Priming Effects of Water Immersion on Paired Associative Stimulation-Induced Neural Plasticity in the Primary Motor Cortex

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. TMS
2.3. Electromyographic Recording
2.4. PAS Session
2.5. WI Intervention
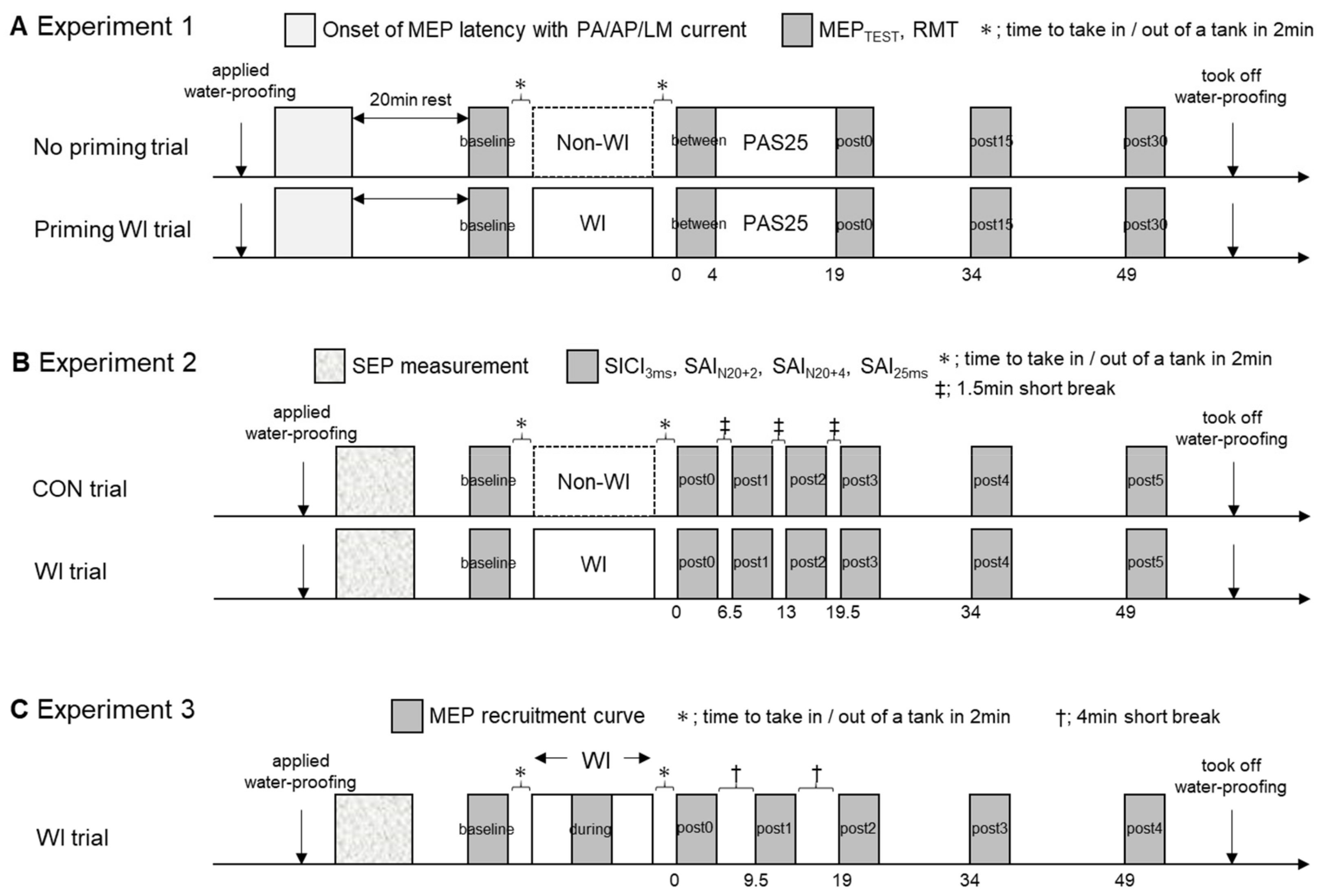
2.6. Experimental Design and Parameters
2.6.1. Experiment 1
2.6.2. Experiment 2
2.6.3. Experiment 3
2.7. Data Analysis and Statistics
2.7.1. Experiment 1
2.7.2. Experiment 2
2.7.3. Experiment 3
3. Results
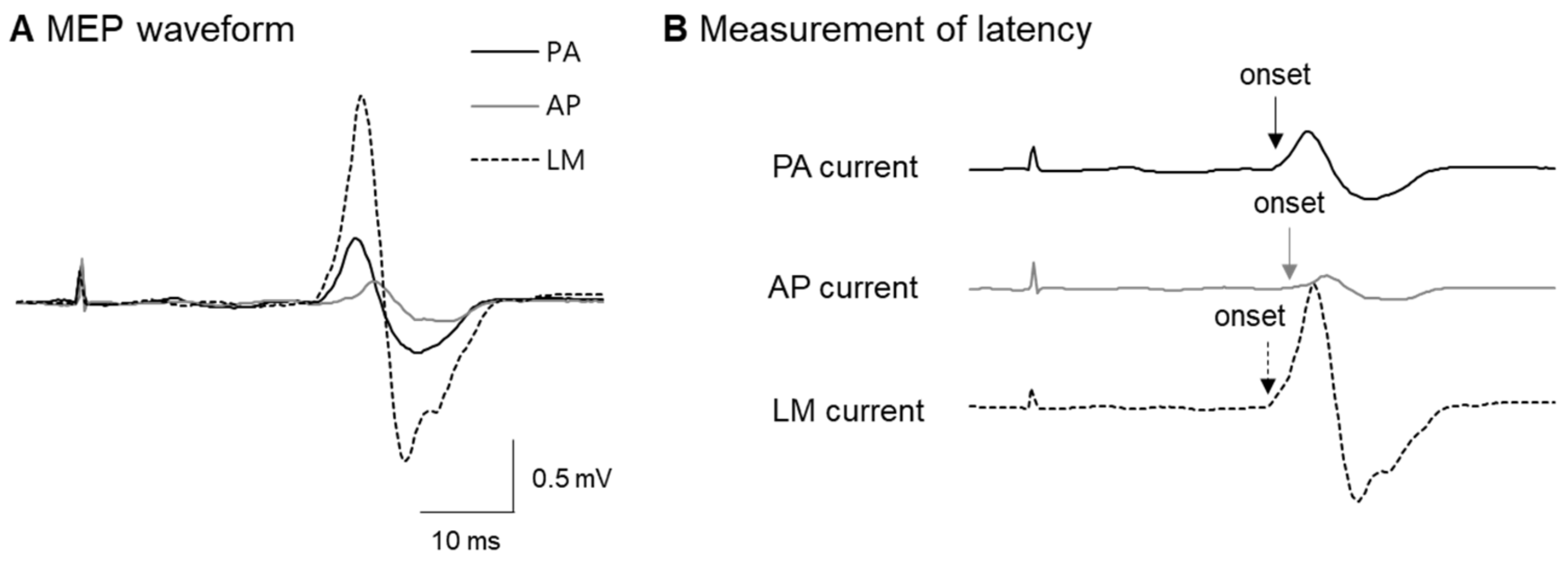
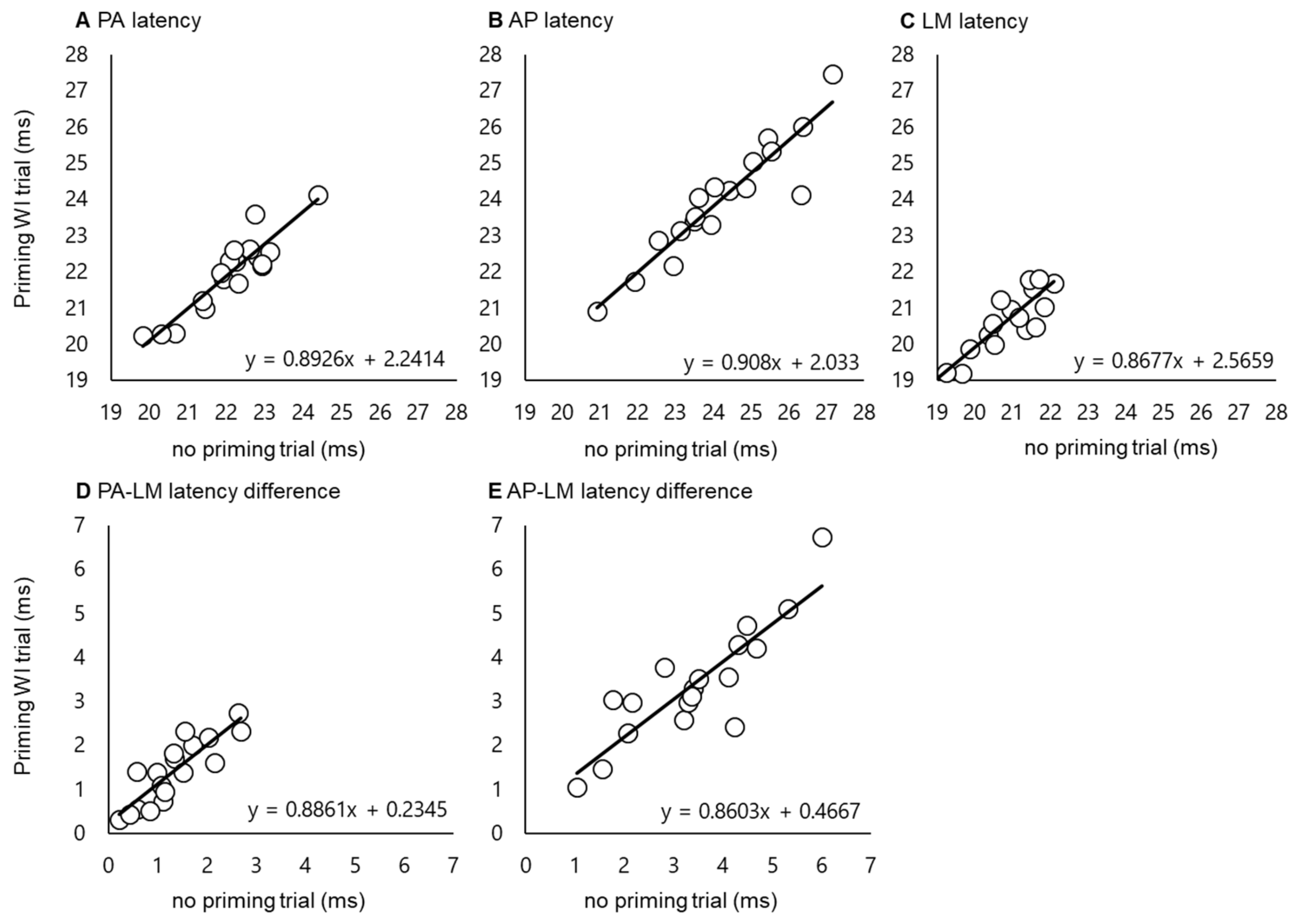
3.1. Latency Difference among Different Coil Orientations
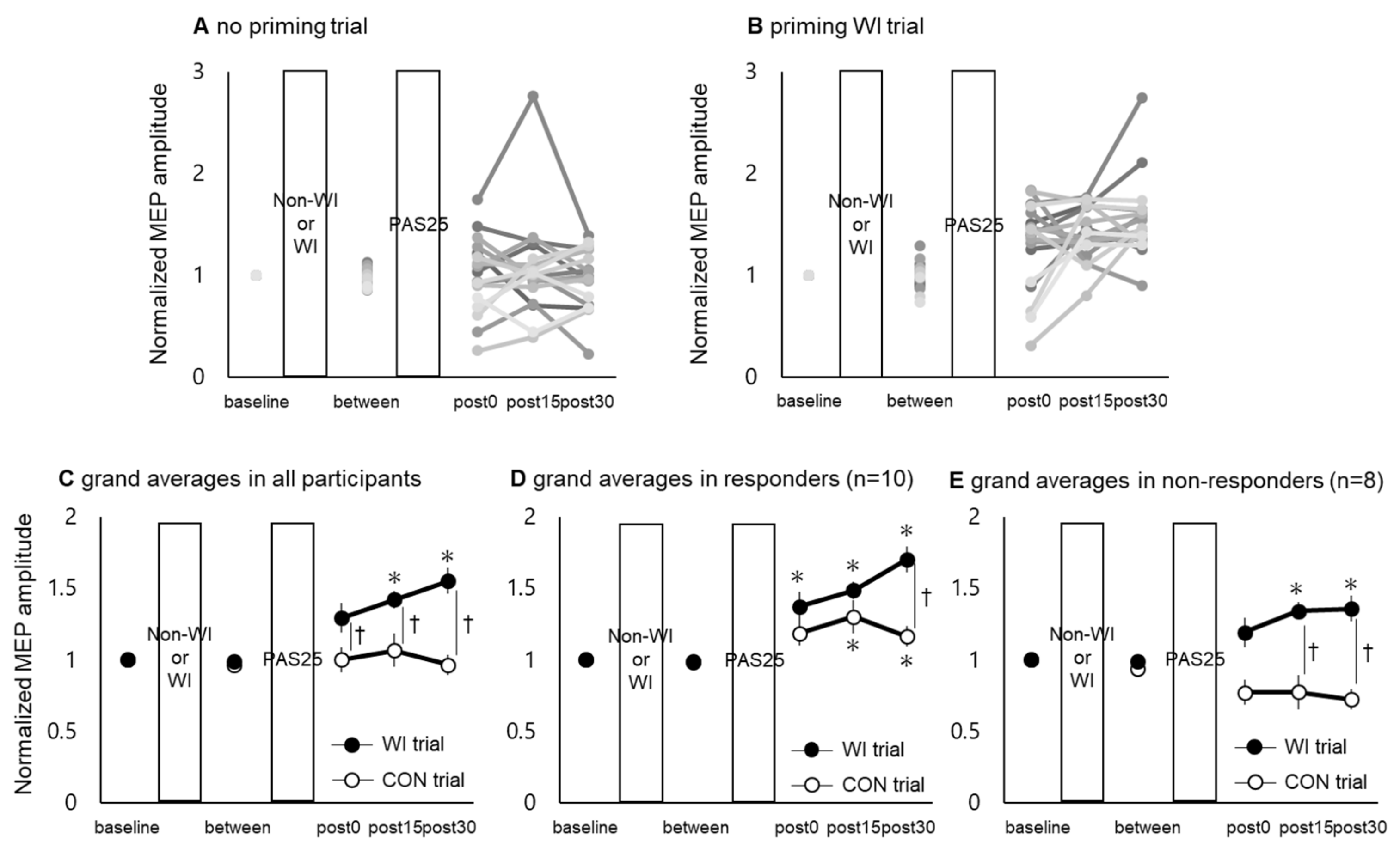
3.2. PAS25-Induced Plasticity in M1
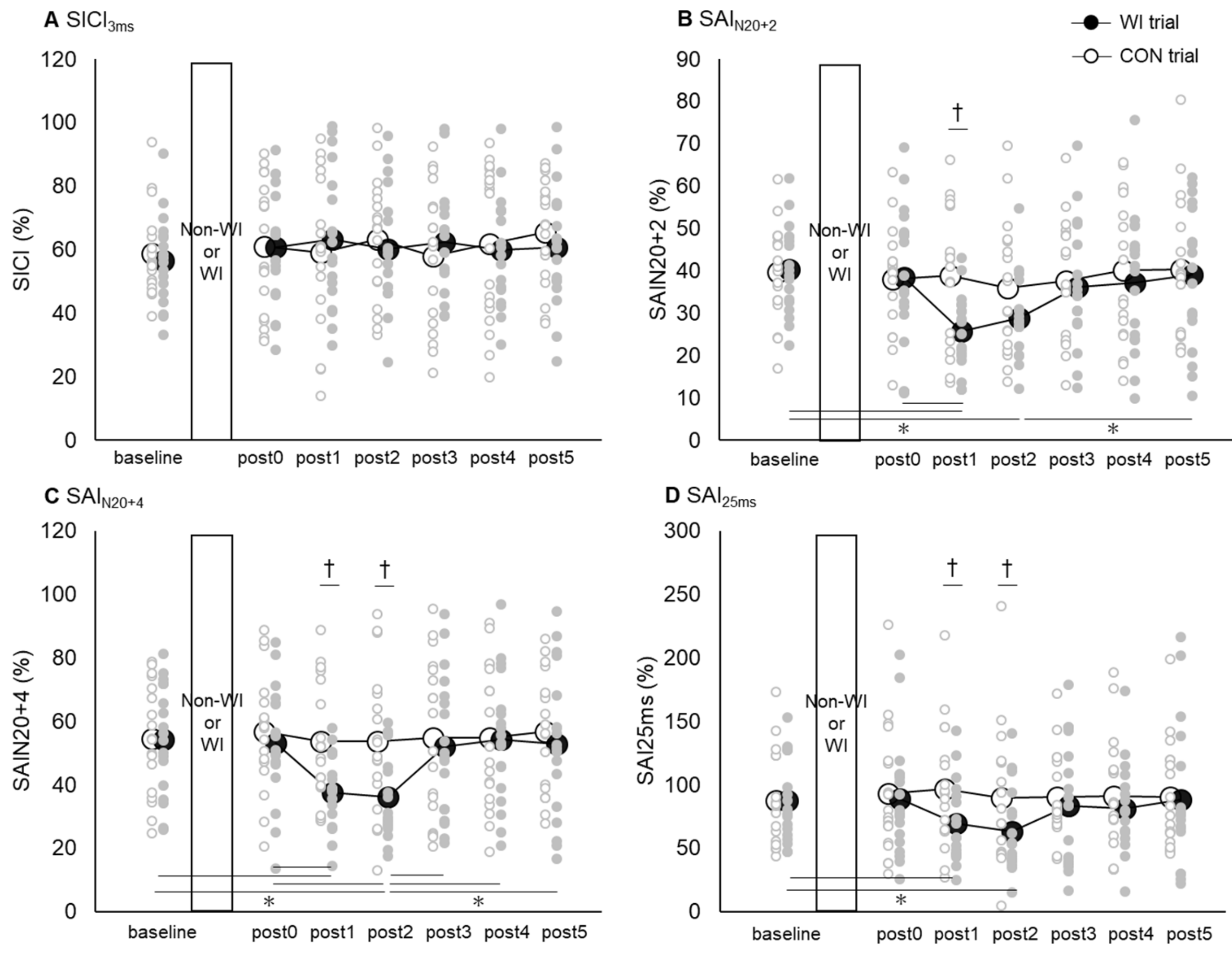
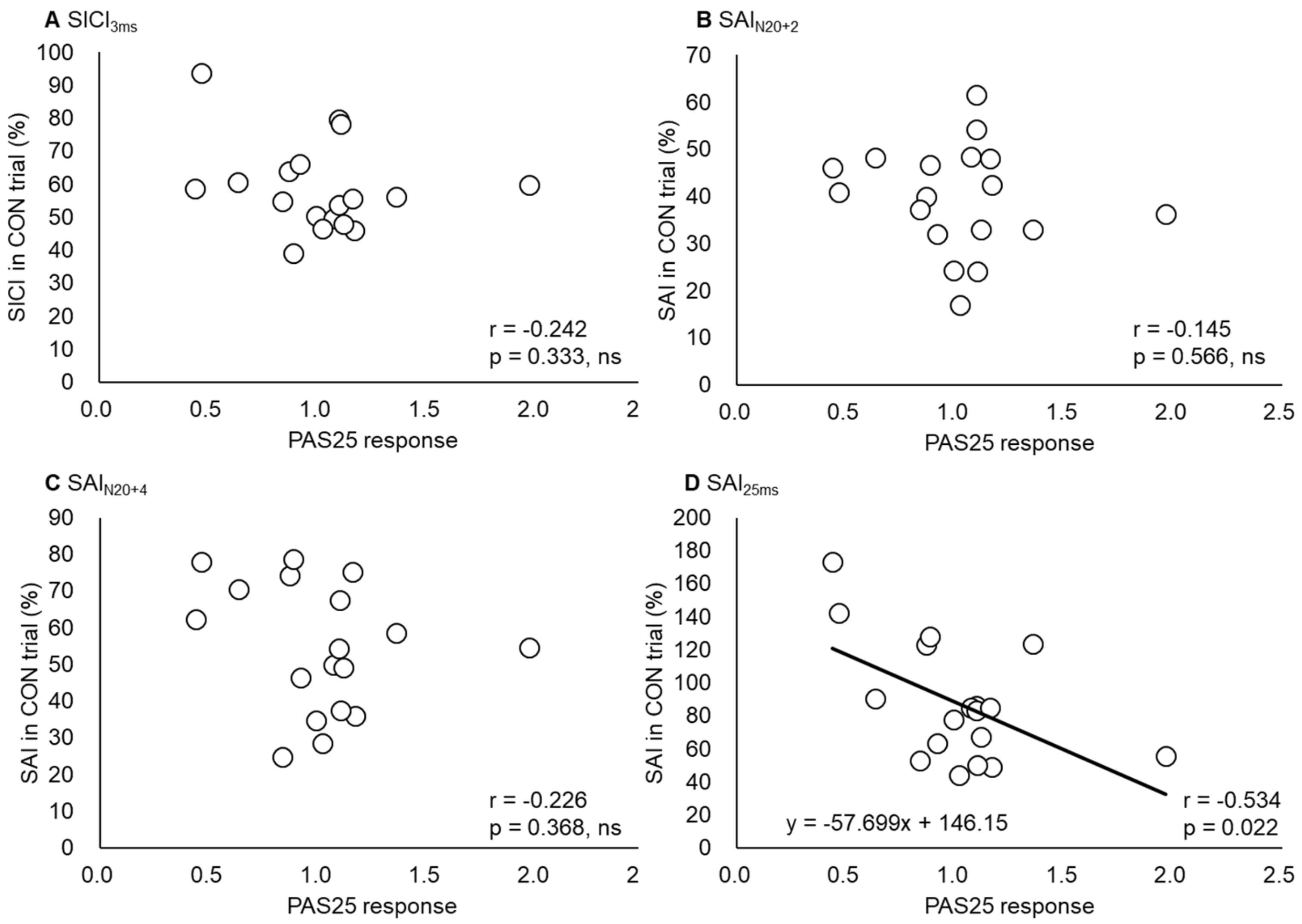
3.3. SAI and SICI
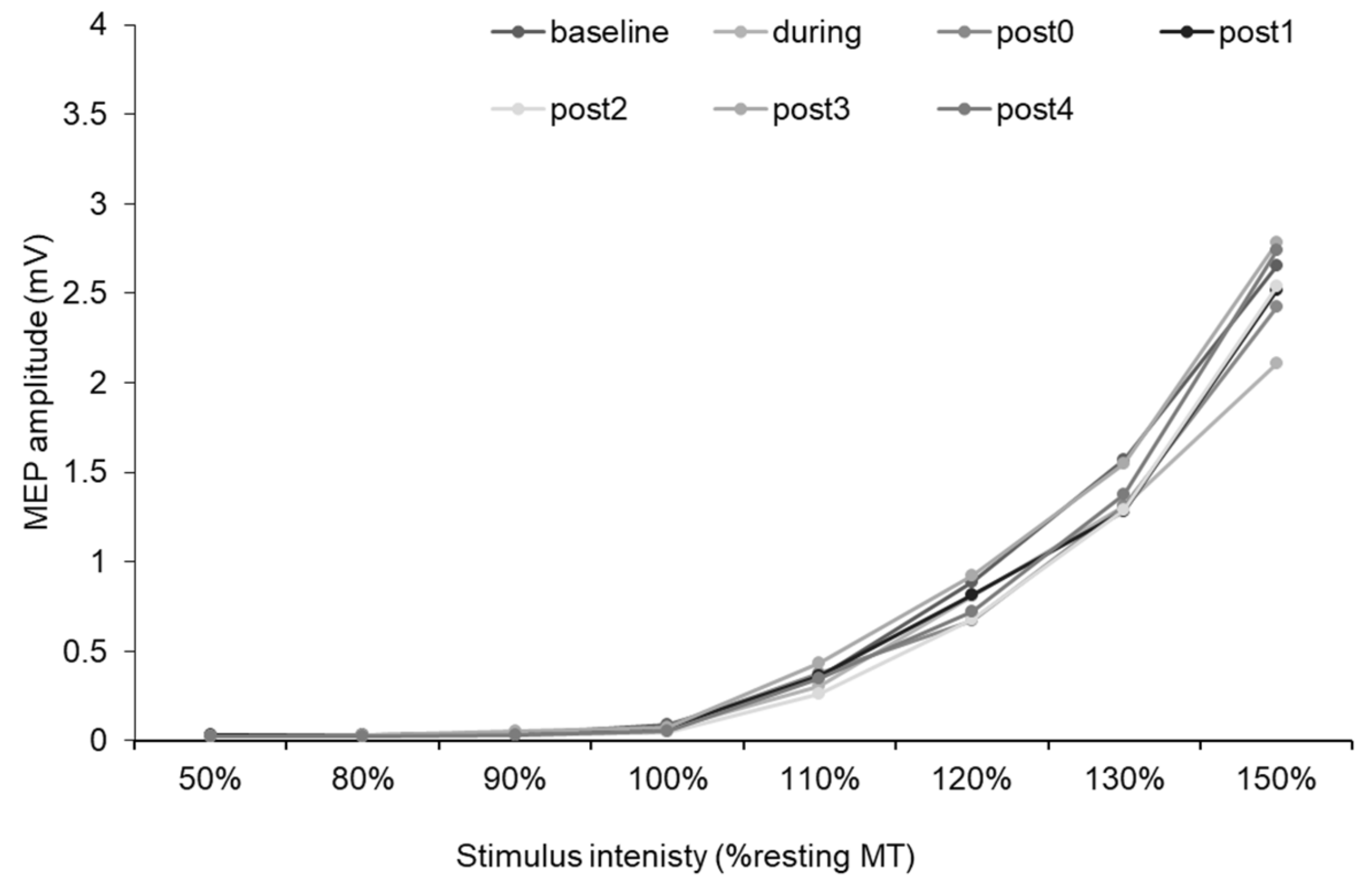
3.4. MEP Recruitment Curve
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Martin, S.J.; Grimwood, P.D.; Morris, R.G. Synaptic plasticity and memory: An evaluation of the hypothesis. Annu. Rev. Neurosci. 2000, 23, 649–711. [Google Scholar] [CrossRef] [Green Version]
- Sjostrom, P.J.; Turrigiano, G.G.; Nelson, S.B. Rate, timing, and cooperativity jointly determine cortical synaptic plasticity. Neuron 2001, 32, 1149–1164. [Google Scholar] [CrossRef] [Green Version]
- Nelson, S.B.; Turrigiano, G.G. Strength through diversity. Neuron 2008, 60, 477–482. [Google Scholar] [CrossRef] [Green Version]
- Abraham, W.C.; Bear, M.F. Metaplasticity: The plasticity of synaptic plasticity. Trends Neurosci. 1996, 19, 126–130. [Google Scholar] [CrossRef]
- Muller-Dahlhaus, F.; Ziemann, U. Metaplasticity in human cortex. Neuroscientist 2015, 21, 185–202. [Google Scholar] [CrossRef] [PubMed]
- Stefan, K.; Kunesch, E.; Cohen, L.G.; Benecke, R.; Classen, J. Induction of plasticity in the human motor cortex by paired associative stimulation. Brain 2000, 123, 572–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fratello, F.; Veniero, D.; Curcio, G.; Ferrara, M.; Marzano, C.; Moroni, F.; Pellicciari, M.C.; Bertini, M.; Rossini, P.M.; De Gennaro, L. Modulation of corticospinal excitability by paired associative stimulation: Reproducibility of effects and intraindividual reliability. Clin. Neurophysiol. 2006, 117, 2667–2674. [Google Scholar] [CrossRef] [PubMed]
- Stefan, K.; Wycislo, M.; Gentner, R.; Schramm, A.; Naumann, M.; Reiners, K.; Classen, J. Temporary occlusion of associative motor cortical plasticity by prior dynamic motor training. Cereb. Cortex 2006, 16, 376–385. [Google Scholar] [CrossRef] [Green Version]
