The Bioaccumulation and Biodegradation of Testosterone by Chlorella vulgaris


Abstract
:1. Introduction
2. Materials and Methods
2.1. Algae and Culture Condition
2.2. Experimental Design
2.3. Algal Density
2.4. Dry Weight of Algal Cell
2.5. Determination of Testosterone in Algal Cells
2.6. Determination of Testosterone in Water
2.7. HPLC-MS Conditions
2.8. Statistical Analysis
3. Results
3.1. Algal Cell Growth
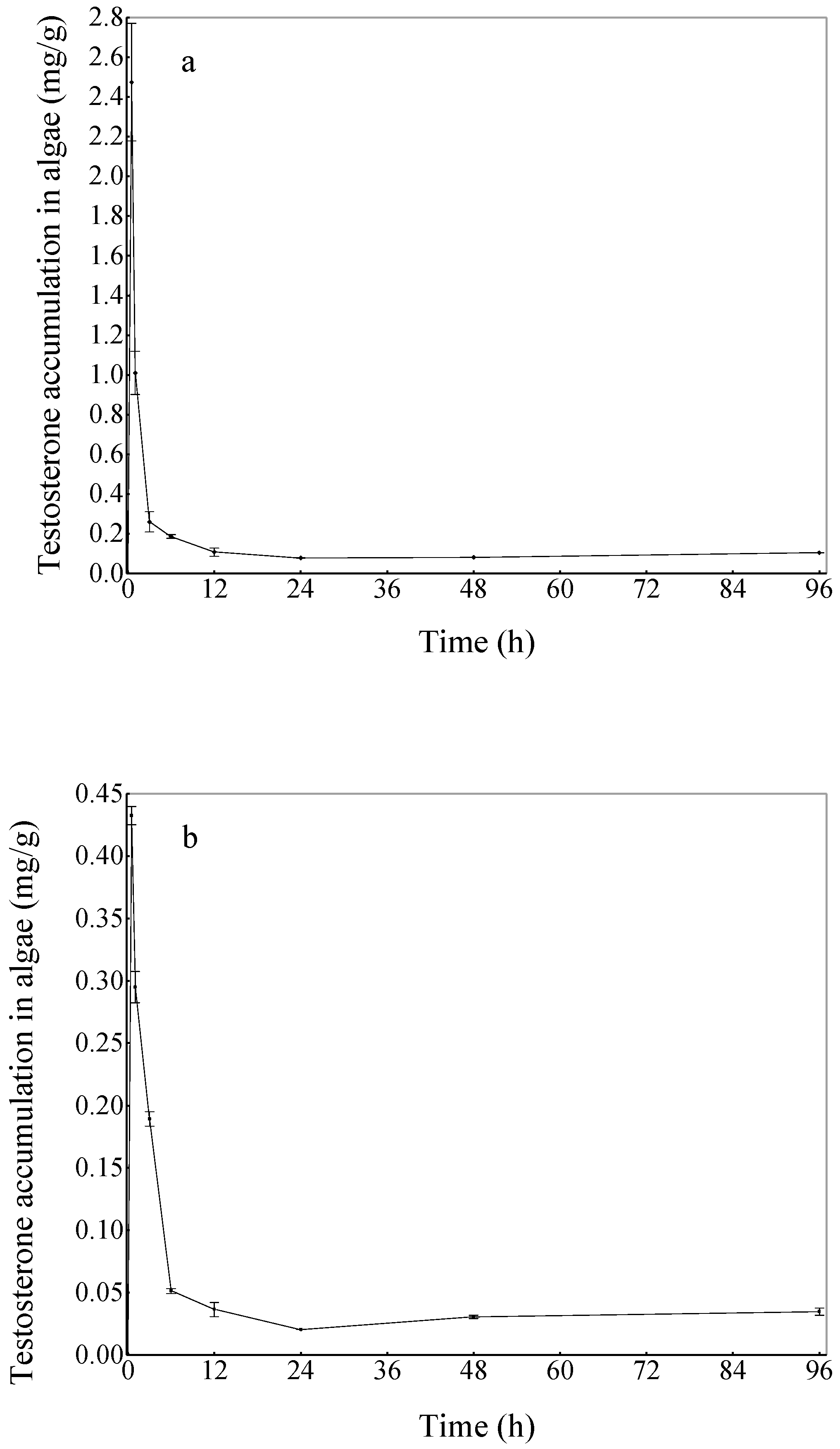
3.2. Accumulation of Testosterone in Chlorella Vulgaris
3.3. Degradation of Testosterone by Chlorella Vulgaris
3.4. The Kinetics Equation of Degradation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, L.S.; Carmosini, N.; Sassman, S.A.; Dion, H.M.; Sepúlveda, M.S. Agricultural contributions of antimicrobials and hormones on soils and water quanlity. Adv. Agron. 2007, 93, 1–68. [Google Scholar]
- Zheng, W.; Yates, S.R.; Bradford, S.A. Analysis of steroid hormones in a typical dairy waste disposal system. Environ. Sci. Technol. 2008, 42, 530–535. [Google Scholar] [CrossRef]
- Seki, M.; Yokota, H.; Matsubara, H.; Maeda, M.; Tadokoro, H.; Kobayashi, K. Fish full life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes). Environ. Toxicol. Chem. 2004, 23, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.B.; Morrison, J.; Galvez, J.I. Reproductive characteristics of adult channel catfish treated with trenbolone acetate during the phenocritical period of sex differentiation. Aquaculture 2000, 189, 351–360. [Google Scholar] [CrossRef]
- Barbosa, I.R.; Nogueira, A.J.A.; Soares, A.M.V.M. Acute and chronic effects of testosterone and 4-hydroxyandrostenedione to the crustacean Daphnia magna. Ecotox. Environ. Saf. 2008, 71, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Fentress, J.A.; Steele, S.L.; Bart, H.L.; Cheek, A.O. Reproductive disruption in wild longear sunfish (Lepomis megalotis) exposed to kraft mill effluent. Environ. Health. Persp. 2006, 114, 40–45. [Google Scholar] [CrossRef]
- Kang, I.J.; Yokota, H.; Oshima, Y.; Tsuruda, Y.; Shimasaki, Y.; Honjo, T. The effects of methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias latipes). Aquat. Toxicol. 2008, 87, 37–46. [Google Scholar] [CrossRef]
- Radl, V.; Pritsch, K.; Munch, J.C.; Schloter, M. Structural and functional diversity of microbial communities from a lake sediment contaminated with trenbolone, an endocrine-disrupting chemical. Environ. Pollut. 2005, 137, 345–353. [Google Scholar] [CrossRef]
- Jenkins, R.; Angus, R.A.; McNatt, H.; Howell, W.M.; Kemppainen, J.A.; Kirk, M.; Wilson, E.M. Identification of androstenedione in a river containing paper mill effluent. Environ. Toxicol. Chem. 2001, 20, 1325–1331. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, R.L.; Wilson, E.M.; Angus, R.A.; Howell, W.M.; Kirk, M. Androstenedione and progesterone in the sediment of a river receiving paper mill effluent. Toxicol. Sci. 2003, 73, 53–59. [Google Scholar]
