Effects of Nanoplastics on Freshwater Biofilm Microbial Metabolic Functions as Determined by BIOLOG ECO Microplates

Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of Nanoplastics
2.2. Biofilm Cultivation
2.3. Ecotoxicity Experiment
2.4. Community-Level Physiological Profiling
3. Results and Discussion
3.1. Characterization of PS Beads
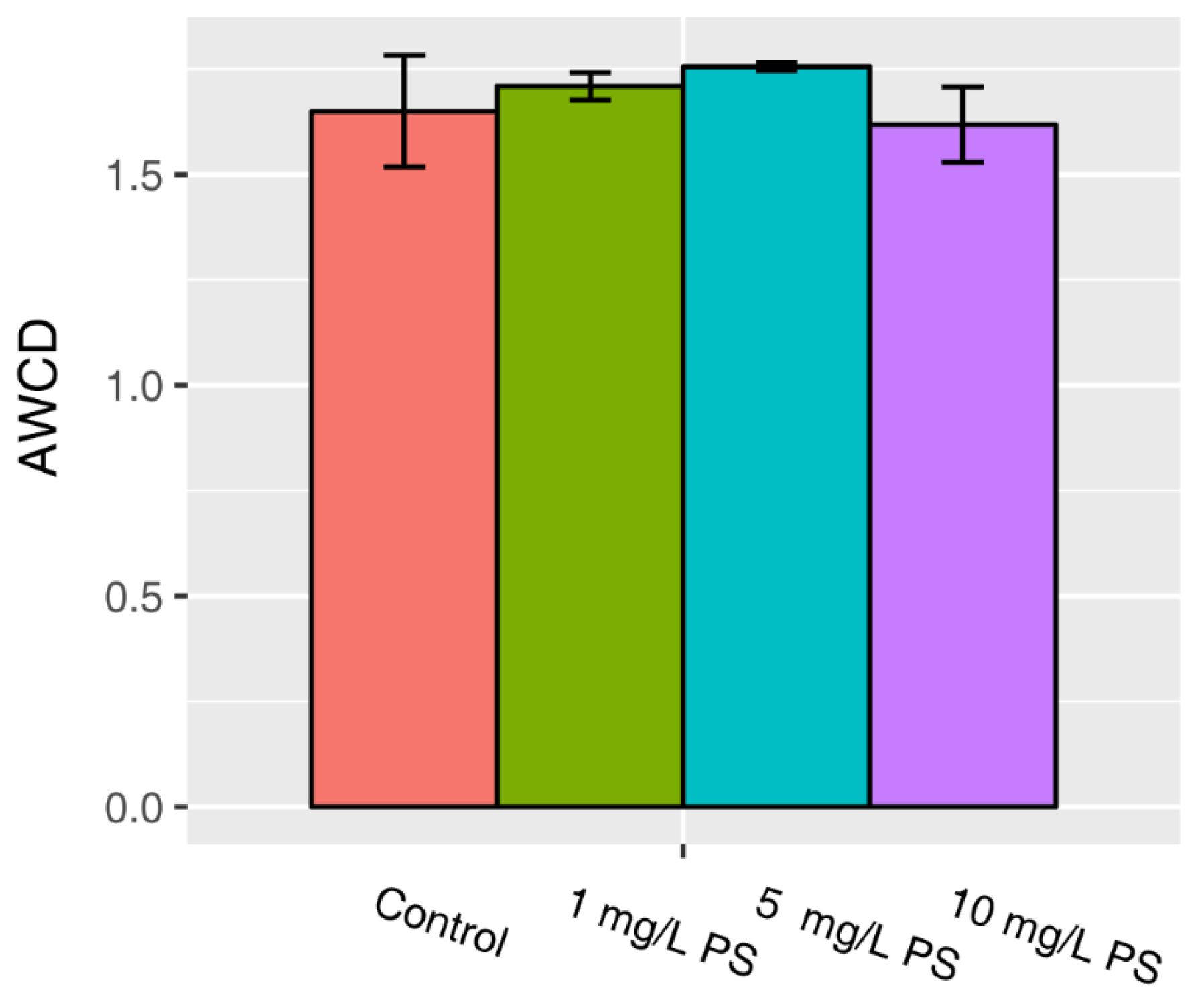
3.2. AWCD of All Carbon Sources
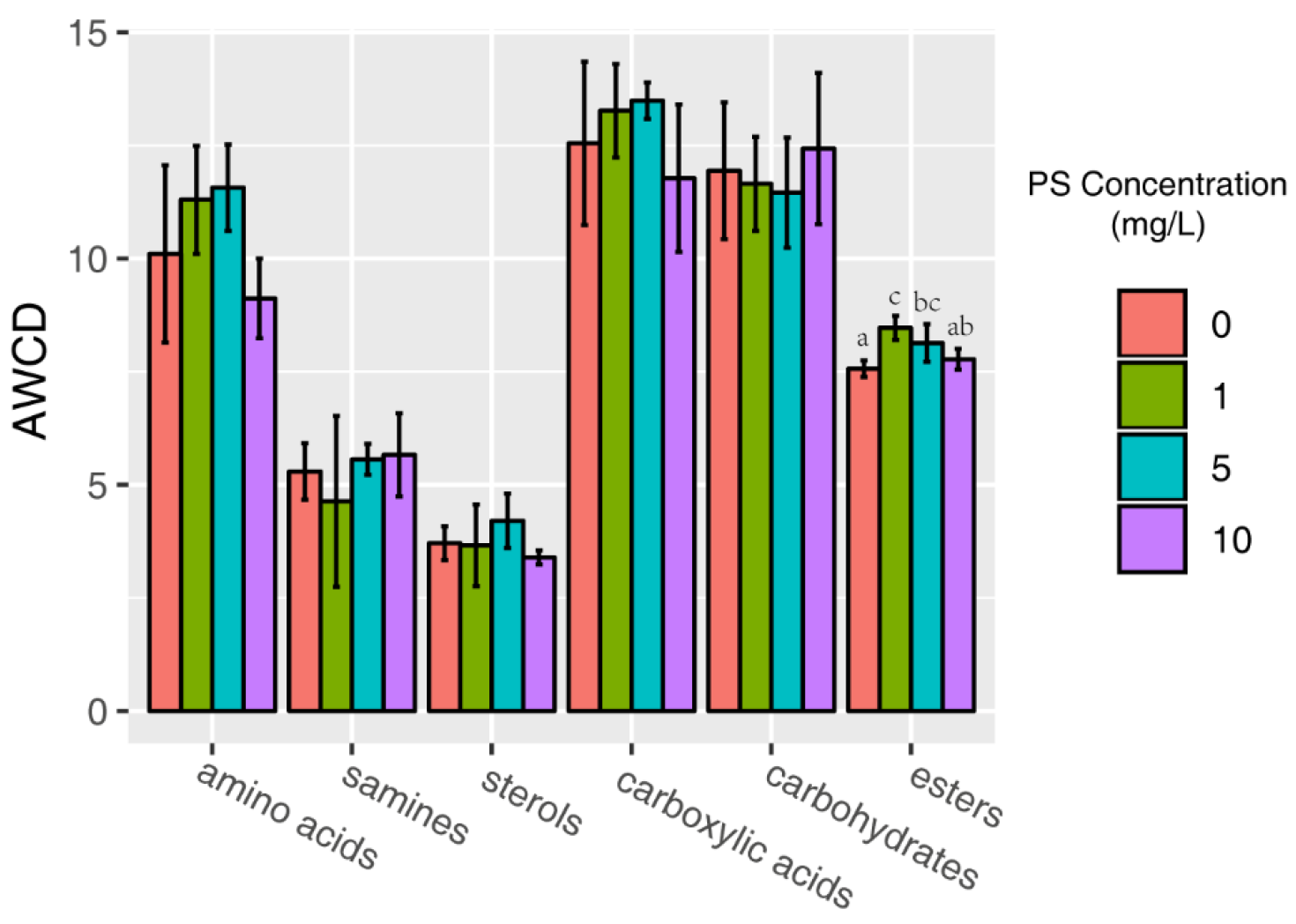
3.3. AWCD by Biochemical Categories
3.4. AWCD by Specific Carbon Source
3.5. Microbial Metabolic Functional Diversity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Plastics Europe Market Research Group (PEMRG) and Conversio Market & Strategy GmbH. Plastics-the Facts 2019; Plastics Europe Market Research Group (PEMRG) and Conversio Market & Strategy GmbH: Mainz, Germany, 2019; p. 14. [Google Scholar]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3. [Google Scholar] [CrossRef] [PubMed]
- Alimi, O.S.; Budarz, J.F.; Hernandez, L.M.; Tufenkji, N. Microplastics and Nanoplastics in Aquatic Environments: Aggregation, Deposition, and Enhanced Contaminant Transport. Environ. Sci. Technol. 2018, 52, 1704–1724. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, J.P.; Santos, P.S.M.; Duarte, A.C.; Rocha-Santos, T. (Nano)plastics in the environment—Sources, fates and effects. Sci. Total. Environ. 2016, 566, 15–26. [Google Scholar] [CrossRef]
- Sharma, S.; Chatterjee, S. Microplastic pollution, a threat to marine ecosystem and human health: A short review. Environ. Sci. Pollut. R. 2017, 24, 21530–21547. [Google Scholar] [CrossRef] [PubMed]
- Shim, W.J.; Hong, S.H.; Eo, S.E. Identification methods in microplastic analysis: A review. Anal. Methods. UK 2017, 9, 1384–1391. [Google Scholar] [CrossRef]
- Triebskorn, R.; Braunbeck, T.; Grummt, T.; Hanslik, L.; Huppertsberg, S.; Jekel, M.; Knepper, T.P.; Krais, S.; Muller, Y.K.; Pittroff, M.; et al. Relevance of nano- and microplastics for freshwater ecosystems: A critical review. Trac-Trend Anal. Chem. 2019, 110, 375–392. [Google Scholar] [CrossRef]
- Ogonowski, M.; Motiei, A.; Ininbergs, K.; Hell, E.; Gerdes, Z.; Udekwu, K.I.; Bacsik, Z.; Gorokhova, E. Evidence for selective bacterial community structuring on microplastics. Environ. Microbiol. 2018, 20, 2796–2808. [Google Scholar] [CrossRef]