- Muller-Dahlhaus, J.F.; Orekhov, Y.; Liu, Y.; Ziemann, U. Interindividual variability and age-dependency of motor cortical plasticity induced by paired associative stimulation. Exp. Brain Res. 2008, 187, 467–475. [Google Scholar] [CrossRef]
- Hamada, M.; Galea, J.M.; Di Lazzaro, V.; Mazzone, P.; Ziemann, U.; Rothwell, J.C. Two distinct interneuron circuits in human motor cortex are linked to different subsets of physiological and behavioral plasticity. J. Neurosci. 2014, 34, 12837–12849. [Google Scholar] [CrossRef]
- Lopez-Alonso, V.; Cheeran, B.; Rio-Rodriguez, D.; Fernandez-Del-Olmo, M. Inter-individual variability in response to non-invasive brain stimulation paradigms. Brain Stimul. 2014, 7, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Ridding, M.C.; Ziemann, U. Determinants of the induction of cortical plasticity by non-invasive brain stimulation in healthy subjects. J. Physiol. 2010, 588, 2291–2304. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Murase, N.; Hasan, A.; Balaratnam, M.; Rothwell, J.C. The role of interneuron networks in driving human motor cortical plasticity. Cereb. Cortex 2013, 23, 1593–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murase, N.; Cengiz, B.; Rothwell, J.C. Inter-individual variation in the after-effect of paired associative stimulation can be predicted from short-interval intracortical inhibition with the threshold tracking method. Brain Stimul. 2015, 8, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Kirimoto, H.; Sugawara, K.; Watanabe, M.; Shimizu, S.; Ishizaka, I.; Yamada, S.; Matsunaga, A.; Fukuda, M.; Onishi, H. Induction of cortical plasticity for reciprocal muscles by paired associative stimulation. Brain Behav. 2014, 4, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Udupa, K.; Ni, Z.; Gunraj, C.; Chen, R. Interactions between short latency afferent inhibition and long interval intracortical inhibition. Exp. Brain Res. 2009, 199, 177–183. [Google Scholar] [CrossRef]
- Udupa, K.; Ni, Z.; Gunraj, C.; Chen, R. Effects of short-latency afferent inhibition on short-interval intracortical inhibition. J. Neurophysiol. 2014, 111, 1350–1361. [Google Scholar] [CrossRef] [Green Version]
- Iyer, M.B.; Schleper, N.; Wassermann, E.M. Priming stimulation enhances the depressant effect of low-frequency repetitive transcranial magnetic stimulation. J. Neurosci. 2003, 23, 10867–10872. [Google Scholar] [CrossRef]
- Muller, J.F.; Orekhov, Y.; Liu, Y.; Ziemann, U. Homeostatic plasticity in human motor cortex demonstrated by two consecutive sessions of paired associative stimulation. Eur. J. Neurosci. 2007, 25, 3461–3468. [Google Scholar] [CrossRef]
- Sato, D.; Onishi, H.; Yamashiro, K.; Iwabe, T.; Shimoyama, Y.; Maruyama, A. Water immersion to the femur level affects cerebral cortical activity in humans: Functional near-infrared spectroscopy study. Brain Topogr. 2012, 25, 220–227. [Google Scholar] [CrossRef]
- Sato, D.; Yamashiro, K.; Onishi, H.; Shimoyama, Y.; Yoshida, T.; Maruyama, A. The effect of water immersion on short-latency somatosensory evoked potentials in human. BMC Neurosci. 2012, 13, 13. [Google Scholar] [CrossRef] [Green Version]
- Di Lazzaro, V.; Oliviero, A.; Profice, P.; Pennisi, M.A.; Di Giovanni, S.; Zito, G.; Tonali, P.; Rothwell, J.C. Muscarinic receptor blockade has differential effects on the excitability of intracortical circuits in the human motor cortex. Exp. Brain Res. 2000, 135, 455–461. [Google Scholar] [CrossRef]
- Brown, R.E.; Basheer, R.; McKenna, J.T.; Strecker, R.E.; McCarley, R.W. Control of sleep and wakefulness. Physiol. Rev. 2012, 92, 1087–1187. [Google Scholar] [CrossRef] [Green Version]
- Kuo, M.F.; Grosch, J.; Fregni, F.; Paulus, W.; Nitsche, M.A. Focusing effect of acetylcholine on neuroplasticity in the human motor cortex. J. Neurosci. 2007, 27, 14442–14447. [Google Scholar] [CrossRef]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [Green Version]
- Buharin, V.E.; Shinohara, M. Corticospinal excitability for flexor carpi radialis decreases with baroreceptor unloading during intentional co-contraction with opposing forearm muscles. Exp. Brain Res. 2019, 237, 1947–1958. [Google Scholar] [CrossRef]
- Sato, D.; Yamashiro, K.; Onishi, H.; Yasuhiro, B.; Shimoyama, Y.; Maruyama, A. Whole-hand water flow stimulation increases motor cortical excitability: A study of transcranial magnetic stimulation and movement-related cortical potentials. J. Neurophysiol. 2015, 113, 822–833. [Google Scholar] [CrossRef] [Green Version]
- Stefan, K.; Wycislo, M.; Classen, J. Modulation of associative human motor cortical plasticity by attention. J. Neurophysiol. 2004, 92, 66–72. [Google Scholar] [CrossRef]
- Sale, M.V.; Ridding, M.C.; Nordstrom, M.A. Cortisol inhibits neuroplasticity induction in human motor cortex. J. Neurosci. 2008, 28, 8285–8293. [Google Scholar] [CrossRef] [Green Version]
- Arai, N.; Okabe, S.; Furubayashi, T.; Terao, Y.; Yuasa, K.; Ugawa, Y. Comparison between short train, monophasic and biphasic repetitive transcranial magnetic stimulation (rTMS) of the human motor cortex. Clin. Neurophysiol. 2005, 116, 605–613. [Google Scholar] [CrossRef]
