1. Introduction
At present, the available arable land is rapidly declining worldwide due, in part, to increased salinity arising from both natural and anthropogenic sources, such as agriculture and oil and gas exploitation activities [
1,
2,
3,
4]. Current methods of mechanical and chemical remediation of disturbed soils are prohibitively expensive, laborious, and disruptive to the local ecology. Bioremediation is a less invasive alternative to chemical and mechanical remediation that typically employs the natural processes of microorganisms and fungi to degrade waste [
5,
6]. Plants can also be used in remediation activities as their macroscopic structures can host microbial communities and may aid in the establishment of successional organisms [
7,
8,
9]. Phytoremediation is the process of improving soil quality using plants via contaminant degradation, stabilization, accumulation, filtration (such as through a constructed wetland), or volatilization. A range of plant species have been identified for potential use in phytoremediation [
10,
11,
12]. However, most native species used for remediation of disturbed sites are susceptible to salt stress, causing declines in germination, establishment, yield, and survival. Salts, in conjunction with pollutants originating from industrial byproducts, cause osmotic stress, cellular toxicity, and nutrient imbalance [
13]. Most halophytic species that can withstand these stresses are not appropriate for phytoremediation efforts: they are usually not native to disturbed sites and are often slower growing with lower total biomass accrual. Therefore, it is imperative to identify native plant species with useful traits for phytoremediation.
Members of the
Salicaceae have shown marked abiotic stress tolerance and have attracted interest for the phytoremediation of saline and industrially polluted land due to a variety of advantageous traits [
14,
15,
16,
17]. For example, both poplars and willows exhibit fast and expansive root growth, can propagate via coppicing, have a wealth of genetic resources, and are widespread in the Northern Hemisphere [
18]. There exists an expanding body of knowledge regarding salinity tolerance in some poplar and willow species, however, a gap remains for
Populus balsamifera L. (balsam poplar) and
Salix eriocephala Michx. (heart leaf willow) which occupy a significant portion of the temperate-boreal forest of North America. By screening for tolerance in native poplar and willow species, there is an opportunity to identify and select genotypes that may be well suited for remediation of industrially disturbed sites. These observations could aid in the selection and breeding of native genotypes for use in amelioration of deteriorated soils, as well as restoration of local biodiversity. Moreover, this information would help elucidate the internal mechanisms that poplar and willow species use to cope with abiotic stress, such as salinity.
Previous studies on balsam poplar examined clinal variation in physiology as well as observed genotypic variation in response to moisture stress [
19]. Additionally, balsam poplar has been used as a pioneer species in the reclamation and restoration of logging roads and well pads, providing a foundation for successional species that contribute to remediation [
20,
21]. These initial observations encourage the investigation of other abiotic stress responses, including potential tolerance to salinity for use in saline well pad and spill site reclamation. The objectives of this study were to observe the physiological response of
Populus balsamifera and
Salix eriocephala to salinity and hydraulic fracturing (fracking) wastewater by measuring growth and survival, gas exchange, non-structural carbohydrate production, and nutrient balance. We hypothesized that these genera would display tolerance primarily through the exclusion of salts from aboveground tissues. These results could directly benefit the remediation efforts of the oil and gas industries, contribute to the existing body of knowledge concerning salt tolerance in glycophytic species, and create opportunities for further genetic research on abiotic stress tolerance.
4. Discussion
The salt tolerance of
Populus balsamifera is not well reported, nor is the effect of fracking wastewater on terrestrial plant species [
23]. Furthermore, an examination of the salinity tolerance of native Canadian
Salix eriocephala genotypes contributes to a limited body of knowledge while providing solutions for industrial reclamation [
24]. Salinity treatments ranging from 20 to 80 mM NaCl were chosen to parallel salinity studies performed on closely related species [
25,
26,
27]. We found that the salt treated willows demonstrated a far greater tolerance to saline conditions than the balsam poplar, and that balsam poplar is likely unsuitable for reclamation of Na
+ dominated soils.
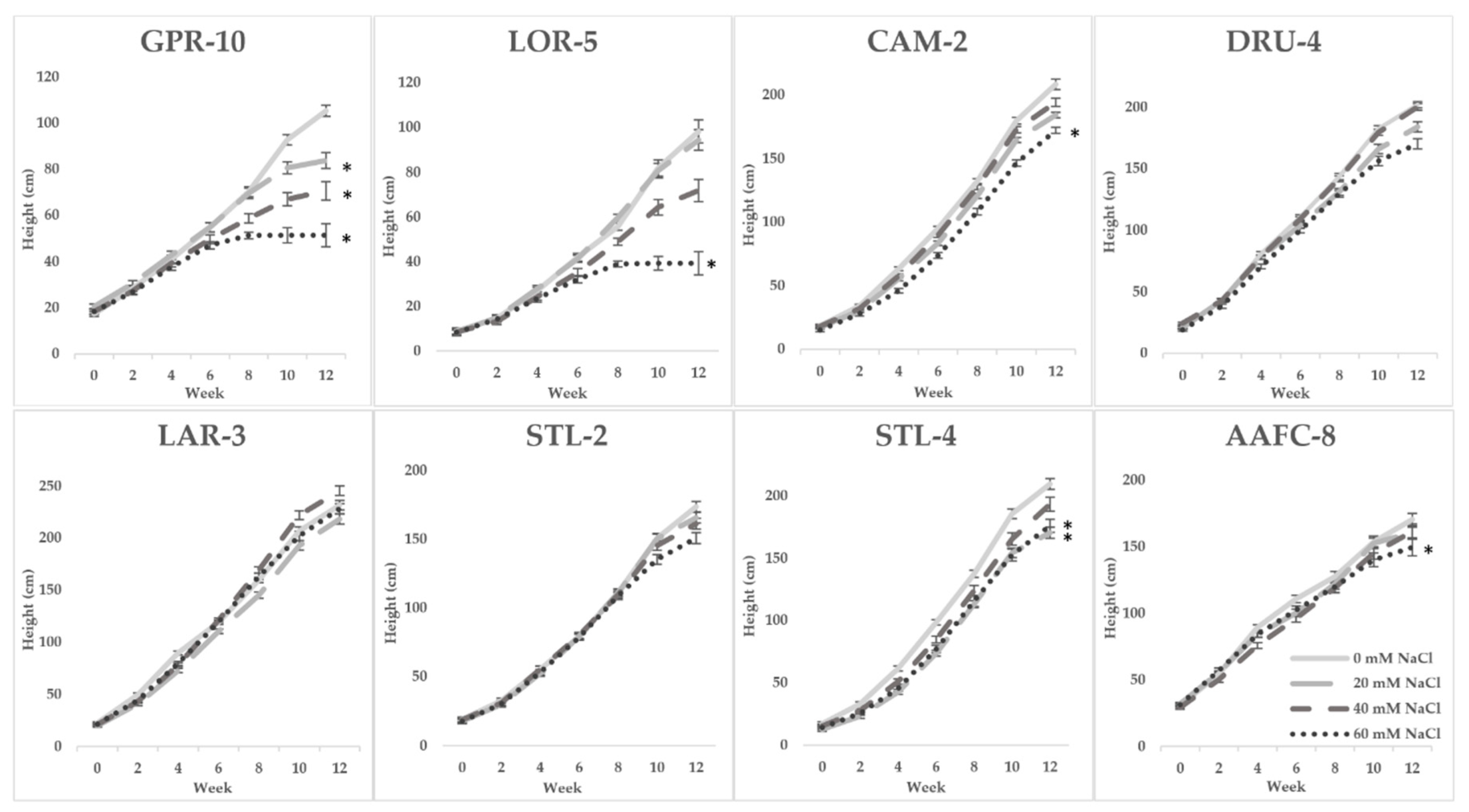
Survival is a crucial parameter in assessing the potential of a species for soil remediation. In this study, balsam poplar genotypes experienced 0% survival at or above 60 mM NaCl indicating that it is a relatively salt-intolerant species, while the comparatively salt-resistant willow genotypes displayed 93% survival following eight weeks treatment with 80 mM NaCl (7.0 mS
−1). Interestingly, a recent study using an eastern population of
Salix eriocephala reported a 40% survival rate after 25 days of 3 mS
−1 saline treatment [
24]. Differences between two populations suggest substantial genotypic variance, possibly arising from differing selective pressures between eastern and western Canadian genotypes, such as the prevalence of saline soils and drier climates characteristic of the Canadian prairies [
28].
The toxic effects of salinity on plants manifest primarily in stunting or cessation of growth, and decreased carbon assimilation [
29,
30]. In the second salinity trial, hybrid willow accumulated more biomass at all salinity treatments compared to other genotypes, accruing ~20% greater dry biomass in the control treatment than the next largest genotype (
Table 1). Additionally, AAFC-8 experienced no significant reduction in root biomass at either 20 or 40 mM NaCl compared to the corresponding control. High growth rates are advantageous for phytoremediation as absorbed salts can be diluted within plant tissues, mitigating toxic effects as well as providing a secondary product for industry [
31,
32]. Willow genotypes STL-2 and CAM-2 exhibited average dry biomass losses of only 32.6% and 39.9% at 60 mM NaCl, substantially lower than the 80% decreases observed in LAR-3 and DRU-4 when subject to similar treatments.
One of the largest factors resulting in losses in biomass is salt-induced necrosis and abscission of leaves, which physically releases biomass and limits carbon uptake [
33]. We observed salinity-induced senescence followed by the emergence of new leaves in the willow genotypes consistent with previous observations [
34]. This phenomenon likely contributed to the overall salt tolerance by shedding Na
+ laden leaves and replacing them with new, unaffected foliage, thus maintaining carbon assimilation.
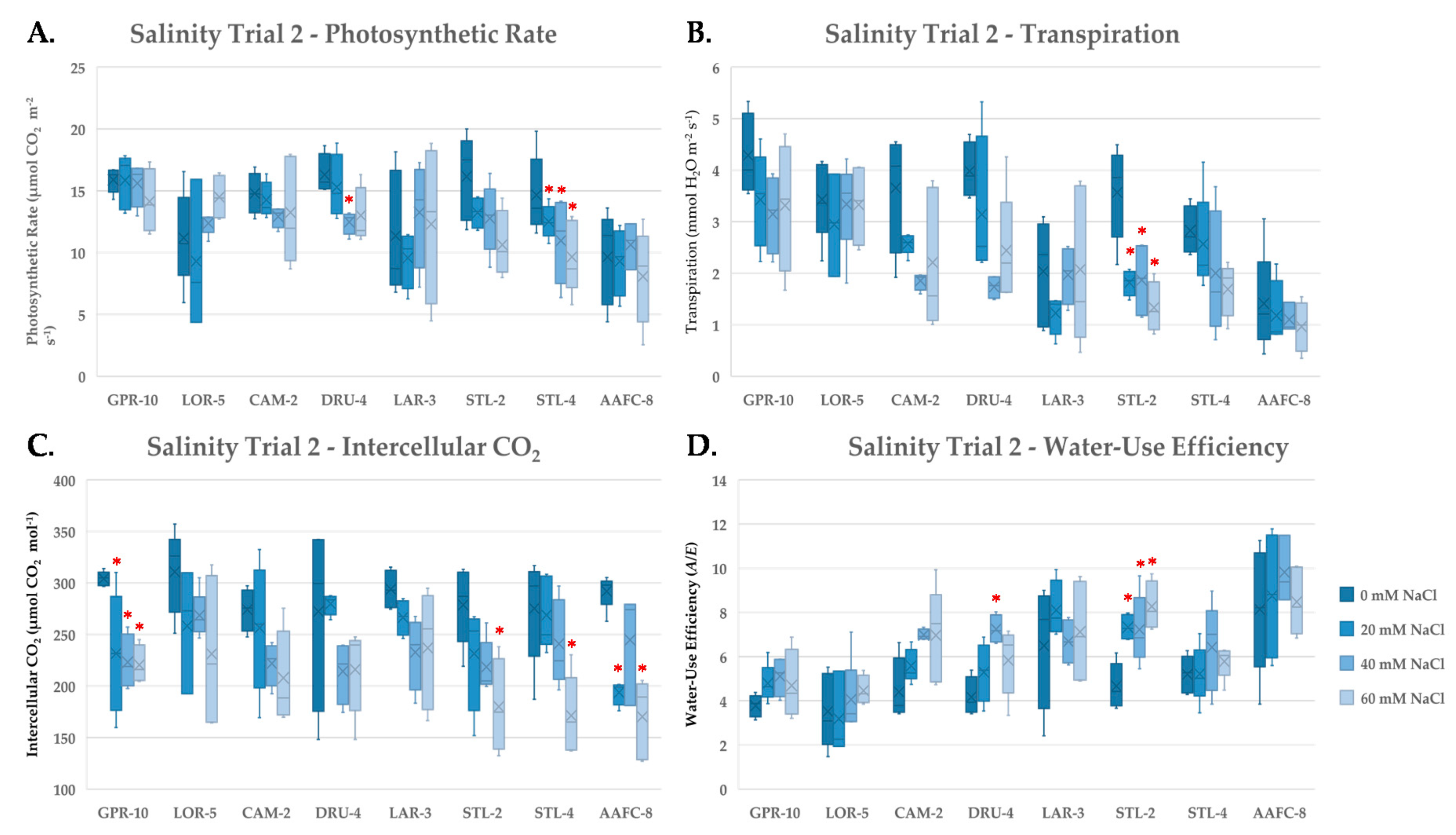
Photosynthesis is particularly susceptible to saline soils due to both toxic and osmotic effects. We observed a decrease in net photosynthesis with salinity treatment, consistent with previous findings [
32,
35,
36]. Stomatal closure is triggered via the ABA signalling pathway in the presence of saline soils in order to preserve water in unfavourable osmotic conditions [
37]. Wide variation in transpiration rate was observed with salinity treatment, ranging from a 2.9% decrease in GPR-10 to a 62.7% reduction in STL-2, the latter indicating considerable stomatal closure. We examined both transpiration and intercellular CO
2 concentrations, which can provide insight into the primary cause of the observed decrease in carbon assimilation [
38]. