.D.; Feachem, R.G.; Bradley, D.J.; Garelick, H.; Mara, D.D. Sanitation and Disease: Health Aspects of Excreta and Wastewater Management; John Wiley & Sons: Hoboken, NJ, USA, 1983. [Google Scholar]
- Castro-Rosas, J.; Cerna-Cortés, J.F.; Méndez-Reyes, E.; Lopez-Hernandez, D.; Gómez-Aldapa, C.A.; Estrada-Garcia, T. Presence of faecal coliforms, Escherichia coli and diarrheagenic E. coli pathotypes in ready-to-eat salads, from an area where crops are irrigated with untreated sewage water. Int. J. Food Microbiol. 2012, 156, 176–180. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization. Guidelines for the Safe Use of Wastewater, Excreta, and Greywater. Available online: https://www.who.int/water_sanitation_health/publications/gsuweg2/en/ (accessed on 29 March 2019).
- Allende, A.; Monaghan, J. Irrigation Water Quality for Leafy Crops: A Perspective of Risks and Potential Solutions. Int. J. Environ. Res. Public Health 2015, 12, 7457–7477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Páll, E.; Niculae, M.; Kiss, T.; Şandru, C.D.; Spînu, M. Human impact on the microbiological water quality of the rivers. J. Med. Microbiol. 2013, 62, 1635–1640. [Google Scholar] [CrossRef] [PubMed]
- Gemmell, M.E.; Schmidt, S. Microbiological assessment of river water used for the irrigation of fresh produce in a sub-urban community in Sobantu, South Africa. Food Res. Int. 2012, 47, 300–305. [Google Scholar] [CrossRef]
- Ahmed, W.; Sawant, S.; Huygens, F.; Goonetilleke, A.; Gardner, T. Prevalence and occurrence of zoonotic bacterial pathogens in surface waters determined by quantitative PCR. Water Res. 2009, 43, 4918–4928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haley, B.J.; Cole, D.J.; Lipp, E.K. Distribution, diversity, and seasonality of waterborne salmonellae in a rural watershed. Appl. Environ. Microbiol. 2009, 75, 1248–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gannon, V.P.J.; Graham, T.A.; Read, S.; Ziebell, K.; Muckle, A.; Mori, J.; Thomas, J.; Selinger, B.; Townshend, I.; Byrne, J. Bacterial pathogens in rural water supplies in Southern Alberta, Canada. J. Toxicol. Environ. Health Part A 2004, 67, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- Greene, S.; Daly, E.; Talbot, E.; Demma, L.; Holzbauer, S.; Patel, N.; Hill, T.; Walderhaug, M.; Hoekstra, R.; Lynch, M.; et al. Recurrent multistate outbreak of Salmonella Newport associated with tomatoes from contaminated fields, 2005. Epidemiol. Infect. 2008, 136, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hofstra, N.; Franz, E. International Journal of Food Microbiology Impacts of climate change on the microbial safety of pre-harvest leafy green vegetables as indicated by Escherichia coli O157 and Salmonella spp. Int. J. Food Microbiol. 2013, 163, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Nontongana, N.; Sibanda, T.; Ngwenya, E.; Okoh, A. Prevalence and Antibiogram Profiling of Escherichia coli Pathotypes Isolated from the Kat River and the Fort Beaufort Abstraction Water. Int. J. Environ. Res. Public Health 2014, 11, 8213–8227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebert, S.; Burke, J.; Faures, J.M.; Frenken, K.; Hoogeveen, J.; Döll, P.; Portmann, F.T. Groundwater use for irrigation-a global inventory. Hydrol. Earth Syst. Sci. 2010, 14, 1863–1880. [Google Scholar] [CrossRef] [Green Version]
- Onyango, A.E.; Okoth, M.W.; Kunyanga, C.N.; Aliwa, B.O. Microbiological Quality and Contamination Level of Water Sources in Isiolo County in Kenya. J. Environ. Public Health 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IGRAC International Groundwater Resources Assessment Centre; Groundwater and Amp. Climate Change. Available online: https://www.un-igrac.org/areas-expertise/groundwater-climate-change (accessed on 14 October 2019).
- Tyrrel, S.F.; Knox, J.W.; Weatherhead, E.K. Microbiological Water Quality Requirements for Salad Irrigation in the United Kingdom. J. Food Prot. 2006, 69, 2029–2035. [Google Scholar] [CrossRef] [PubMed]
- Chigor, V.N.; Umoh, V.J.; Smith, S.I.; Igbinosa, E.O.; Okoh, A.I. Multidrug Resistance and Plasmid Patterns of Escherichia coli O157 and Other E. coli Isolated from Diarrhoeal Stools and Surface Waters from Some Selected Sources in Zaria, Nigeria. Int. J. Environ. Res. Public Health 2010, 7, 3831–3841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamin, L.; Atwill, E.R.; Jay-Russell, M.; Cooley, M.; Carychao, D.; Gorski, L.; Mandrell, R.E. Occurrence of generic Escherichia coli, E. coli O157 and Salmonella spp. in water and sediment from leafy green produce farms and streams on the Central California coast. Int. J. Food Microbiol. 2013, 165, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Castro-Ibáñez, I.; Gil, M.I.; Tudela, J.A.; Ivanek, R.; Allende, A. Assessment of microbial risk factors and impact of meteorological conditions during production of baby spinach in the Southeast of Spain. Food Microbiol. 2015, 49, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Gardner, T.; Toze, S. Microbiological quality of roof-harvested rainwater and health risks: A review. J. Environ. Qual. 2011, 40, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chidamba, L.; Korsten, L. Pyrosequencing analysis of roof-harvested rainwater and river water used for domestic purposes in Luthengele village in the Eastern Cape Province of South Africa. Environ. Monit. Assess. 2015, 187, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrowsky, P.H.; De Kwaadsteniet, M.; Cloete, T.E.; Khan, W. Distribution of indigenous bacterial pathogens and potential pathogens associated with roof-harvested rainwater. Appl. Environ. Microbiol. 2014, 80, 2307–2316. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Sidhu, J.P.S.; Toze, S. An Attempt to Identify the Likely Sources of Escherichia coli Harboring Toxin Genes in Rainwater Tanks. Environ. Sci. Technol. 2012, 46, 5193–5197. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Brandes, H.; Gyawali, P.; Sidhu, J.P.S.; Toze, S. Opportunistic pathogens in roof-captured rainwater samples, determined using quantitative PCR. Water Res. 2014, 53, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.S.; Cohim, E.; Kalid, R.A. A review on physicochemical and microbiological contamination of roof-harvested rainwater in urban areas. Sustain. Water Qual. Ecol. 2015, 6, 119–137. [Google Scholar] [CrossRef]
- Islam, M.; Morgan, J.; Doyle, M.P.; Phatak, S.C.; Millner, P.; Jiang, X. Persistence of Salmonella enterica Serovar Typhimurium on Lettuce and Parsley and in Soils on Which They were Grown in Fields Treated with Contaminated Manure Composts or Irrigation Water. Foodborne Pathog. Dis. 2004, 1, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.B.; Yaron, S.; Matthews, K.R. Transmission of Escherichia coli O157:H7 from contaminated manure and irrigation water to lettuce plant tissue and its subsequent internalization. Appl. Environ. Microbiol. 2002, 68, 397–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemmell, M.E.; Schmidt, S. Is the microbiological quality of the Msunduzi River (KwaZulu-Natal, South Africa) suitable for domestic, recreational, and agricultural purposes? Environ. Sci. Pollut. Res. 2013, 20, 6551–6562. [Google Scholar] [CrossRef] [PubMed]