- Rochman, C.M.; Browne, M.A.; Underwood, A.J.; van Franeker, J.A.; Hompson, R.C.T.; Amaral-Zettler, L.A. The ecological impacts of marine debris: Unraveling the demonstrated evidence from what is perceived. Ecology 2016, 97, 302–312. [Google Scholar] [CrossRef]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R. Accumulation of Microplastic on Shorelines Woldwide: Sources and Sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef]
- Sussarellu, R.; Suquet, M.; Thomas, Y.; Lambert, C.; Fabioux, C.; Pernet, M.E.J.; Le Goic, N.; Quillien, V.; Mingant, C.; Epelboin, Y.; et al. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. USA 2016, 113, 2430–2435. [Google Scholar] [CrossRef]
- Farrell, P.; Nelson, K. Trophic level transfer of microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ. Pollut. 2013, 177, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Nienhaus, K.; Nienhaus, G.U. Engineered nanoparticles interacting with cells: Size matters. J. Nanobiotechnol. 2014, 12. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Loder, M.G.J.; Labrenz, M. Marine microplastic- associated biofilms—A review. Environ. Chem. 2015, 12, 551–562. [Google Scholar] [CrossRef]
- Forte, M.; Iachetta, G.; Tussellino, M.; Carotenuto, R.; Prisco, M.; De Falco, M.; Laforgia, V.; Valiante, S. Polystyrene nanoparticles internalization in human gastric adenocarcinoma cells. Toxicol. In Vitro 2016, 31, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.F.; Ai, H.N.; Chen, Y.; Zhang, Z.Y.; Zeng, P.; Kang, L.; Li, W.; Gu, W.K.; He, Q.; Li, H. Phytoplankton response to polystyrene microplastics: Perspective from an entire growth period. Chemosphere 2018, 208, 59–68. [Google Scholar] [CrossRef]
- Besseling, E.; Wang, B.; Lurling, M.; Koelmans, A.A. Nanoplastic Affects Growth of S. obliquus and Reproduction of D. magna. Environ. Sci. Technol. 2014, 48, 14065. [Google Scholar] [CrossRef]
- Sun, X.M.; Chen, B.J.; Li, Q.F.; Liu, N.; Xia, B.; Zhu, L.; Qu, K.M. Toxicities of polystyrene nano- and microplastics toward marine bacterium Halomonas alkaliphila. Sci. Total. Environ. 2018, 642, 1378–1385. [Google Scholar] [CrossRef]
- Besseling, E.; Quik, J.T.K.; Sun, M.; Koelmans, A.A. Fate of nano- and microplastic in freshwater systems: A modeling study. Environ. Pollut. 2017, 220, 540–548. [Google Scholar] [CrossRef]
- Battin, T.J.; Besemer, K.; Bengtsson, M.M.; Romani, A.M.; Packmann, A.I. The ecology and biogeochemistry of stream biofilms. Nat. Rev. Microbiol. 2016, 14, 251–263. [Google Scholar] [CrossRef]