- Sakai, K.; Ugawa, Y.; Terao, Y.; Hanajima, R.; Furubayashi, T.; Kanazawa, I. Preferential activation of different I waves by transcranial magnetic stimulation with a figure-of-eight-shaped coil. Exp. Brain Res. 1997, 113, 24–32. [Google Scholar] [CrossRef]
- Day, B.L.; Rothwell, J.C.; Thompson, P.D.; Dick, J.P.; Cowan, J.M.; Berardelli, A.; Marsden, C.D. Motor cortex stimulation in intact man. 2. Multiple descending volleys. Brain 1987, 110, 1191–1209. [Google Scholar] [CrossRef]
- Rothwell, J.C.; Thompson, P.D.; Day, B.L.; Dick, J.P.; Kachi, T.; Cowan, J.M.; Marsden, C.D. Motor cortex stimulation in intact man. 1. General characteristics of EMG responses in different muscles. Brain 1987, 110, 1173–1190. [Google Scholar] [CrossRef]
- Kujirai, T.; Caramia, M.D.; Rothwell, J.C.; Day, B.L.; Thompson, P.D.; Ferbert, A.; Wroe, S.; Asselman, P.; Marsden, C.D. Corticocortical inhibition in human motor cortex. J. Physiol. 1993, 471, 501–519. [Google Scholar] [CrossRef]
- Ziemann, U.; Rothwell, J.C.; Ridding, M.C. Interaction between intracortical inhibition and facilitation in human motor cortex. J. Physiol. 1996, 496, 873–881. [Google Scholar] [CrossRef]
- Tokimura, H.; Di Lazzaro, V.; Tokimura, Y.; Oliviero, A.; Profice, P.; Insola, A.; Mazzone, P.; Tonali, P.; Rothwell, J.C. Short latency inhibition of human hand motor cortex by somatosensory input from the hand. J. Physiol. 2000, 523, 503–513. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Oliviero, A.; Saturno, E.; Dileone, M.; Pilato, F.; Nardone, R.; Ranieri, F.; Musumeci, G.; Fiorilla, T.; Tonali, P. Effects of lorazepam on short latency afferent inhibition and short latency intracortical inhibition in humans. J. Physiol. 2005, 564, 661–668. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Pilato, F.; Dileone, M.; Profice, P.; Ranieri, F.; Ricci, V.; Bria, P.; Tonali, P.A.; Ziemann, U. Segregating two inhibitory circuits in human motor cortex at the level of GABAA receptor subtypes: A TMS study. Clin. Neurophysiol. 2007, 118, 2207–2214. [Google Scholar] [CrossRef]
- Sato, D.; Yamashiro, K.; Yoshida, T.; Onishi, H.; Shimoyama, Y.; Maruyama, A. Effects of water immersion on short- and long-latency afferent inhibition, short-interval intracortical inhibition, and intracortical facilitation. Clin. Neurophysiol. 2013, 124, 1846–1852. [Google Scholar] [CrossRef]
- Sato, D.; Kaneda, K.; Wakabayashi, H.; Nomura, T. Comparison of 2-year effects of once and twice weekly water exercise on activities of daily living ability of community dwelling frail elderly. Arch. Gerontol. Geriatr. 2009, 49, 123–128. [Google Scholar] [CrossRef]