- Chigor, V.N.; Sibanda, T.; Okoh, A.I. Studies on the bacteriological qualities of the Buffalo River and three source water dams along its course in the Eastern Cape Province of South Africa. Environ. Sci. Pollut. Res. 2013, 20, 4125–4136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitton, G.; Farrah, S.R.; Ruskin, R.H.; Butner, J.; Chou, Y.J. Survival of Pathogenic and Indicator Organisms in Ground Water. Ground Water 1983, 21, 405–410. [Google Scholar] [CrossRef]
- Avery, L.M.; Williams, A.P.; Killham, K.; Jones, D.L. Survival of Escherichia coli O157:H7 in waters from lakes, rivers, puddles and animal-drinking troughs. Sci. Total Environ. 2008, 389, 378–385. [Google Scholar] [CrossRef] [PubMed]
- VanderZaag, A.C.; Campbell, K.J.; Jamieson, R.C.; Sinclair, A.C.; Hynes, L.G. Survival of Escherichia coli in agricultural soil and presence in tile drainage and shallow groundwater. Can. J. Soil Sci. 2010, 90, 495–505. [Google Scholar] [CrossRef]
- Dowd, S.E.; Pillai, S.D. Survival and transport of selected bacterial pathogens and indicator viruses under sandy aquifer conditions. J. Environ. Sci. Health Part A Environ. Sci. Eng. Toxicol. 1997, 32, 2245–2258. [Google Scholar] [CrossRef]
- Söderström, A.; Österberg, P.; Lindqvist, A.; Jönsson, B.; Lindberg, A.; Blide Ulander, S.; Welinder-Olsson, C.; Löfdahl, S.; Kaijser, B.; De Jong, B.; et al. A Large Escherichia coli O157 Outbreak in Sweden Associated with Locally Produced Lettuce. Foodborne Pathog. Dis. 2008, 5, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, M.; Sundborger, C.; Hergens, M.-P.; Ivarsson, S.; Dryselius, R.; Insulander, M.; Jernberg, C.; Hutin, Y.; Wallensten, A. Barriers to Trace-back in a Salad-associated EHEC Outbreak, Sweden, June 2013. PLoS Curr. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. Investigation of an Escherichia coli O157:H7 Outbreak Associated with Dole Pre-Packaged Spinach. Available online: https://www.cdc.gov/nceh/ehs/docs/investigation_of_an_e_coli_outbreak_associated_with_dole_pre-packaged_spinach.pdf (accessed on 22 March 2019).
- USFDA; CFERT. Investigation of the Taco John’s Escherichia coli O157:H7 Outbreak Associated with Iceberg Lettuce; Department of Health: Sacramento, CA, USA, 2008.
- Harris, L.J.; Bender, J.; Bihn, E.A.; Blessington, T.; Danyluk, M.D.; Delaquis, P.; LeJeune, J.T. A framework for developing research protocols for evaluation of microbial hazards and controls during production that pertain to the quality of agricultural water contacting fresh produce that may be consumed raw. J. Food Prot. 2012, 75, 2251–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, P. Irrigation: Definition, Sources and Methods of Irrigation. Available online: http://www.yourarticlelibrary.com/irrigation/irrigation-definition-sources-and-methods-of-irrigation/31452 (accessed on 28 March 2019).
- CDC Centers for Disease Control and Prevention. Types of Agricultural Water Use. Available online: https://www.cdc.gov/healthywater/other/agricultural/types.html (accessed on 29 March 2019).
- Fonseca, J.M.; Fallon, S.D.; Sanchez, C.A.; Nolte, K.D. Escherichia coli survival in lettuce fields following its introduction through different irrigation systems. J. Appl. Microbiol. 2011, 110, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.B.; Pang, H.-J.; Matthews, K.R. Persistence of Escherichia coli O157:H7 on lettuce plants following spray irrigation with contaminated water. J. Food Prot. 2003, 66, 2198–2202. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.B.; Potenski, C.J.; Matthews, K.R. Effect of irrigation method on transmission to and persistence of Escherichia coli O157:H7 on lettuce. J. Food Prot. 2002, 65, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Van der Linden, I.; Cottyn, B.; Uyttendaele, M.; Vlaemynck, G.; Heyndrickx, M.; Maes, M. Survival of enteric pathogens during butterhead lettuce growth: Crop stage, leaf age, and irrigation. Foodborne Pathog. Dis. 2004, 10, 485–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, M.C.; Webb, C.C.; Davey, L.E.; Payton, A.S.; Flitcroft, I.D.; Doyle, M. Internalization and Fate of Escherichia coli O157:H7 in Leafy Green Phyllosphere Tissue Using Various Spray Conditions. J. Food Prot. 2014, 77, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Kisluk, G.; Yaron, S. Presence and persistence of Salmonella enterica serotype typhimurium in the phyllosphere and rhizosphere of spray-irrigated parsley. Appl. Environ. Microbiol. 2012, 78, 4030–4036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, M.I.; Selma, M.V.; Suslow, T.; Jacxsens, L.; Allende, A.; Gil, M.I.; Selma, M.V.; Suslow, T.; Jacxsens, L. Pre- and Postharvest Preventive Measures and Intervention Strategies to Control Microbial Food Safety Hazards of Fresh Leafy Vegetables. Crit. Rev. Food Sci. Nutr. 2015, 55, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Stine, S.W.; Song, I.; Choi, C.Y.; Gerba, C.P. Effect of relative humidity on preharvest survival of bacterial and viral pathogens on the surface of cantaloupe, lettuce, and bell peppers. J. Food Prot. 2005, 68, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Stine, S.W.; Song, I.; Choi, C.Y.; Gerba, C.P. Application of microbial risk assessment to the development of standards for enteric pathogens in water used to irrigate fresh produce. J. Food Prot. 2005, 68, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C.; Webb, C.C.; Diaz-Perez, J.C.; Phatak, S.C.; Silvoy, J.J.; Davey, L.; Payton, A.S.; Liao, J.; Ma, L.; Doyle, M.P. Infrequent internalization of Escherichia coli O157:H7 into field-grown leafy greens. J. Food Prot. 2010, 73, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Chen, J.; Brackett, R.E.; Beuchat, L.R. Survival of Salmonella on tomatoes stored at high relative humidity, in soil, and on tomatoes in contact with soil. J. Food Prot. 2002, 65, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Lang, N.L.; Smith, S.R. Influence of soil type, moisture content and biosolids application on the fate of Escherichia coli in agricultural soil under controlled laboratory conditions. J. Appl. Microbiol. 2007, 103, 2122–2131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ma, L.; Beuchat, L.R.; Erickson, M.C.; Phelan, V.H.; Doyle, M.P. Lack of internalization of Escherichia coli O157:H7 in lettuce (Lactuca sativa L.) after leaf surface and soil inoculation. J. Food Prot. 2009, 72, 2028–2037. [Google Scholar] [CrossRef] [PubMed]
