Rapid Biodegradation of the Organophosphorus Insecticide Chlorpyrifos by Cupriavidus nantongensis X1T


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Culture Medium
2.2. Inoculum Preparation
2.3. Degradation of Chlorpyrifos by Strain X1T
2.4. Determination of Chlorpyrifos and TCP
2.5. RT-qPCR Analysis of the Expression of Chlorpyrifos Degradation Gene
2.6. Determination of the Metabolites of Chlorpyrifos by Strain X1T
2.7. Degradation of Chlorpyrifos by Crude Enzyme
2.8. Calculation and Statistical Analysis
3. Results
3.1. Effects of Inoculation Volume on Chlorpyrifos Degradation by Strain X1T
3.2. Effects of pH on Chlorpyrifos Degradation by Strain X1T
3.3. Effects of Temperature on Chlorpyrifos Degradation by Strain X1T
3.4. Degradation Kinetics of Chlorpyrifos by Strain X1T
3.5. Epression of opdB by RT-qPCR
3.6. Metabolites and Degradation Pathways of Chlorpyrifos by Strain X1T
3.7. Degradation Activity of Crude Enzyme
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yadav, M.; Shukla, A.K.; Srivastva, N.; Upadhyay, S.N.; Dubey, S.K. Utilization of microbial community potential for removal of chlorpyrifos: A review. Crit. Rev. Biotechnol. 2016, 36, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Ge, J.; Li, Y.; Cheng, J.; Zhong, J.; Yu, X. Isolation, colonization, and chlorpyrifos degradation mediation of the endophytic bacterium Sphingomonas strain HJY in Chinese Chives (Allium tuberosum). J. Agric. Food Chem. 2017, 65, 1131–1138. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Singhasemanon, N.; Goh, K.S. A statistical assessment of pesticide pollution in surface waters using environmental monitoring data: Chlorpyrifos in Central Valley, California. Sci. Total Environ. 2016, 571, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Walker, A.; Morgan, J.A.W.; Wright, D.J. Effects of soil pH on the biodegradation of chlorpyrifos and isolation of a chlorpyrifos-degrading bacterium. Appl. Environ. Microbiol. 2003, 69, 5198–5206. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.W.; Gilliom, R.J.; Ryberg, K.R. Pesticides in U.S. streams and rivers: Occurrence and trends during 1992–2011. Environ. Sci. Technol. 2014, 48, 11025–11030. [Google Scholar] [CrossRef] [PubMed]