- Sato, D.; Kaneda, K.; Wakabayashi, H.; Shimoyama, Y.; Baba, Y.; Nomura, T. Comparison of once and twice weekly water exercise on various bodily functions in community-dwelling frail elderly requiring nursing care. Arch. Gerontol. Geriatr. 2011, 52, 331–335. [Google Scholar] [CrossRef]
- Sato, D.; Seko, C.; Hashitomi, T.; Sengoku, Y.; Nomura, T. Differential effects of water-based exercise on the cognitive function in independent elderly adults. Aging Clin. Exp. Res. 2015, 27, 149–159. [Google Scholar] [CrossRef]
- Hamada, M.; Hanajima, R.; Terao, Y.; Okabe, S.; Nakatani-Enomoto, S.; Furubayashi, T.; Matsumoto, H.; Shirota, Y.; Ohminami, S.; Ugawa, Y. Primary motor cortical metaplasticity induced by priming over the supplementary motor area. J. Physiol. 2009, 587, 4845–4862. [Google Scholar] [CrossRef]
- Popa, T.; Velayudhan, B.; Hubsch, C.; Pradeep, S.; Roze, E.; Vidailhet, M.; Meunier, S.; Kishore, A. Cerebellar processing of sensory inputs primes motor cortex plasticity. Cereb. Cortex 2013, 23, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Sato, D.; Yamashiro, K.; Onishi, H.; Baba, Y.; Nakazawa, S.; Shimoyama, Y.; Maruyama, A. Whole-body water flow stimulation to the lower limbs modulates excitability of primary motor cortical regions innervating the hands: A transcranial magnetic stimulation study. PLoS ONE 2014, 9, 102472. [Google Scholar] [CrossRef]
- Kamke, M.R.; Hall, M.G.; Lye, H.F.; Sale, M.V.; Fenlon, L.R.; Carroll, T.J.; Riek, S.; Mattingley, J.B. Visual attentional load influences plasticity in the human motor cortex. J. Neurosci. 2012, 32, 7001–7008. [Google Scholar] [CrossRef]
- Kamke, M.R.; Ryan, A.E.; Sale, M.V.; Campbell, M.E.; Riek, S.; Carroll, T.J.; Mattingley, J.B. Visual spatial attention has opposite effects on bidirectional plasticity in the human motor cortex. J. Neurosci. 2014, 34, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Ziemann, U.; Ilic, T.V.; Pauli, C.; Meintzschel, F.; Ruge, D. Learning modifies subsequent induction of long-term potentiation-like and long-term depression-like plasticity in human motor cortex. J. Neurosci. 2004, 24, 1666–1672. [Google Scholar] [CrossRef]
- Kleim, J.A.; Chan, S.; Pringle, E.; Schallert, K.; Procaccio, V.; Jimenez, R.; Cramer, S.C. BDNF val66met polymorphism is associated with modified experience-dependent plasticity in human motor cortex. Nat. Neurosci. 2006, 9, 735–737. [Google Scholar] [CrossRef] [Green Version]