- Holley, R.A.; Arrus, K.M.; Ominski, K.H.; Tenuta, M.; Blank, G. Survival in Manure-Treated Soils during Simulated Seasonal Temperature Exposure. J. Environ. Qual. 2006, 35, 1170. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.C.; Habteselassie, M.Y.; Liao, J.; Webb, C.C.; Mantripragada, V.; Davey, L.E.; Doyle, M.P. Examination of factors for use as potential predictors of human enteric pathogen survival in soil. J. Appl. Microbiol. 2014, 116, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, R.; Gordon, R.; Sharples, K.; Stratton, G.; Madani, A. Movement and persistence of fecal bacteria in agricultural soils and subsurface drainage water: A review. Can. Biosyst. Eng. 2002, 44, 1.1–1.9. [Google Scholar]
- Mubiru, D.N.; Coyne, M.S.; Grove, J.H. Mortality of Escherichia colt O157:H7 in Two Soils with Different Physical and Chemical Properties. Plant Soil Sci. Fac. Publ. 2000, 7, 1821–1825. [Google Scholar] [CrossRef]
- Tate, R. Cultural and environmental factors affecting the longevity of Escherichia coli in Histosols. Appl. Environ. Microbiol. 1978, 35, 925–929. [Google Scholar] [PubMed]
- Fongaro, G.; García-González, M.C.; Hernández, M.; Kunz, A.; Barardi, C.R.M.; Rodríguez-Lázaro, D. Different Behavior of Enteric Bacteria and Viruses in Clay and Sandy Soils after Biofertilization with Swine Digestate. Front. Microbiol. 2017, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyberg, K.A.; Ottoson, J.R.; Vinnerås, B.; Albihn, A. Fate and survival of Salmonella Typhimurium and Escherichia coli O157:H7 in repacked soil lysimeters after application of cattle slurry and human urine. J. Sci. Food Agric. 2014, 94, 2541–2546. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, J.V.; Karns, J.S. Persistence of Escherichia coli O157:H7 in soil and on plant roots. Environ. Microbiol. 2002, 4, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Reynnells, R. Importance of Soil Amendments: Survival of Bacterial Pathogens in Manure and Compost Used as Organic Fertilizers. Microbiol. Spectr. 2016, 4, 159–175. [Google Scholar] [CrossRef] [Green Version]
- Naganandhini, S.; Kennedy, Z.J.; Uyttendaele, M.; Balachandar, D. Persistence of Pathogenic and Non-Pathogenic Escherichia coli Strains in Various Tropical Agricultural Soils of India. PLoS ONE 2015, 10, e0130038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagedorn, C.; Hansen, D.T.; Simonson, G.H. Survival and Movement of Fecal Indicator Bacteria in Soil under Conditions of Saturated Flow. J. Environ. Qual. 1978, 7, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Rochelle-Newall, E.J.; Ribolzi, O.; Viguier, M.; Thammahacksa, C.; Silvera, N.; Latsachack, K.; Dinh, R.P.; Naporn, P.; Sy, H.T.; Soulileuth, B.; et al. Effect of land use and hydrological processes on Escherichia coli concentrations in streams of tropical, humid headwater catchments. Sci. Rep. 2016, 6, 32974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leahy, J.G.; Colwell, R.R. Microbial degradation of hydrocarbons in the environment. Microbiol. Rev. 1990, 54, 305–315. [Google Scholar] [PubMed]
- van Veen, J.A.; van Overbeek, L.S.; van Elsas, J.D. Fate and activity of microorganisms introduced into soil. Microbiol. Mol. Biol. Rev. 1997, 61, 121–135. [Google Scholar] [PubMed]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Santamaría, J.; Toranzos, G.A. Enteric pathogens and soil: A short review. Int. Microbiol. 2003, 6, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Mosaddeghi, M.R.; Mahboubi, A.A.; Zandsalimi, S.; Unc, A. Influence of organic waste type and soil structure on the bacterial filtration rates in unsaturated intact soil columns. J. Environ. Manag. 2009, 90, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Schoonover, J.E.; Crim, J.F. An Introduction to Soil Concepts and the Role of Soils in Watershed Management. J. Contemp. Water Res. Educ. 2015, 154, 21–47. [Google Scholar] [CrossRef]
- Diacono, M.; Montemurro, F. Long-term effects of organic amendments on soil fertility: A review. Development 2010, 30, 401–422. [Google Scholar] [CrossRef] [Green Version]
- Weller, D.M. Biological Control of Soil borne Plant Pathogens in the Rhizosphere with Bacteria. Annu. Rev. Phytopathol. 1988, 26, 379–407. [Google Scholar] [CrossRef]
- Beuchat, L.R. Vectors and conditions for preharvest contamination of fruits and vegetables with pathogens capable of causing enteric diseases. Br. Food J. 2006, 108, 38–53. [Google Scholar] [CrossRef]
- Ingham, S.C.; Losinski, J.A.; Andrews, M.P.; Breuer, J.E.; Breuer, J.R.; Wood, T.M.; Wright, T.H. Escherichia coli Contamination of Vegetables Grown in Soils Fertilized with Noncomposted Bovine Manure: Garden-Scale Studies. Appl. Environ. Microbiol. 2004, 70, 6420–6427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.L. Examination of Microbiological Quality of In-Field Leafy Vegetables and Identification of On-Farm Generic Escherichia coli Transmission Dynamics. Bachelor’s Thesis, University of British Columbia, Vancouver, BC, Canada, 2013. [Google Scholar]
- Semenov, A.V.; van Overbeek, L.; Termorshuizen, A.J.; van Bruggen, A.H.C. Influence of aerobic and anaerobic conditions on survival of Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium in Luria–Bertani broth, farm-yard manure and slurry. J. Environ. Manag. 2011, 92, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Heinonen-Tanski, H.; Niskanen, E.M.; Salmela, P.; Lanki, E. Salmonella in animal slurry can be destroyed by aeration at low temperatures. J. Appl. Microbiol. 1998, 85, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Tiquia, S.M.; Tam, N.F.Y.; Hodgkiss, I.J. Salmonella elimination during composting of spent pig litter. Bioresour. Technol. 1998, 63, 193–196. [Google Scholar] [CrossRef]
- Wiley, B.B.; Westerberg, S.C. Survival of Human Pathogens in Composted Sewage. Appl. Environ. Microbiol. 1969, 18, 994–1001. [Google Scholar]
- Kearney, T.E.; Larkin, M.J.; Levett, P.N. The effect of slurry storage and anaerobic digestion on survival of pathogenic bacteria. J. Appl. Bacteriol. 1993, 74, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jiang, X. The growth potential of Escherichia coli O157:H7, Salmonella spp. and Listeria monocytogenes in dairy manure-based compost in a greenhouse setting under different seasons. J. Appl. Microbiol. 2010, 109, 2095–2104. [Google Scholar] [CrossRef] [PubMed]
- Franz, E.; van Diepeningen, A.D.; de Vos, O.J.; van Bruggen, A.H.C. Effects of Cattle Feeding Regimen and Soil Management Type on the Fate of Escherichia coli O157:H7 and Salmonella enterica Serovar Typhimurium in Manure, Manure-Amended Soil, and Lettuce. Appl. Environ. Microbiol. 2005, 71, 6165–6174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, E.; van Bruggen, A.H.C. Ecology of E. coli O157:H7 and Salmonella enterica in the primary vegetable production chain. Crit. Rev. Microbiol. 2008, 34, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Hassen, A.; Belguith, K.; Jedidi, N.; Cherif, A.; Cherif, M.; Boudabous, A. Microbial characterization during composting of municipal solid waste. Bioresour. Technol. 2001, 80, 217–225. [Google Scholar] [CrossRef]
- USDA United States Department of Agriculture. USDA National Organic Program 7 CFR Part 2015. 26 October 2015. Available online: https://www.nofany.org/files/NOP_Organic_Regulations.10.26.15.pdf (accessed on 1 May 2019).