- Tuncel, S.G.; Oztas, N.B.; Erduran, M.S. Air and groundwater pollution in an agricultural region of the Turkish Mediterranean coast. J. Air Waste Manag. Assoc. 2008, 58, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.; Khan, Q.M.; Ali, R.; Ali, T.; Mobeen, A. Genotoxicity of chlorpyrifos in freshwater fish Labeo rohita using Alkaline Single-cell Gel Electrophoresis (Comet) assay. Drug Chem. Toxicol. 2014, 37, 466–471. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, J.; Wu, C.; Lv, S.; Lu, D.; Qi, X.; Jiang, S.; Feng, C.; Yu, H.; Liang, W.; et al. Associations of prenatal and childhood chlorpyrifos exposure with Neurodevelopment of 3-year-old children. Environ. Pollut. 2019, 251, 538–546. [Google Scholar] [CrossRef]
- Affam, A.C.; Chaudhuri, M. Degradation of pesticides chlorpyrifos, cypermethrin and chlorothalonil in aqueous solution by TiO2 photocatalysis. J. Environ. Manag. 2013, 130, 160–165. [Google Scholar] [CrossRef]
- Bootharaju, M.S.; Pradeep, T. Understanding the degradation pathway of the pesticide, chlorpyrifos by noble metal nanoparticles. Langmuir ACS J. Surf. Colloids 2012, 28, 2671–2679. [Google Scholar] [CrossRef]
- Kumar, S.; Kaushik, G.; Dar, M.A.; Nimesh, S.; Lopez-Chuken, U.J.; Villarreal-Chiu, J.F. Microbial degradation of organophosphate pesticides: A review. Pedosphere 2018, 28, 190–208. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, S.; Ma, C.; Wu, N.; Li, C.; Yang, X. Simultaneous biodegradation of bifenthrin and chlorpyrifos by Pseudomonas sp. CB2. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2018, 53, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wang, S.; Shi, Y.; Qu, J.; Zhai, Y.; Xu, L.; Xu, Y.; Song, J.; Liu, L.; Rahman, M.A.; et al. Genome sequence of Paracoccus sp. strain TRP, a chlorpyrifos biodegrader. J. Bacteriol. 2011, 193, 1786–1787. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, J.; Li, S. Isolation of a chlorpyrifos-degrading bacterium, Sphingomonas sp. strain Dsp-2, and cloning of the mpd gene. Res. Microbiol. 2007, 158, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Li, Q.; Liu, H.; Feng, Z.; Yan, X.; Hong, Q.; Li, S. Biodegradation of chlorpyrifos and 3,5,6-trichloro-2-pyridinol by Cupriavidus sp. DT-1. Bioresour. Technol. 2013, 127, 337–342. [Google Scholar] [CrossRef]
- Abraham, J.; Silambarasan, S. Biodegradation of chlorpyrifos and its hydrolysis product 3,5,6-trichloro-2-pyridinol using a novel bacterium Ochrobactrum sp. JAS2: A proposal of its metabolic pathway. Pestic. Biochem. Physiol. 2016, 126, 13–21. [Google Scholar] [CrossRef]
- Lei, W.J.; Zhou, X.Y. Experiment and simulation on adsorption of 3,5,6-trichloro-2-pyridinol in typical farmland of purple soil, Southwestern China. Soil Sediment Contam. 2017, 26, 345–363. [Google Scholar] [CrossRef]