- Kolok, A.S.; Sellin, M.K. The Environmental Impact of Growth-Promoting Compounds Employed by the United States Beef Cattle Industry: History, Current Knowledge, and Future Directions. In Reviews of Environmental Contamination and Toxicology. Reviews of Environmental Contamination and Toxicology; Whitacre, D.M., Ed.; Springer: New York, NY, USA, 2008; Volume 195. [Google Scholar]
- Kolodziej, E.P.; Sedlak, D.L. Rangeland grazing as a source of steroid hormones to surface waters. Environ. Sci. Technol. 2007, 41, 3514–3520. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, A.; Hendel, J.G.; Conn, K.L.; Bittman, S.; Kwabiah, A.B.; Lazarovitz, G.; Massé, D.; McAllister, T.A.; Topp, E. Survey of hormone activities in municipal biosolids and animal manures. Environ. Toxicol. 2004, 19, 216–225. [Google Scholar] [CrossRef]
- Murray, C.M.; Merchant, M.; Easter, M.; Padilla, S.; Garrigós, D.B.; Sasa Marin, M.; Guyer, C. Detection of a synthetic sex steroid in the American crocodile (Crocodylus acutus): Evidence for a novel environmental androgen. Chemosphere 2017, 180, 125–129. [Google Scholar] [CrossRef]
- Jenkins, M.B.; Endale, D.M.; Schomberg, H.H.; Hartel, P.G.; Cabrera, M.L. 17β-Estradiol and testosterone in drainage and runoff from poultry litter applications to tilled and no-till crop land under irrigation. J. Environ. Manag. 2009, 90, 2659–2664. [Google Scholar] [CrossRef]
- Arnon, S.; Dahan, O.; Elhanany, S.; Cohen, K.; Pankratov, I.; Gross, A.; Ronen, Z.; Baram, S.; Shore, L.S. Transport of testosterone and estrogen from dairy-farm waste lagoons to groundwater. Environ. Sci. Technol. 2008, 42, 5521–5526. [Google Scholar] [CrossRef] [PubMed]
- Lange, I.G.; Daxenberger, A.; Schiffer, B.; Witters, H.; Ibarreta, D.; Meyer, H.H.D. Sex hormones originating from different livestock production systems: Fate and potential disrupting activity in the environment. Anal. Chim. Acta 2002, 473, 27–37. [Google Scholar] [CrossRef]
- Vulliet, E.; Wiest, L.; Baudot, R.; Grenier-Loustalot, M. Multi-residue analysis of steroids at sub-ng/L levels in surface and ground-waters using liquid chromatography coupled to tandem mass spectrometry. J. Chromatogr. 2008, 1210, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Orn, S.; Svenson, A.; Viktor, T.; Holbech, H.; Norrgren, L. Male-biased sex ratios and vitellogenin induction in zebrafish exposed to effluent water from a Swedish pulp mill. Arch. Environ. Contam. Toxicol. 2006, 51, 445–451. [Google Scholar] [CrossRef]
- Orlando, E.F.; Kolok, A.S.; Binzcik, G.A.; Gates, J.L.; Horton, M.K.; Lambright, C.S.; Gray, L.E.; Soto, A.M.; Guillette, L.J. Endocrine-disrupting effects of cattle feedlot effluent on an aquatic sentinel species, the fathead minnow. Environ. Health. Persp. 2004, 112, 353–358. [Google Scholar] [CrossRef]