- Altizer, S.; Bartel, R.; Han, B.A. Animal Migration and Infectious Disease Risk. Science 2011, 331, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC Centers for Disease Control and Prevention. Outbreaks of Escherichia coli O157:H7 Infection and Cryptosporidiosis Associated with Drinking Unpasteurized Apple Cider—Connecticut and New York, October 1996. Available online: http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/9011776 (accessed on 7 May 2019).
- Ackers, M.L.; Mahon, B.E.; Leahy, E.; Goode, B.; Damrow, T.; Hayes, P.S.; Bibb, W.F.; Rice, D.H.; Barrett, T.J.; Hutwagner, L.; et al. An outbreak of Escherichia coli O157:H7 infections associated with leaf lettuce consumption. J. Infect. Dis. 1998, 177, 1588–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuorti, J.P.; Niskanen, T.; Hallanvuo, S.; Mikkola, J.; Kela, E.; Hatakka, M.; Fredriksson-Ahomaa, M.; Lyytikäinen, O.; Siitonen, A.; Korkeala, H.; et al. A Widespread Outbreak of Yersinia pseudotuberculosis O:3 Infection from Iceberg Lettuce. J. Infect. Dis. 2004, 189, 766–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méric, G.; Hitchings, M.D.; Pascoe, B.; Sheppard, S.K. From Escherich to the Escherichia coli genome. Lancet Infect. Dis. 2016, 16, 634–636. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Steitz, T.A. Crystal Structures of the E. coli Transcription Initiation Complexes with a Complete Bubble. Mol. Cell 2015, 58, 534–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagier, J.C.; Hugon, P.; Khelaifia, S.; Fournier, P.E.; La Scola, B.; Raoult, D. The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota. Clin. Microbiol. Rev. 2015, 28, 237–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, N.; Neumann, P.; Konevega, A.L.; Bock, L.V.; Ficner, R.; Rodnina, M.V.; Stark, H. Structure of the E. coli ribosome–EF-Tu complex at <3 A resolution by Cs-corrected cryo-EM. Nature 2015, 520, 567–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Elsas, J.D.; Semenov, A.V.; Costa, R.; Trevors, J.T. Survival of Escherichia coli in the environment: Fundamental and public health aspects. ISME J. 2011, 5, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adefisoye, M.A.; Okoh, A.I. Identification and antimicrobial resistance prevalence of pathogenic Escherichia coli strains from treated wastewater effluents in Eastern Cape, South Africa. Microbiol. Open 2016, 5, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahandeh, N.; Ranjbar, R.; Behzadi, P.; Behzadi, E. Uropathogenic Escherichia coli virulence genes: Invaluable approaches for designing dna microarray probes. Cent. Eur. J. Urol. 2015, 68, 452–458. [Google Scholar] [CrossRef] [Green Version]
- CDC Centers for Disease Control and Prevention. E. coli and Food Safety. Available online: https://www.cdc.gov/features/ecoliinfection/index.html (accessed on 11 October 2019).
- Goswami, K.; Chen, C.; Xiaoli, L.; Eaton, K.A.; Dudley, E.G. Co-culturing Escherichia coli O157:H7 with a non-pathogenic E. coli strain increases toxin production and virulence in a germ-free mouse model. Infect. Immun. 2015, 4185–4193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakre, T.; Warke, S.; Bobade, S.; Kalorey, D.R. Characterization of E. coli pathotypes of bovine and livestock farm environment origin. Indian J. Anim. Res. 2017, 51, 1125–1129. [Google Scholar] [CrossRef]
- Rasheed, M.U.; Thajuddin, N.; Ahamed, P.; Teklemariam, Z.; Jamil, K. Antimicrobial drug resistance in strains of Escherichia coli isolated from food sources. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupindu, A.M. Epidemiology of Shiga toxin-producing Escherichia coli O157:H7 in Africa in review. S. Afr. J. Infect. Dis. 2018, 33, 24–30. [Google Scholar] [CrossRef]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157:H7 outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Rahal, E.A.; Kazzi, N.; Nassar, F.J.; Matar, G.M. Escherichia coli O157:H7—Clinical aspects and novel treatment approaches. Front. Cell. Infect. Microbiol. 2012, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iweriebor, B.C.; Iwu, C.J.; Obi, L.C.; Nwodo, U.U.; Okoh, A.I. Multiple antibiotic resistances among Shiga toxin producing Escherichia coli O157 in feces of dairy cattle farms in Eastern Cape of South Africa. BMC Microbiol. 2015, 15, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.Y.; Yoon, J.W.; Hovde, C.J. A Brief Overview of Escherichia coli O157:H7 and Its Plasmid O157. J. Microbiol. Biotechnol. 2010, 20, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, C.A.; Rubinelli, P.M.; Park, H.S.; Carbonero, F.; Ricke, S.C. Shiga toxin-producing Escherichia coli in food: Incidence, ecology, and detection strategies. Food Control 2016, 59, 407–419. [Google Scholar] [CrossRef]
- Pruimboom-Brees, I.M.; Morgan, T.W.; Ackermann, M.R.; Nystrom, E.D.; Samuel, J.E.; Cornick, N.A.; Moon, H.W. Cattle lack vascular receptors for Escherichia coli O157:H7 Shiga toxins. Proc. Natl. Acad. Sci. USA 2000, 97, 10325–10329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, Y.; Irie, Y.; Sawada, T.; Nakazawa, M. A 3-Year Epidemiological Surveillance of Escherichia coli O157:H7 in Dogs and Cats in Japan. J. Vet. Med. Sci. 2010, 72, 791–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urdahl, A.M.; Beutin, L.; Skjerve, E.; Zimmermann, S.; Wasteson, Y. Animal host associated differences in Shiga toxin-producing Escherichia coli isolated from sheep and cattle on the same farm. J. Appl. Microbiol. 2003, 95, 92–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, S.; Vidal, D.; Herrera-León, S.; Sánchez, S. Sorbitol-Fermenting, β-Glucuronidase–Positive, Shiga toxin–negative Escherichia coli O157:H7 in free-ranging red deer in South-Central Spain. Foodborne Pathog. Dis. 2011, 8, 1313–1315. [Google Scholar] [CrossRef] [PubMed]
- Mersha, G.; Asrat, D.; Zewde, B.M.; Kyule, M. Occurrence of Escherichia coli O157:H7 in faeces, skin and carcasses from sheep and goats in Ethiopia. Lett. Appl. Microbiol. 2010, 50, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Lengacher, B.; Kline, T.R.; Harpster, L.; Williams, M.L.; LeJeune, J.T. Low prevalence of Escherichia coli O157:H7 in horses in Ohio, USA. J. Food Prot. 2010, 73, 2089–2092. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.Y.; Mazin, H.; Saadi, A.; Hussein, S.O. Detection of Escherichia coli O157 in vegetables. IOSR J. Agric. Vet. Sci. 2013, 6, 16–18. [Google Scholar] [CrossRef]