- Dores, E.F.G.C.; Spadotto, C.A.; Weber, O.L.S.; Villa, R.D.; Vecchiato, A.B.; Pinto, A.A. Environmental behavior of chlorpyrifos and endosulfan in a tropical soil in Central Brazil. J. Agric. Food Chem. 2016, 64, 3942–3948. [Google Scholar] [CrossRef]
- Caceres, T.; He, W.X.; Naidu, R.; Meharaj, M. Toxicity of chlorpyrifos and TCP alone and in combination to Daphnia carinata: The influence of microbial degradation in natural water. Water Res. 2007, 41, 4497–4503. [Google Scholar] [CrossRef]
- Fang, L.; Shi, T.; Chen, Y.; Wu, X.; Zhang, C.; Tang, X.; Li, Q.X.; Hua, R. Kinetics and catabolic pathways of the insecticide chlorpyrifos, annotation of the degradation genes, and characterization of enzymes TcpA and Fre in Cupriavidus nantongensis X1T. J. Agric. Food Chem. 2019, 67, 2245–2254. [Google Scholar] [CrossRef]
- Feng, Y.; Racke, K.D.; Bollag, J. Isolation and characterization of a chlorinated-pyridinol-degrading bacterium. Appl. Environ. Microbiol. 1997, 63, 4096–4098. [Google Scholar]
- Cao, L.; Xu, J.H.; Wu, G.; Li, M.X.; Jiang, J.D.; He, J.; Li, S.P.; Hong, Q. Identification of two combined genes responsible for dechlorination of 3,5,6-trichloro-2-pyridinol (TCP) in Cupriavidus pauculus P2. J. Hazard. Mater. 2013, 260, 700–706. [Google Scholar] [CrossRef]
- Anspaugh, D.D.; Roe, R.M. Assays for the classification of two types of esterases: Carboxylic ester hydrolases and phosphoric triester hydrolases. Curr. Protoc. Toxicol. 2002, 13, 4–10. [Google Scholar]
- van Dyk, J.S.; Pletschke, B. Review on the use of enzymes for the detection of organochlorine, organophosphate and carbamate pesticides in the environment. Chemosphere 2011, 82, 291–307. [Google Scholar] [CrossRef]
- Schenk, G.; Mateen, I.; Ng, T.K.; Pedroso, M.M.; Mitic, N.; Jafelicci, M.; Marques, R.F.C.; Gahan, L.R.; Ollis, D.L. Organophosphate-degrading metallohydrolases: Structure and function of potent catalysts for applications in bioremediation. Coord. Chem. Rev. 2016, 317, 122–131. [Google Scholar] [CrossRef]
- Fang, L.C.; Chen, Y.F.; Zhou, Y.L.; Wang, D.S.; Sun, L.N.; Tang, X.Y.; Hua, R.M. Complete genome sequence of a novel chlorpyrifos degrading bacterium, Cupriavidus nantongensis X1. J. Biotechnol. 2016, 227, 1–2. [Google Scholar] [CrossRef]
- Sun, L.-N.; Wang, D.-S.; Yang, E.-D.; Fang, L.-C.; Chen, Y.-F.; Tang, X.-Y.; Hua, R.-M. Cupriavidus nantongensis sp. nov., a novel chlorpyrifos-degrading bacterium isolated from sludge. Int. J. Syst. Evol. Microbiol. 2016, 66, 2335–2341. [Google Scholar] [CrossRef]
- Chen, S.; Liu, C.; Peng, C.; Liu, H.; Hu, M.; Zhong, G. Biodegradation of chlorpyrifos and its hydrolysis product 3,5,6-trichloro-2-pyridinol by a new fungal strain Cladosporium cladosporioides Hu-01. PLoS ONE 2012, 7, e47205. [Google Scholar] [CrossRef]
- Anwar, S.; Liaquat, F.; Khan, Q.M.; Khalid, Z.M.; Iqbal, S. Biodegradation of chlorpyrifos and its hydrolysis product 3,5,6-trichloro-2-pyridinol by Bacillus pumilus strain C2A1. J. Hazard. Mater. 2009, 168, 400–405. [Google Scholar] [CrossRef]
- Gilani, R.A.; Rafique, M.; Rehman, A.; Munis, M.F.H.; Rehman, S.U.; Chaudhary, H.J. Biodegradation of chlorpyrifos by bacterial genus Pseudomonas. J. Basic Microbiol. 2016, 56, 105–119. [Google Scholar] [CrossRef]
- Singh, D.P.; Nadda, J.; Singh, Y.; Garg, A.; Kaur, N.; Gulati, A. Chlorpyrifos degradation by the cyanobacterium Synechocystis sp. strain PUPCCC 64. Environ. Sci. Pollut. Res. 2011, 18, 1351–1359. [Google Scholar] [CrossRef]
- Mallick, K.; Bharati, K.; Banerji, A.; Shakil, N.A.; Sethunathan, N. Bacterial degradation of chlorpyrifos in pure cultures and in soil. Bull. Environ. Contam. Toxicol. 1999, 62, 48–54. [Google Scholar] [CrossRef]
- Duraisamy, K.; Muthusamy, S.; Balakrishnan, S. An eco-friendly detoxification of chlorpyrifos by Bacillus cereus MCAS02 native isolate from agricultural soil, Namakkal, Tamil Nadu, India. Biocatal. Agric. Biotechnol. 2018, 13, 283–290. [Google Scholar] [CrossRef]
- Briceño, G.; Fuentes, M.S.; Palma, G.; Jorquera, M.A.; Amoroso, M.J.; Diez, M.C. Chlorpyrifos biodegradation and 3,5,6-trichloro-2-pyridinol production byactinobacteria isolated from soil. Int. Biodeterior. Biodegrad. 2012, 73, 1–7. [Google Scholar] [CrossRef]
- Xu, G.; Zheng, W.; Li, Y.; Wang, S.; Zhang, J.; Yan, Y. Biodegradation of chlorpyrifos and 3,5,6-trichloro-2-pyridinol by a newly isolated Paracoccus sp. strain TRP. Int. Biodeter. Biodegr. 2008, 62, 51–56. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, Y.H.; Zhang, B.X.; Yang, C.H.; Zhang, X. Isolation and characterization of a chlorpyrifos and 3,5,6-trichloro-2-pyridinol degrading bacterium. FEMS Microbiol. Lett. 2010, 251, 67–73. [Google Scholar] [CrossRef]
- Abraham, J.; Silambarasan, S. Biodegradation of chlorpyrifos and its hydrolyzing metabolite 3,5,6-trichloro-2-pyridinol by Sphingobacterium sp. JAS3. Process Biochem. 2013, 48, 1559–1564. [Google Scholar] [CrossRef]
- Silambarasan, S.; Abraham, J. Kinetic studies on enhancement of degradation of chlorpyrifos and its hydrolyzing metabolite TCP by a newly isolated Alcaligenes sp. JAS1. J. Taiwan Inst. Chem. Eng. 2013, 44, 438–445. [Google Scholar] [CrossRef]