- Cheeran, B.; Talelli, P.; Mori, F.; Koch, G.; Suppa, A.; Edwards, M.; Houlden, H.; Bhatia, K.; Greenwood, R.; Rothwell, J.C. A common polymorphism in the brain-derived neurotrophic factor gene (BDNF) modulates human cortical plasticity and the response to rTMS. J. Physiol. 2008, 586, 5717–5725. [Google Scholar] [CrossRef]
- Wassermann, E.M. Variation in the response to transcranial magnetic brain stimulation in the general population. Clin. Neurophysiol. 2002, 113, 1165–1171. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Dileone, M.; Pilato, F.; Profice, P.; Oliviero, A.; Mazzone, P.; Insola, A.; Capone, F.; Ranieri, F.; Tonali, P.A. Associative motor cortex plasticity: Direct evidence in humans. Cereb. Cortex 2009, 19, 2326–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller-Dahlhaus, F.; Ziemann, U.; Classen, J. Plasticity resembling spike-timing dependent synaptic plasticity: The evidence in human cortex. Front. Synaptic Neurosci. 2010, 2, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallett, M. Transcranial magnetic stimulation: A primer. Neuron 2007, 55, 187–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta burst stimulation of the human motor cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lazzaro, V.; Profice, P.; Ranieri, F.; Capone, F.; Dileone, M.; Oliviero, A.; Pilato, F. I-wave origin and modulation. Brain Stimul. 2012, 5, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Strigaro, G.; Hamada, M.; Murase, N.; Cantello, R.; Rothwell, J.C. Interaction between different interneuron networks involved in human associative plasticity. Brain Stimul. 2014, 7, 658–664. [Google Scholar] [CrossRef]
- Todd, G.; Flavel, S.C.; Ridding, M.C. Priming theta-burst repetitive transcranial magnetic stimulation with low- and high-frequency stimulation. Exp. Brain Res. 2009, 195, 307–315. [Google Scholar] [CrossRef]
- Hirotsu, I.; Hori, N.; Katsuda, N.; Ishihara, T. Effect of anticholinergic drug on long-term potentiation in rat hippocampal slices. Brain Res. 1989, 482, 194–197. [Google Scholar] [CrossRef]
- Blitzer, R.D.; Gil, O.; Landau, E.M. Cholinergic stimulation enhances long-term potentiation in the CA1 region of rat hippocampus. Neurosci. Lett. 1990, 119, 207–210. [Google Scholar] [CrossRef]
- Huerta, P.T.; Lisman, J.E. Heightened synaptic plasticity of hippocampal CA1 neurons during a cholinergically induced rhythmic state. Nature 1993, 364, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Brocher, S.; Artola, A.; Singer, W. Agonists of cholinergic and noradrenergic receptors facilitate synergistically the induction of long-term potentiation in slices of rat visual cortex. Brain Res. 1992, 573, 27–36. [Google Scholar] [CrossRef]