- Agger, M.; Scheutz, F.; Villumsen, S.; Mølbak, K.; Petersen, A.M. Antibiotic treatment of verocytotoxin-producing Escherichia coli (VTEC) infection: A systematic review and a proposal. J. Antimicrob. Chemother. 2015, 70, 2440–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amézquita-López, B.A.; Quiñones, B.; Soto-Beltrán, M.; Lee, B.G.; Yambao, J.C.; Lugo-Melchor, O.Y.; Chaidez, C. Antimicrobial resistance profiles of Shiga toxin-producing Escherichia coli O157 and Non-O157 recovered from domestic farm animals in rural communities in Northwestern Mexico. Antimicrob. Resist. Infect. Control 2016, 5, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncuoglu, M.; Bilir Ormanci, F.S.; Ayaz, N.D.; Erol, I. Antibiotic resistance of Escherichia coli O157:H7 isolated from cattle and sheep. Ann. Microbiol. 2010, 60, 489–494. [Google Scholar] [CrossRef]
- Abakpa, G.O.; Umoh, V.J.; Ameh, J.B.; Yakubu, S.E.; Kwaga, J.K.P.; Kamaruzaman, S. Diversity and antimicrobial resistance of Salmonella enterica isolated from fresh produce and environmental samples. Environ. Nanotechnol. Monit. Manag. J. 2015, 3, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Sodagari, H.R.; Mashak, Z.; Ghadimianazar, A. Prevalence and antimicrobial resistance of Salmonella serotypes isolated from retail chicken meat and giblets in Iran. J. Infect. Dev. Ctries. 2014, 9, 463–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marks, F.; Von Kalckreuth, V.; Aaby, P.; Adu-sarkodie, Y.; Ahmed, M.; Tayeb, E.; Ali, M.; Aseff, A.; Baker, S.; Biggs, H.M.; et al. Incidence of invasive salmonella disease in sub-Saharan Africa: A multicentre population-based surveillance study. Lancet Glob. Health 2017, 5, e310–e323. [Google Scholar] [CrossRef] [Green Version]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Salmonella Infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariuki, S.; Gordon, M.A.; Feasey, N.; Parry, C.M. Antimicrobial resistance and management of invasive Salmonella disease. Vaccine 2015, 33, S21–S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.-A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States-major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Shi, Z.; Wei, J.; Ma, Z. A brief review of foodborne zoonoses in China. Epidemiol. Infect. 2011, 139, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Ran, L.; Wu, S.; Ke, B.; He, D.; Yang, X.; Zhang, Y.; Ke, C.; Klena, J.D.; Yan, M.; et al. Laboratory-Based Surveillance of Non-typhoidal Salmonella Infections in Guangdong Province, China. Foodborne Pathog. Dis. 2012, 9, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The Global Burden of Nontyphoidal Salmonella Gastroenteritis. Clin. Infect. Dis. 2010, 50, 882–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Gu, G.; Ginn, A.; Giurcanu, M.C.; Adams, P.; Vellidis, G.; Van Bruggen, A.H.C.; Danyluk, M.D.; Wright, A.C. Distribution and Characterization of Salmonella enterica Isolates from Irrigation Ponds in the Southeastern United States. Appl. Environ. Microbiol. 2015, 81, 4376–4387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorski, L.; Parker, C.T.; Liang, A.; Cooley, M.B.; Jay-Russell, M.T.; Gordus, A.G.; Atwill, E.R.; Mandrell, R.E. Prevalence, Distribution, and Diversity of Salmonella enterica in a Major Produce Region of California. Appl. Environ. Microbiol. 2011, 77, 2734–2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajabi, M.; Jones, M.; Hubbard, M.; Rodrick, G.; Wright, A.C. Distribution and Genetic Diversity of Salmonella enterica in the Upper Suwannee River. Int. J. Microbiol. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, M.C.; Doyle, M.P. Improving Food Safety Through a One Health Approach: Workshop Summary; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Lee, S.-J.; Liang, L.; Juarez, S.; Nanton, M.R.; Gondwe, E.N.; Msefula, C.L.; Kayala, M.A.; Necchi, F.; Heath, J.N.; Hart, P.; et al. Identification of a common immune signature in murine and human systemic Salmonellosis. Proc. Natl. Acad. Sci. USA 2012, 109, 4998–5003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Huang, J.; Wu, Q.; Zhang, J.; Liu, S.; Guo, W.; Cai, S.; Yu, S. Prevalence, antimicrobial resistance and genetic diversity of Salmonella isolated from retail ready-to-eat foods in China. Food Control 2016, 60, 50–56. [Google Scholar] [CrossRef]
- Sansonetti, P.J. The Bacterial Weaponry: Lessons from Shigella. Ann. N. Y. Acad. Sci. 2006, 1072, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Nave, H.H.; Mansouri, S.; Emaneini, M.; Moradi, M. Distribution of genes encoding virulence factors and molecular analysis of Shigella spp. isolated from patients with diarrhea in Kerman, Iran. Microb. Pathog. 2016, 92, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh-hesar, M.; Bakhshi, B.; Najar-peerayeh, S. Clonal dissemination of a single Shigella sonnei strain among Iranian children during Fall 2012 in Tehran, I.R. Iran. Infect. Genet. Evolut. 2015, 34, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Shimamoto, T. Molecular characterization of multidrug-resistant Shigella spp. of food origin. Int. J. Food Microbiol. 2015, 194, 78–82. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization (WHO). Shigellosis: Disease Burden, Epidemiology and Case Management. Wkly. Epidemiol. Rec. 2005, 80, 94–99. [Google Scholar]
- Bakhshi, B.; Bayat, B.; Rastegar, A. Multiple-locus variable-number tandem-repeat analysis (MLVA ) of Shigella sonnei isolates of 2012 outbreak I. R. Iran. Microb. Pathog. 2017, 102, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Farshad, S.; Ranjbar, R.; Hosseini, M. Molecular Genotyping of Shigella sonnei Strains Isolated from Children with Bloody Diarrhea Using Pulsed Field Gel Electrophoresis on the Total Genome and PCR-RFLP of IpaH and IpaBCD Genes. Jundishapur J. Microbiol. 2015, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]
- CDC Centers for Disease Control and Prevention. Outbreaks of Multidrug-Resistant Shigella sonnei Gastroenteritis Associated with Day Care Centers -Kansas, Kentucky, and Missouri. 2005. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm5539a3.htm (accessed on 1 May 2019).