- Battin, T.J.; Kaplan, L.A.; Newbold, J.D.; Hansen, C.M.E. Contributions of microbial biofilms to ecosystem processes in stream mesocosms. Nature 2003, 426, 439–442. [Google Scholar] [CrossRef]
- Wu, Y.; He, J.; Yang, L. Evaluating adsorption and biodegradation mechanisms during the removal of microcystin-RR by periphyton. Environ. Sci. Technol. 2010, 44, 6319–6324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.S.; Zhang, S.H.; Lv, X.Y.; Qiu, Z.; Zhang, Z.Q.; Yan, L.Y. Dissolved organic matter release in overlying water and bacterial community shifts in biofilm during the decomposition of Myriophyllum verticillatum. Sci. Total. Environ. 2018, 633, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Price, O.R.; van der Gast, C.J.; Finnegan, C.J.; van Egmond, R.A.; Schafer, H.; Bending, G.D. Spatial and temporal variability in the potential of river water biofilms to degrade p-nitrophenol. Chemosphere 2016, 164, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.Z.; Wang, P.F.; Hou, J.; Wang, C.; Yao, Y. Research progress on toxicity of metallic nanomaterials to different microbial aggregates. Water Resour. Prot. 2019, 94, 73–78. (In Chinese) [Google Scholar]
- Miao, L.Z.; Wang, P.F.; Wang, C.; Hou, J.; Yao, Y.; Liu, J.; Lv, B.W.; Yang, Y.Y.; You, G.X.; Xu, Y.; et al. Effect of TiO2 and CeO2 nanoparticles on the metabolic activity of surficial sediment microbial communities based on oxygen microelectrodes and high-throughput sequencing. Water. Res. 2018, 129, 287–296. [Google Scholar] [CrossRef]
- Wang, C.; Miao, L.Z.; Hou, J.; Wang, P.F.; Qian, J.; Dai, S.S. The effect of flow velocity on the distribution and composition of extracellular polymeric substances in biofilms and the detachment mechanism of biofilms. Water Sci. Technol. 2014, 69, 825–832. [Google Scholar] [CrossRef]
- Miao, L.; Hou, J.; You, G.; Liu, Z.; Liu, S.; Li, T.; Mo, Y.; Guo, S.; Qu, H. Acute effects of nanoplastics and microplastics on periphytic biofilms depending on particle size, concentration and surface modification. Environ. Pollut. 2019, 255, 113300. [Google Scholar] [CrossRef]
- Qu, M.; Nida, A.; Kong, Y.; Du, H.H.; Xiao, G.S.; Wang, D.Y. Nanopolystyrene at predicted environmental concentration enhances microcystin-LR toxicity by inducing intestinal damage in Caenorhabditis elegans. Ecotox. Environ. Saf. 2019, 183. [Google Scholar] [CrossRef]