- Hasselmo, M.E.; Barkai, E. Cholinergic modulation of activity-dependent synaptic plasticity in the piriform cortex and associative memory function in a network biophysical simulation. J. Neurosci. 1995, 15, 6592–6604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meintzschel, F.; Ziemann, U. Modification of practice-dependent plasticity in human motor cortex by neuromodulators. Cereb. Cortex 2006, 16, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Sawaki, L.; Boroojerdi, B.; Kaelin-Lang, A.; Burstein, A.H.; Butefisch, C.M.; Kopylev, L.; Davis, B.; Cohen, L.G. Cholinergic influences on use-dependent plasticity. J. Neurophysiol. 2002, 87, 166–171. [Google Scholar] [CrossRef]
- Blokland, A.; Honig, W.; Raaijmakers, W.G. Effects of intra-hippocampal scopolamine injections in a repeated spatial acquisition task in the rat. Psychopharmacology 1992, 109, 373–376. [Google Scholar] [CrossRef]
- Winters, B.D.; Bussey, T.J. Removal of cholinergic input to perirhinal cortex disrupts object recognition but not spatial working memory in the rat. Eur. J. Neurosci. 2005, 21, 2263–2270. [Google Scholar] [CrossRef]
- Patil, M.M.; Linster, C.; Lubenov, E.; Hasselmo, M.E. Cholinergic agonist carbachol enables associative long-term potentiation in piriform cortex slices. J. Neurophysiol. 1998, 80, 2467–2474. [Google Scholar] [CrossRef]
- Rasmusson, D.D.; Dykes, R.W. Long-term enhancement of evoked potentials in cat somatosensory cortex produced by co-activation of the basal forebrain and cutaneous receptors. Exp. Brain Res. 1988, 70, 276–286. [Google Scholar] [CrossRef]
- Tremblay, N.; Warren, R.A.; Dykes, R.W. Electrophysiological studies of acetylcholine and the role of the basal forebrain in the somatosensory cortex of the cat. II. Cortical neurons excited by somatic stimuli. J. Neurophysiol. 1990, 64, 1212–1222. [Google Scholar] [CrossRef] [Green Version]
- Hordacre, B.; Goldsworthy, M.R.; Vallence, A.M.; Darvishi, S.; Moezzi, B.; Hamada, M.; Rothwell, J.C.; Ridding, M.C. Variability in neural excitability and plasticity induction in the human cortex: A brain stimulation study. Brain Stimul. 2017, 10, 588–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment 1 | ||
|---|---|---|
| Participants’ Characteristics | ||
| Number | 18 (female 3, male 15) | |
| Age (years old) | 21.37 ± 0.18 | |
| TMS and ES Parameter | ||
| No Priming | Priming WI | |
| RMT (%) | 42.28 ± 1.93 | 40.72 ± 1.68 |
| AMTpa (%) | 34.83 ± 1.55 | 32.61 ± 1.29 |
| AMTap (%) | 49.89 ± 2.24 | 47.50 ± 2.00 |
| AMTlm (%) | 37.56 ± 1.67 | 36.50 ± 1.26 |
| TS1mV_base | 52.72 ± 2.37 | 52.83 ± 2.23 |