- Qureshi, S. Klebsiella Infections: Background, Pathophysiology, Epidemiology of Klebsiellae. Available online: https://emedicine.medscape.com/article/219907-overview (accessed on 16 May 2018).
- Podschun, R.; Pietsch, S.; Höller, C.; Ullmann, U. Incidence of Klebsiella species in surface waters and their expression of virulence factors. Appl. Environ. Microbiol. 2001, 67, 3325–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ristuccia, P.A.; Cunha, B.A. Klebsiella. Infect. Control 1984, 5, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Hudson, C.M.; Bent, Z.W.; Meagher, R.J.; Williams, K.P. Resistance Determinants and Mobile Genetic Elements of an NDM-1-Encoding Klebsiella pneumoniae Strain. PLoS ONE 2014, 9, e99209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathisuwan, S.; Burgess, D.S.; Lewis, J.S. Extended-Spectrum β-Lactamases: Epidemiology, Detection, and Treatment. Pharmacotherapy 2001, 21, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Hendrik, T.C.; Voor in’t holt, A.F.; Vos, M.C. Clinical and Molecular Epidemiology of Producing Klebsiella spp: A Systematic Review and Meta-Analyses. PLoS ONE 2015, 10, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Nada, T.; Baba, H.; Kawamura, K.; Ohkura, T.; Torii, K.; Ohta, M. A small outbreak of third generation cephem-resistant Citrobacter freundii infection on a surgical ward. Jpn. J. Infect. Dis. 2004, 57, 181–182. [Google Scholar] [PubMed]
- Bae, I.K.; Park, I.; Lee, J.J.; Sun, H.I.; Park, K.S.; Lee, J.E.; Ahn, J.H.; Lee, S.H.; Woo, G.-J. Novel variants of the qnrB gene, qnrB22 and qnrB23, in Citrobacter werkmanii and Citrobacter freundii. Antimicrob. Agents Chemother. 2010, 54, 3068–3069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Lan, R.; Liu, L.; Wang, Y.; Zhang, Y.; Wang, Y.; Xu, J. Antimicrobial Resistance and Cytotoxicity of Citrobacter spp. in Maanshan Anhui Province, China. Front. Microbiol. 2017, 8, 1357. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, K.P.; Ranjan, N. Citrobacter: An emerging health care associated urinary pathogen. Urol. Ann. 2013, 5, 313–314. [Google Scholar] [PubMed]
- Bai, L.; Xia, S.; Lan, R.; Liu, L.; Ye, C.; Wang, Y.; Jin, D.; Cui, Z.; Jing, H.; Xiong, Y.; et al. Isolation and Characterization of Cytotoxic, Aggregative Citrobacter freundii. PLoS ONE 2012, 7, e33054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanamori, H.; Yano, H.; Hirakata, Y.; Endo, S.; Arai, K.; Ogawa, M.; Shimojima, M.; Aoyagi, T.; Hatta, M.; Yamada, M.; et al. High prevalence of extended-spectrum Beta-lactamases and qnr determinants in Citrobacter species from Japan: Dissemination of CTX-M-2. J. Antimicrob. Chemother. 2011, 66, 2255–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, W.-S.; Yunos, N.; Tan, P.-W.; Mohamad, N.; Adrian, T.-G.-S.; Yin, W.-F.; Chan, K.-G. Freshwater-Borne Bacteria Isolated from a Malaysian Rainforest Waterfall Exhibiting Quorum Sensing Properties. Sensors 2014, 14, 10527–10537. [Google Scholar] [CrossRef] [PubMed]
- Cabral, J.P.S. Water Microbiology. Bacterial Pathogens and Water. Int. J. Environ. Res. Public Health 2010, 7, 3657–3703. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, D.J.; Marcon, M.J. Etiologic Agents of Infectious Diseases. Principles and Practice of Pediatric Infectious Diseases, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Fraser, S.L.; Sinave, C.P. Enterobacter Infections: Practice Essentials, Background, Pathophysiology. Available online: https://emedicine.medscape.com/article/216845-overview (accessed on 20 May 2018).
- Rogers, K. Enterobacter. Available online: https://www.britannica.com/science/Enterobacter (accessed on 18 May 2018).
- Shaikh, S.; Fatima, J.; Shakil, S.; Danish Rizvi, S.M.; Amjad Kamal, M. Antibiotic resistance and extended spectrum beta-lactamases: Types, epidemiology and treatment. Saudi J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, C. Antibiotics: Actions, origins, resistance. Protein Sci. 2004, 13, 3059–3060. [Google Scholar] [CrossRef] [Green Version]
- Dzidic, S.; Bedekovic, V. Horizontal gene transfer-emerging multidrug resistance. Acta Pharmacol. Sin. 2003, 24, 519–526. [Google Scholar] [PubMed]
- Grudén, M. Antibiotic Resistance Associated with Bacteria in Irrigation Water—A Case Study of Irrigation Ponds in Southern Sweden; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2013. [Google Scholar]
- CDC Centers for Disease Control and Prevention. Office of Infectious Disease. Antibiotic Resistance Threats in the United States, Antibiotic/Antimicrobial Resistance/CDC. 2013. Available online: https://www.cdc.gov/drugresistance/threat-report-2013/index.html (accessed on 3 January 2018).
- Gross, M. Antibiotics in crisis. Feature 2000, 23, 1063–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocsis, B.; Szabó, D. Antibiotic resistance mechanisms in Enterobacteriaceae. Formatex 2013, 251–257. [Google Scholar]
- Bae, D.; Mezal, E.H.; Smiley, R.D.; Cheng, C.; Khan, A.A. The sub-species characterization and antimicrobial resistance of Listeria monocytogenes isolated from domestic and imported food products from 2004 to 2011. Food Res. Int. J. 2014, 64, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Komora, N.; Bruschi, C.; Magalhães, R.; Ferreira, V.; Teixeira, P. Survival of Listeria monocytogenes with different antibiotic resistance patterns to food-associated stresses. Int. J. Food Microbiol. 2017, 245, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.J.; Wa, E.; Chen, J.C.; Katarzyna, K.; Devlieghere, F.; Van Meervenne, E.; Osek, J.; Wieczorek, K.; Bania, J. Listeria monocytogenes: An examination of food chain factors potentially contributing to antimicrobial resistance. Food Microbiol. J. 2016, 54, 178–189. [Google Scholar] [CrossRef]
- Wang, X.-M.; Lü, X.-F.; Yin, L.; Liu, H.-F.; Zhang, W.-J.; Si, W.; Yu, S.-Y.; Shao, M.-L.; Liu, S.-G. Occurrence and antimicrobial susceptibility of Listeria monocytogenes isolates from retail raw foods. Food Control 2013, 32, 153–158. [Google Scholar] [CrossRef]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in Fresh Produce: Outbreaks, Prevalence and Contamination Levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakicevic, B.; Nastasijevic, I. Listeria monocytogenes in retail establishments: Contamination routes and control strategies. Food 2017, 33, 247–269. [Google Scholar] [CrossRef]
- Noll, M.; Kleta, S.; Al, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health 2017, 12, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Välimaa, A.-L.; Tilsala-Timisjärvi, A.; Virtanen, E. Rapid detection and identification methods for Listeria monocytogenes in the food chain—A review. Food Control 2015, 55, 103–114. [Google Scholar] [CrossRef]
- Wu, S.; Wu, Q.; Zhang, J.; Chen, M.; Yan, Z.; Hu, H. Listeria monocytogenes Prevalence and Characteristics in Retail Raw Foods in China. PLoS ONE 2015, 10, e0136682. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization. Listeriosis. Available online: https://www.who.int/news-room/fact-sheets/detail/listeriosis (accessed on 11 October 2019).