- Hotchkiss, A.K.; Lambright, C.S.; Ostby, J.S.; Parks-Saldutti, L.; Vandenbergh, J.G.; Gray, L.E. Prenatal testosterone exposure permanently masculinizes anogenital distance, nipple development, and reproductive tract morphology in female Sprague-Dawley rats. Toxicol. Sci. 2007, 96, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, A.K.; Ostby, J.S.; Vandenburgh, J.G.; Gray, L.E. Androgens and environmental antiandrogens affect reproductive development and play behavior in the Sprague-Dawley rat. Environ. Health. Persp. 2002, 110, 435–439. [Google Scholar] [CrossRef]
- Jacobsen, A.-M.; Lorenzen, A.; Chapman, R.; Topp, E. Persistence of testosterone and 17beta-estradiol in soils receiving swine manure or municipal biosolids. J. Environ. Qual. 2005, 34, 861–871. [Google Scholar] [CrossRef]
- Khan, B.; Lee, L.S.; Sassman, S.A. Degradation of Synthetic Androgens 17α- and 17β-Trenbolone and Trendione in Agricultural Soils. Environ. Sci. Technol. 2008, 42, 3570–3574. [Google Scholar] [CrossRef]
- Bradley, P.M.; Barber, L.B.; Chapelle, F.H.; Gray, J.L.; Kolpin, D.W.; McMahon, P.B. Biodegradation of 17β-Estradiol, Estrone and Testosterone in Stream Sediments. Environ. Sci. Technol. 2009, 43, 1902–1910. [Google Scholar] [CrossRef]
- Liu, W.; Chen, Q.; He, N.; Sun, K.; Sun, D.; Wu, X.; Duan, S. Removal and Biodegradation of 17β-Estradiol and Diethylstilbestrol by the Freshwater Microalgae Raphidocelis subcapitata. Int. J. Environ. Res. Pub. Health 2018, 15, 452. [Google Scholar] [CrossRef]
- Pan, C.; Peng, F.; Ying, G. Removal, biotransformation and toxicity variations of climbazole by freshwater algae Scenedesmus obliquus. Environ. Pollut. 2018, 240, 534–540. [Google Scholar] [CrossRef]
- He, N.; Sun, X.; Zhong, Y.; Sun, K.; Liu, W.; Duan, S. Removal and Biodegradation of Nonylphenol by Four Freshwater Microalgae. Int. J. Environ. Res. Pub. Health 2016, 13, 1239. [Google Scholar] [CrossRef]
- Rashkov, G.D.; Dobrikova, A.G.; Pouneva, I.D.; Misra, A.N.; Apostolova, E.L. Sensitivity of Chlorella vulgaris to herbicides. Possibility of using it as a biological receptor in biosensors. Sens. Actuators Chem. 2012, 161, 151–155. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, G.; Ding, J.; Zou, H.; Shi, H.; Huang, C. Evaluation of the Removal of Potassium Cyanide and its Toxicity in Green Algae (Chlorella vulgaris). Bull. Environ. Contam. Toxicol. 2018, 100, 228–233. [Google Scholar] [CrossRef]
- Lai, K.M.; Scrimshaw, M.D.; Lester, J.N. Biotransformation and bioconcentration of steroid estrogens by Chlorella vulgaris. Appl. Environ. Microb. 2002, 68, 859–864. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar]