- Yang, L.L.; Yang, X.; Li, G.B.; Fan, K.G.; Yin, P.F.; Chen, X.G. An integrated molecular docking and rescoring method for predicting the sensitivity spectrum of various serine hydrolases to organophosphorus pesticides. J. Sci. Food Agric. 2016, 96, 2184–2192. [Google Scholar] [CrossRef]
- Li, Y.; Yang, H.; Xu, F. Identifying and engineering a critical amino acid residue to enhance the catalytic efficiency of Pseudomonas sp. methyl parathion hydrolase. Appl. Microbiol. Biotechnol. 2018, 102, 6537–6545. [Google Scholar] [CrossRef]
- Pedroso, M.M.; Ely, F.; Mitic, N.; Carpenter, M.C.; Gahan, L.R.; Wilcox, D.E.; Larrabee, J.L.; Ollis, D.L.; Schenk, G. Comparative investigation of the reaction mechanisms of the organophosphate-degrading phosphotriesterases from Agrobacterium radiobacter (OpdA) and Pseudomonas diminuta (OPH). J. Biol. Inorg. Chem. 2014, 19, 1263–1275. [Google Scholar] [CrossRef]
- Sharma, G.; Hu, Q.Y.; Jayasinghe-Arachchige, V.M.; Paul, T.J.; Schenk, G.; Prabhakar, R. Investigating coordination flexibility of glycerophosphodiesterase (GpdQ) through interactions with mono-, di-, and triphosphoester (NPP, BNPP, GPE, and paraoxon) substrates. Phys. Chem. Chem. Phys. 2019, 21, 5499–5509. [Google Scholar] [CrossRef]
- Singh, B.K.; Walker, A.; Morgan, J.A.; Wright, D.J. Biodegradation of chlorpyrifos by enterobacter strain B-14 and its use in bioremediation of contaminated soils. Appl. Environ. Microbiol. 2004, 70, 4855–4863. [Google Scholar] [CrossRef] [Green Version]
- Cortes-Eslava, J.; Gomez-Arroyo, S.; Risueno, M.C.; Testillano, P.S. The effects of organophosphorus insecticides and heavy metals on DNA damage and programmed cell death in two plant models. Environ. Pollut. 2018, 240, 77–86. [Google Scholar] [CrossRef]
- Musilek, K.; Dolezal, M.; Gunn-Moore, F.; Kuca, K. Design, evaluation and structure-activity relationship studies of the AChE reactivators against organophosphorus pesticides. Med. Res. Rev. 2011, 31, 548–575. [Google Scholar] [CrossRef]
- Jiang, X.L.; Li, M.; Zhou, J.G.; Yang, Q.B.; Du, L.J.; Du, J. Plasma paraoxonase-1, oxidized low-density lipoprotein and lipid peroxidation levels in gout patients. Cell Biochem. Biophys. 2011, 61, 461–466. [Google Scholar] [CrossRef]
- Bae, S.Y.; Myslinski, J.M.; McMahon, L.R.; Height, J.J.; Bigley, A.N.; Raushel, F.M.; Harvey, S.P. An OPAA enzyme mutant with increased catalytic efficiency on the nerve agents sarin, soman, and GP. Enzym. Microb. Technol. 2018, 112, 65–71. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (mg/L) | Degradation Kinetics Equation | k | Half-Life T1/2 (h) |
|---|---|---|---|---|
| Chlorpyrifos | 20 | Ct = 20.49e−0.308t R2 = 0.9612 | 0.308 | 2.25 |
| 200 | Ct = 238.02e−0.071t R2 = 0.9003 | 0.071 | 9.76 | |
| 500 | Ct = 568.20e−0.044t R2 = 0.9795 | 0.044 | 15.75 |
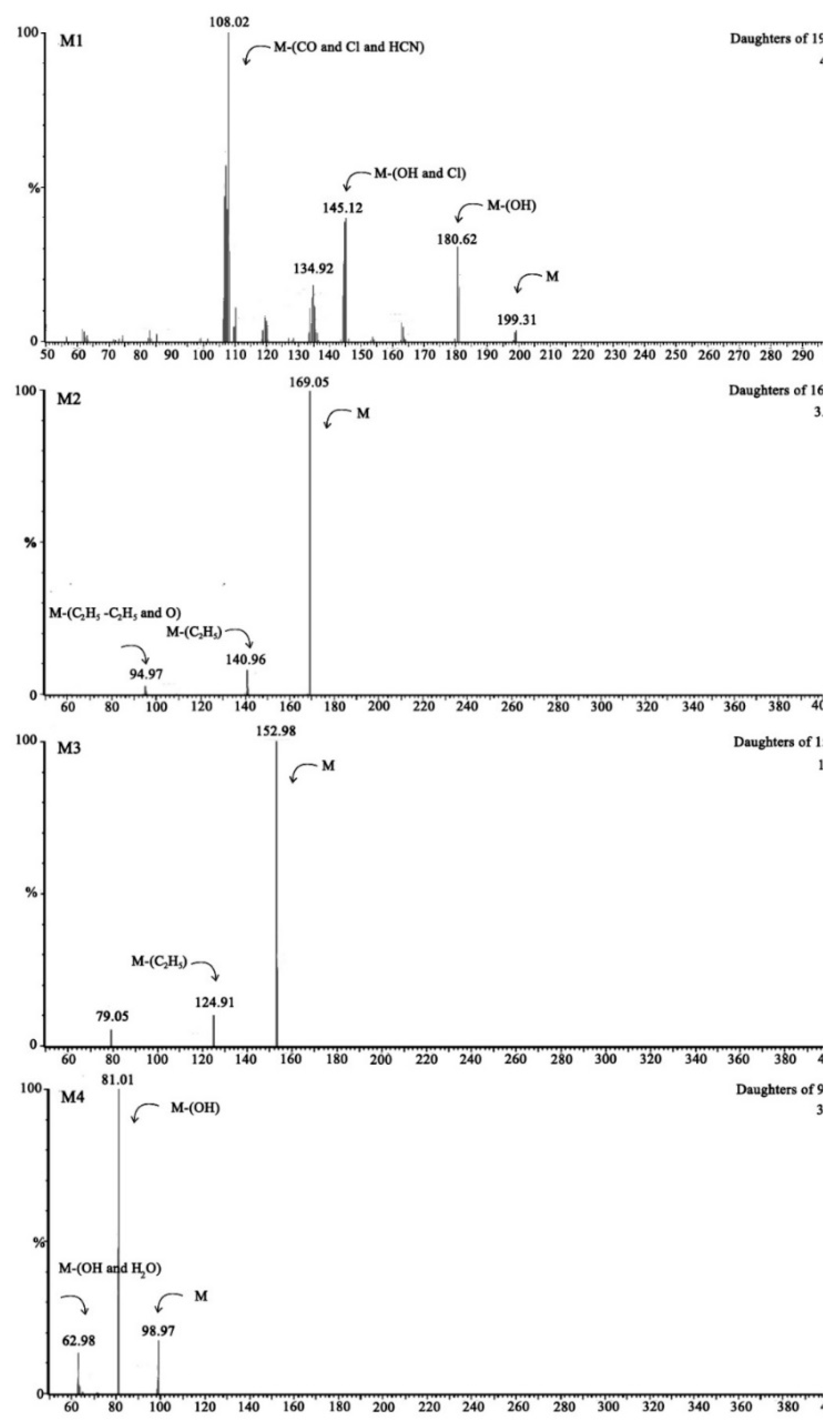
| Metabolites | Compounds and Molecular Weight (amu) | Molecular Ions (m/z) | Major Fragments (m/z) | |
|---|---|---|---|---|
| M1 | TCP | 198.43 | 199.31 | 181[M-OH], 145[M-Cl-OH], 108[M-Cl-CO-HCN] |
| M2 | DETP | 170.17 | 169.05 | 141[M-C2H5], 95[M-C2H5-C2H5-O] |
| M3 | DEP | 154.10 | 152.98 | 125[M-C2H5] |
| M4 | H3PO4 | 98.00 | 98.97 | 81[M-OH],63[M-OH-H2O] |
| M5 | DCPD | 180.00 | 178.85 | 163[M-O],136[M-CHON],108[M-2Cl] |
| M6 | DHPD | 141.08 | 140.29 | 112[M-CO], 84[M-C2O2], 68[M-C2O2-O] |
| M7 | ATOPA | 159.10 | 159.06 | 144[M-NH2],116[M-NH2-CO], |
| Strains | Initial Concentration (mg·L−1) | Time (day) | Degradation (%) | References |
|---|---|---|---|---|
| C. nantongensis.X1T | 20 | 0.67 | 100 | This study |
| C. nantongensis.X1T | 200 | 2 | 100 | This study |
| C. nantongensis.X1T | 500 | 2 | 90 | This study |
| Synechocystis sp. PUPCCC 64 | 5 | 5 | 93.8 | [31] |
| Flavobacterium sp.TCC27551 | 10 | 2 | 100 | [32] |
| Enterobacter B-14 | 20 | 3 | 100 | [28] |
| C. cladosporioides Hu-01 | 50 | 5 | 100 | [33] |
| B. cereus MCAS02 | 50 | 7 | 89 | [34] |
| Streptomyces sp. AC5 | 50 | 1 | 90 | [35] |
| Paracoccus sp. TRP | 50 | 4 | 100 | [36] |
| A. faecalisDSP3 | 100 | 12 | 100 | [28] |
| Sphingobacterium sp. JAS3 | 300 | 5 | 100 | [37] |
| Alcaligenes sp. JAS1 | 300 | 3 | 100 | [38] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, T.; Fang, L.; Qin, H.; Chen, Y.; Wu, X.; Hua, R. Rapid Biodegradation of the Organophosphorus Insecticide Chlorpyrifos by Cupriavidus nantongensis X1T. Int. J. Environ. Res. Public Health 2019, 16, 4593. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16234593
Shi T, Fang L, Qin H, Chen Y, Wu X, Hua R. Rapid Biodegradation of the Organophosphorus Insecticide Chlorpyrifos by Cupriavidus nantongensis X1T. International Journal of Environmental Research and Public Health. 2019; 16(23):4593. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16234593
Chicago/Turabian StyleShi, Taozhong, Liancheng Fang, Han Qin, Yifei Chen, Xiangwei Wu, and Rimao Hua. 2019. "Rapid Biodegradation of the Organophosphorus Insecticide Chlorpyrifos by Cupriavidus nantongensis X1T" International Journal of Environmental Research and Public Health 16, no. 23: 4593. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16234593