- Lomonaco, S.; Nucera, D.; Filipello, V. Infection, Genetics and Evolution The evolution and epidemiology of Listeria monocytogenes in Europe and the United States. Infect. Genet. Evolut. 2015, 35, 172–183. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization. Listeriosis—South Africa. Available online: https://www.who.int/csr/don/28-march-2018-listeriosis-south-africa/en/ (accessed on 2 May 2019).
- NLIMT National Listeria Incident Management Team. Listeriosis Outbreak Situation Report. 21 May 2018. Available online: http://www.nicd.ac.za/wp-content/uploads/2018/05/Listeriosis-outbreak-situation-report-draft-_21May2018_final.pdf (accessed on 21 May 2018).
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of Antibiotic Resistance in Listeria monocytogenes Isolated from Food Products: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292. [Google Scholar] [CrossRef] [Green Version]
- Abdollahzadeh, E.; Ojagh, S.M.; Hosseini, H.; Ghaemi, E.A.; Irajian, G.; Heidarlo, M.N. Antimicrobial resistance of Listeria monocytogenes isolated from seafood and humans in Iran. Microb. Pathog. 2016, 100, 70–74. [Google Scholar] [CrossRef] [PubMed]
- WHO World Health Organization. Antimicrobial resistance in the Food Chain. Available online: https://www.who.int/foodsafety/areas_work/antimicrobial-resistance/amrfoodchain/en/ (accessed on 7 February 2019).
- WHO World Health Organization. Antibiotic resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antibiotic-resistance (accessed on 12 October 2019).
- Åström, J.; Carlander, A.; Sahlén, K.; Stenström, T.A. Fecal Indicator and Pathogen Reduction in Vegetation Microcosms. Water Air Soil Pollut. 2006, 176, 375–387. [Google Scholar] [CrossRef]
- Oliver, D.M.; Haygarth, P.M.; Clegg, C.D.; Heathwaite, A.L. Differential, E. coli Die-Off Patterns Associated with Agricultural Matrices. Environ. Sci. Technol. 2006, 40, 5710–5716. [Google Scholar] [CrossRef] [PubMed]
- Moynihan, E.L.; Richards, K.G.; Ritz, K.; Tyrrel, S.F.; Brennan, F.P. Impact of soil type, biology and temperature on the survival of non-toxigenic Escherichia coli O157. Biol. Environ. Proc. R. Ir. Acad. 2013, 113, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, D.; Huber, A.; Weese, S.J.; Warriner, K. Persistence of Clostridium difficile in wastewater treatment-derived biosolids during land application or windrow composting. J. Appl. Microbiol. 2016, 120, 312–320. [Google Scholar] [CrossRef] [PubMed]
- FAO Food and Agriculture Organization of the United Nations. Good Agricultural Practices for Greenhouse Vegetable Production in the South East European Countries; FAO Plant Production and Protection Paper; FAO: Rome, Italy, 2017. [Google Scholar]
- FAO Food and Agriculture Organization of the United Nations. Antimicrobial Resistance (AMR) in Aquaculture; FAO: Geneva, Switzerland, 2017. [Google Scholar]
- Weller, D.; Wiedmann, M.; Strawn, L.K. Spatial and Temporal Factors Associated with an Increased Prevalence of Listeria monocytogenes in Spinach Fields in New York State. Appl. Environ. Microbiol. 2015, 81, 6059–6069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Généreux, M.; Breton, M.J.; Fairbrother, J.M.; Fravalo, P.; Côté, C. Persistence of Indicator and Pathogenic Microorganisms in Broccoli following Manure Spreading and Irrigation with Fecally Contaminated Water: Field Experiment. J. Food Prot. 2015, 78, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; He, Z.; Powell, C.A.; Stoffella, P.J. Survival of Escherichia coli in soil with modified microbial community composition. Soil Biol. Biochem. 2011, 43, 1591–1599. [Google Scholar] [CrossRef]
- Oladeinde, A.; Bohrmann, T.; Wong, K.; Purucker, S.T.; Bradshaw, K.; Brown, R.; Snyder, B.; Molina, M. Decay of fecal indicator bacterial populations and bovine-associated source-tracking markers in freshly deposited cow pats. Appl. Environ. Microbiol. 2014, 80, 110–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayrapetyan, M.; Williams, T.C.; Baxter, R.; Oliver, J.D. Viable but Nonculturable and Persister Cells Coexist Stochastically and Are Induced by Human Serum. Infect. Immun. 2015, 83, 4194–4203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kussell, E.; Kishony, R.; Balaban, N.Q.; Leibler, S. Bacterial persistence: A model of survival in changing environments. Genetics 2005, 169, 1807–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stier, R.F.; Nagle, N.E. Growers Beware: Adopt GAPs or Else. Available online: https://www.foodsafetymagazine.com/magazine-archive1/octobernovember-2001/growers-beware-adopt-gaps-or-else/ (accessed on 7 May 2019).