- Bricaud, A.; Bédhomme, A.L.; Morel, A. Optical properties of diverse phytoplanktonic species: Experimental results and theoretical interpretation. J. Plankton. Res. 1988, 10, 851–873. [Google Scholar] [CrossRef]
- Yan, H.; Ye, C.; Yin, C. Kinetics of phthalate ester biodegradation by Chlorella pyrenoidosa. Environ. Toxicol. Chem. 1995, 14, 931–938. [Google Scholar] [CrossRef]
- Brewer, R. Principles of Ecology; W.B. Saunders Company: Philadelphia, PA, USA, 1979. [Google Scholar]
- Skoglund, R.S.; Stange, K.; Swackhamer, D.L. A Kinetics Model for Predicting the Accumulation of PCBs in Phytoplankton. Environ. Sci. Technol. 1996, 30, 2113–2120. [Google Scholar] [CrossRef]
- Zaranko, D.T.; Griffiths, R.W.; Kaushik, N.K. Biomagnification of polychlorinated biphenyls through a riverine food web. Environ. Toxicol. Chem. 1997, 16, 1463–1471. [Google Scholar] [CrossRef]
- Song, C.; Wei, Y.; Qiu, Y.; Qi, Y.; Li, Y.; Kitamura, Y. Biodegradability and mechanism of florfenicol via Chlorella sp. UTEX1602 and L38: Experimental study. Bioresour. Technol. 2018, 272, 529–534. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Testosterone Concentration (mg/L) | Initial Density of Algal Cells (cell/mL) |
|---|---|---|
| High-concentration | 0.2 | 5 × 106 |
| Low-concentration | 0.02 | 5 × 106 |
| Control I | 0.2 | 〇 1 |
| Control II | 0.02 | 〇 |
| Control III | 〇 | 5 × 106 |
| Time (h) | Density of Cells (106 cell/mL) | ||
|---|---|---|---|
| Control III | Low-Concentration Group | High-Concentration Group | |
| 0 | 5.18 ± 0.04 | 5.21 ± 0.05 | 5.23 ± 0.02 |
| 0.5 | 5.28 ± 0.02 | 5.21 ± 0.05 | 5.30 ± 0.02 |
| 1 | 5.43 ± 0.10 | 5.21 ± 0.05 | 5.43 ± 0.02 |
| 3 | 6.02 ± 0.40 | 6.09 ± 0.00 | 6.18 ± 0.10 |
| 6 | 7.38 ± 0.17 | 7.44 ± 0.08 | 7.46 ± 0.09 |
| 12 | 9.99 ± 0.15 | 10.18 ± 0.05 | 10.53 ± 0.10 |
| 24 | 13.71 ± 0.06 | 13.89 ± 0.04 | 14.29 ± 0.19 |
| 48 | 30.82 ± 0.15 | 30.70 ± 0.56 | 30.40 ± 0.66 |
| 96 | 52.83 ± 0.42 | 52.18 ± 0.41 | 52.49 ± 0.32 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, M.; Deng, B.; Lü, H.; Yao, W.; Su, S.; Wang, D. The Bioaccumulation and Biodegradation of Testosterone by Chlorella vulgaris. Int. J. Environ. Res. Public Health 2019, 16, 1253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16071253
Fu M, Deng B, Lü H, Yao W, Su S, Wang D. The Bioaccumulation and Biodegradation of Testosterone by Chlorella vulgaris. International Journal of Environmental Research and Public Health. 2019; 16(7):1253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16071253
Chicago/Turabian StyleFu, Mei, Bixiang Deng, Hongjian Lü, Weizhi Yao, Shengqi Su, and Dingyong Wang. 2019. "The Bioaccumulation and Biodegradation of Testosterone by Chlorella vulgaris" International Journal of Environmental Research and Public Health 16, no. 7: 1253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph16071253