- Liu, Y.H.; Wang, Z.; Wang, S.; Fang, H.; Ye, N.; Wang, D.G. Ecotoxicological effects on Scenedesmus obliquus and Danio rerio Co-exposed to polystyrene nano-plastic particles and natural acidic organic polymer. Environ. Toxicol. Phar. 2019, 67, 21–28. [Google Scholar] [CrossRef]
- Zhu, N.Y.; Wang, S.C.; Tang, C.L.; Duan, P.F.; Yao, L.G.; Tang, J.; Wong, P.K.; An, T.C.; Dionysiou, D.D.; Wu, Y.H. Protection Mechanisms of Periphytic Biofilm to Photocatalytic Nanoparticle Exposure. Environ. Sci. Technol. 2019, 53, 1585–1594. [Google Scholar] [CrossRef]
- Tang, J.; Zhu, N.Y.; Zhu, Y.; Liu, J.Z.; Wu, C.X.; Kerr, P.; Wu, Y.H.; Lam, P.K.S. Responses of Periphyton to Fe2O3 Nanoparticles: A Physiological and Ecological Basis for Defending Nanotoxicity. Environ. Sci. Technol. 2017, 51, 10797–10805. [Google Scholar] [CrossRef] [PubMed]
- Rutgers, M.; Wouterse, M.; Drost, S.M.; Breure, A.M.; Mulder, C.; Stone, D.; Creamer, R.E.; Winding, A.; Bloem, J. Monitoring soil bacteria with community-level physiological profiles using Biolog (TM) ECO-plates in the Netherlands and Europe. Appl. Soil. Ecol. 2016, 97, 23–35. [Google Scholar] [CrossRef]
- Garland, J.L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil. Biol. Biochem. 1996, 28, 213–221. [Google Scholar] [CrossRef]
- Ge, Z.W.; Du, H.J.; Gao, Y.L.; Qiu, W.F. Analysis on Metabolic Functions of Stored Rice Microbial Communities by BIOLOG ECO Microplates. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Button, M.; Nivala, J.; Weber, K.P.; Aubron, T.; Muller, R.A. Microbial community metabolic function in subsurface flow constructed wetlands of different designs. Ecol. Eng. 2015, 80, 162–171. [Google Scholar] [CrossRef]
- Xu, H.; Xu, M.; Li, Y.; Liu, X.; Guo, L.; Jiang, H. Characterization, origin and aggregation behavior of colloids in eutrophic shallow lake. Water Res. 2018, 142, 176–186. [Google Scholar] [CrossRef]
- Su, L.; Xue, Y.G.; Li, L.Y.; Yang, D.Q.; Kolandhasamy, P.; Li, D.J.; Shi, H.H. Microplastics in Taihu Lake, China. Environ. Pollut. 2016, 216, 711–719. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Wu, Y.H.; Zhuang, L.L.; Wang, X.X.; Hu, H.Y. Screening heterotrophic microalgal strains by using the Biolog method for biofuel production from organic wastewater. Algal. Res. 2014, 6, 175–179. [Google Scholar] [CrossRef]