{kind=link}
| Fresh Produce | Bacterial Pathogens | Number of Cases (Mortalities) | Country | Year | References |
|---|---|---|---|---|---|
| Fresh papayas | Salmonella Uganda | 71 (0) | USA | 2019 | [36] |
| Pre-cut melon | Salmonella Carrau | 117 (0) | USA | 2019 | [37] |
| Romaine lettuce | STEC E. coli O157:H7 | 62 (0) | USA | 2018 | [38] |
| Leafy greens | E. coli O157:H7 | 25 (1) | USA | 2018 | [39] |
| Pre-cut melons | Salmonella adelaide | 77 (0) | USA | 2018 | [40] |
| Alfalfa and raw clover sprouts | E. coli | 59 (0) | USA | 2017 | [24] |
| Spinach | E. coli | 199 (3) | USA | 2017 | [24] |
| Apples | Listeria monocytogenes | 35 (7) | USA | 2017 | [24] |
| Bean sprouts | Listeria monocytogenes | 5 (2) | USA | 2017 | [24] |
| Cantaloupe | Listeria monocytogenes | 147 (33) | USA | 2017 | [24] |
| Alfalfa and raw clover sprouts | Salmonella spp. | 506 (0) | USA | 2017 | [24] |
| Cucumbers | Salmonella spp. | 991 (6) | USA | 2017 | [24] |
| Bean sprouts | Salmonella spp. | 115 (0) | USA | 2017 | [24] |
| Mangoes | Salmonella spp. | 127 (0) | USA | 2017 | [24] |
| Papayas | Salmonella spp. | 106 (0) | USA | 2017 | [24] |
| Jalepenos and serrano peppers | Salmonella spp. | 1442 (2) | USA | 2017 | [24] |
| Cantaloupe | Salmonella spp. | 332 (3) | USA | 2017 | [24] |
| Tomatoes | Salmonella spp. | 111 (0) | USA | 2017 | [24] |
| Lettuce | E. coli | 34 (NS) | Canada | 2017 | [24] |
| Cantaloupe | Salmonella spp. | NS | Canada | 2017 | [24] |
| Watercress | E. coli O157 | NS | UK | 2016 | [41] |
| Lettuce, cucumber | E. coli 096 | 50 (NS) | UK | 2016 | [41] |
| Alfalfa sprouts | STEC E. coli O157:H7 | 11 (0) | USA | 2016 | [42] |
| Frozen vegetables | Listeria monocytogenes | 9 (3) | USA | 2016 | [43] |
| Alfalfa sprouts | Salmonella Muenchen (25 people) and Salmonella Kentucky (1 person) | 26 (0) | USA | 2016 | [44] |
| Packaged salads | Listeria monocytogenes | 19 (1) | USA | 2016 | [45] |
| Imported cucumbers | Salmonella Poona | 907 (6) | USA | 2015 | [46] |
| Caramel apples | Listeria monocytogenes | 35 (7) | USA | 2014 | [47] |
| Beans sprouts | Salmonella Enteritidis | 115 (0) | USA | 2014 | [48] |
| Raw clover sprouts | STEC E. coli O121 | 19 (0) | USA | 2014 | [49] |
| Lettuce, cucumber | Enteroinvasive E. coli O96 | 50 | UK | 2014 | [50] |
| Salads | Salmonella Singapore | 4 | UK | 2014 | [50] |
| Watercress | Verocytotoxin-producing E. coli O157 | NS | UK | 2013 | [50] |
| RTE salads | STEC E. coli O157:H7 | 33 (0) | USA | 2013 | [51] |
| Imported cucumbers | Salmonella Saintpaul | 84 (0) | USA | 2013 | [52] |
| Organic Spinach and Spring Mix Blend | STEC E. coli O157:H7 | 33 (0) | USA | 2012 | [53] |
| Mangoes | Salmonella Braenderup | 127 (0) | USA | 2012 | [54] |
| Cantaloupe | Salmonella Typhimurium and Salmonella Newport | 261 (3) | USA | 2012 | [55] |
| Raw clover sprouts | STEC E. coli O26 | 29 (0) | USA | 2012 | [56] |
| Romain lettuce | E. coli O157:H7 | 58 | USA | 2011 | [57] |
| Cantaloupes | Listeria monocytogenes | 147 (33) | USA | 2011 | [58] |
| Whole, freshly imported papayas | Salmonella Agona | 106 | USA | 2011 | [59] |
| Alfalfa sprouts and spicy sprouts | Salmonella Enteritidis | 25 (0) | USA | 2011 | [60] |
| Cantaloupe | Salmonella Panama | 20 (0) | USA | 2011 | [58] |
| Vegetable sprouts | E. coli O104:H4 | 3911 (47) | Europe | 2011 | [61,62] |
| Alfalfa sprouts | Salmonella I 4,[5],12:i:- | 140 (0) | USA | 2010 | [63] |
| Raw alfalfa sprouts | Salmonella Newport | 44 (0) | USA | 2010 | [64] |
| Romaine lettuce | E. coli O145 | 26 (0) | USA | 2010 | [65] |
| Raw alfalfa sprouts | Salmonella Saintpaul | 234 (0) | USA | 2009 | [66] |
| Raw produce | Salmonella Saintpaul | 565 (2) | USA | 2008 | [67] |
| Peppers | Salmonella | 1442 (2) | USA, Canada | 2008 | [68,69] |
| Cantaloupes | Salmonella Litchfield | 51 (0) | USA | 2008 | [70] |
| Lettuce | E. coli O157:H7 | 134 | USA, Canada | 2008 | [71] |
| Basil | Salmonella | 32 | UK | 2008 | [32] |
| Baby spinach | Salmonella | 354 | Europe | 2007 | [72] |
| Basil | Salmonella | 51 | North America, Europe | 2007 | [73] |
| Baby carrots | Shigella sonnei | 230 | Australia, Europe | 2007 | [74] |
| Alfalfa sprouts | Salmonella | 45 | Europe | 2007 | [75] |
| Tomatoes | Salmonella Typhimurium | 183 (0) | USA | 2006 | [76] |
| Fresh spinach | E. coli O157:H7 | 199 (3) | USA | 2006 | [77] |
| Alfalfa sprouts | Salmonella | 125 | Australia | 2006 | [78] |
| Cantaloupe | Salmonella | 115 | Australia | 2006 | [79] |
| Mung bean sprouts | Salmonella | 592 | Canada | 2005 | [80] |
| Tomatoes | Salmonella | 459 | USA | 2005 | [81] |
| Ambler Classification | Bush–Jacoby–Medeiros Classification | Distinctive Substrate | Inhibitor | Representative Enzyme |
|---|---|---|---|---|
| C | 1 | cephalosporins | none | AmpC |
| A | 2b | penicillins, early cephalosporins | beta-lactamase inhibitors | TEM-1, TEM-2, TEM-13, SHV-1 |
| A | 2be | extended-spectrum cephalosporins and aztreonam | beta-lactamase inhibitors | TEM-3, SHV-2, PER, VEB, CTX-M-15 |
| D | 2 | cloxacillin | beta-lactamase inhibitors | OXA-1, OXA-10 |
| D | 2de | extended-spectrum cephalosporins | beta-lactamase inhibitors | OXA-11, OXA-15 |
| D | 2df | carbapenems | beta-lactamaseinhibitors | OXA-23, OXA-48 |
| A | 2f | carbapenems | beta-lactamase inhibitors | KPC, IMI, SME, NMC |
| B | 3a | carbapenems | EDTA | MBL |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwu, C.D.; Okoh, A.I. Preharvest Transmission Routes of Fresh Produce Associated Bacterial Pathogens with Outbreak Potentials: A Review. Int. J. Environ. Res. Public Health 2019, 16, 4407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16224407
Iwu CD, Okoh AI. Preharvest Transmission Routes of Fresh Produce Associated Bacterial Pathogens with Outbreak Potentials: A Review. International Journal of Environmental Research and Public Health. 2019; 16(22):4407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16224407
Chicago/Turabian StyleIwu, Chidozie Declan, and Anthony Ifeanyi Okoh. 2019. "Preharvest Transmission Routes of Fresh Produce Associated Bacterial Pathogens with Outbreak Potentials: A Review" International Journal of Environmental Research and Public Health 16, no. 22: 4407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16224407