| ST (mA) | 3.63 ± 0.28 | 3.73 ± 0.34 |
| MEP Latency | ||
| PA (ms) | 22.10 ± 0.26 | 21.96 ± 0.25 |
| AP (ms) | 24.17 ± 0.39 | 23.98 ± 0.38 |
| LM (ms) | 20.77 ± 0.26 | 20.59 ± 0.25 |
| Latency Differences | ||
| PA–LM (ms) | 1.33 ± 0.17 | 1.38 ± 0.19 |
| AP–LM (ms) | 3.40 ± 0.32 | 3.39 ± 0.31 |
| AP–PA (ms) | 2.08 ± 0.25 | 2.02 ± 0.28 |
| Experiment 2 | ||
| Participants’ Characteristics | ||
| Number | 18 (female 3, male 15) | |
| Age (years old) | 21.37 ± 0.18 | |
| N20 (msec) | 18.62 ± 0.16 | |
| TMS and ES Parameter | ||
| CON Trial | WI Trial | |
| RMT (%) | 45.29 ± 1.65 | 44.00 ± 1.02 |
| AMTpa (%) | 35.12 ± 1.26 | 33.61 ± 0.83 |
| TS1mV_base | 58.88 ± 1.99 | 57.44 ± 1.92 |
| ST (mA) | 3.35 ± 0.21 | 3.67 ± 0.20 |
| CSSICI (%) | 31.46 ± 1.22 | 30.48 ± 0.71 |
| CSSAI (mA) | 10.04 ± 0.64 | 10.97 ± 0.55 |
| Experiment 3 | ||
| Participants’ Characteristics | ||
| Number | 9 (female 1, male 8) | |
| Age (y.o.) | 21.89 ± 0.31 | |
| RMT (%) | 42.71 ± 1.77 | |
| Experiment 1 | ||
|---|---|---|
| r | p | |
| RMT | −0.472 | 0.048 * |
| AMTpa | −0.582 | 0.011 * |
| AMTap | −0.351 | 0.154 |
| AMTlm | −0.348 | 0.157 |
| TS1mV_base | −0.437 | 0.047 * |
| ST | 0.288 | 0.246 |
| PA latency | 0.169 | 0.502 |
| AP latency | 0.040 | 0.876 |
| LM latency | 0.095 | 0.706 |
| PA–LM | 0.127 | 0.615 |
| AP–LM | −0.019 | 0.940 |
| Experiment 2 | ||
| r | p | |
| SICI3ms | −0.24 | 0.333 |
| SAIN20+2 | −0.15 | 0.566 |
| SAIN20+4 | −0.23 | 0.368 |
| SAI25ms | −0.53 | 0.022 * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, D.; Yamashiro, K.; Yamazaki, Y.; Ikarashi, K.; Onishi, H.; Baba, Y.; Maruyama, A. Priming Effects of Water Immersion on Paired Associative Stimulation-Induced Neural Plasticity in the Primary Motor Cortex. Int. J. Environ. Res. Public Health 2020, 17, 215. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17010215
Sato D, Yamashiro K, Yamazaki Y, Ikarashi K, Onishi H, Baba Y, Maruyama A. Priming Effects of Water Immersion on Paired Associative Stimulation-Induced Neural Plasticity in the Primary Motor Cortex. International Journal of Environmental Research and Public Health. 2020; 17(1):215. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17010215
Chicago/Turabian StyleSato, Daisuke, Koya Yamashiro, Yudai Yamazaki, Koyuki Ikarashi, Hideaki Onishi, Yasuhiro Baba, and Atsuo Maruyama. 2020. "Priming Effects of Water Immersion on Paired Associative Stimulation-Induced Neural Plasticity in the Primary Motor Cortex" International Journal of Environmental Research and Public Health 17, no. 1: 215. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17010215