- Kong, X.; Wang, C.; Ji, M. Analysis of microbial metabolic characteristics in mesophilic and thermophilic biofilters using Biolog plate technique. Chem. Eng. J. 2013, 230, 415–421. [Google Scholar] [CrossRef]
- Zhifei, L.I.; Guangjun, W.; Jun, X.; Ermeng, Y.; Deguang, Y.; Yun, X.; Nan, W. Microbial carbon metabolic characteristics of biofilm communities in the grass carp culture pond based on Biolog-ECO plates. J. Fish. China 2014, 38, 1985–1995. (In Chinese) [Google Scholar]
- Konopka, J.B. N-acetylglucosamine (GlcNAc) functions in cell signaling. Scientifica (Cairo) 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Wu, Y.H.; Esquivel-Elizondo, S.; Sorensen, S.J.; Rittmann, B.E. How Microbial Aggregates Protect against Nanoparticle Toxicity. Trends Biotechnol. 2018, 36, 1171–1182. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Particle Size | Particle Size Distribution (nm) | Zeta Potential (mV) | ||
|---|---|---|---|---|
| Milli-Q Water | Experimental Solution a | Milli-Q Water | Experimental Solution | |
| 100 nm | 129 ± 34 | 569 ± 124 * | −39.4 ± 3.9 | −19.4 ± 3.5 * |
| PS Concentration | H′ | D | E | D |
|---|---|---|---|---|
| 0 mg/L | 3.36 ± 0.012a | 0.964 ± 0.001a | 2.275 ± 0.008a | 0.838 ± 0.028a |
| 1 mg/L | 3.347 ± 0.017a | 0.963 ± 0.001a | 2.281 ± 0.01a | 0.82 ± 0.012a |
| 5 mg/L | 3.376 ± 0.009a | 0.964 ± 0a | 2.271 ± 0.01a | 0.815 ± 0.003a |
| 10 mg/L | 3.318 ± 0.041b | 0.961 ± 0.002b | 2.239 ± 0.021b | 0.832 ± 0.01a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, L.; Guo, S.; Liu, Z.; Liu, S.; You, G.; Qu, H.; Hou, J. Effects of Nanoplastics on Freshwater Biofilm Microbial Metabolic Functions as Determined by BIOLOG ECO Microplates. Int. J. Environ. Res. Public Health 2019, 16, 4639. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16234639
Miao L, Guo S, Liu Z, Liu S, You G, Qu H, Hou J. Effects of Nanoplastics on Freshwater Biofilm Microbial Metabolic Functions as Determined by BIOLOG ECO Microplates. International Journal of Environmental Research and Public Health. 2019; 16(23):4639. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16234639
Chicago/Turabian StyleMiao, Lingzhan, Song Guo, Zhilin Liu, Songqi Liu, Guoxiang You, Hao Qu, and Jun Hou. 2019. "Effects of Nanoplastics on Freshwater Biofilm Microbial Metabolic Functions as Determined by BIOLOG ECO Microplates" International Journal of Environmental Research and Public Health 16, no. 23: 4639. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16234639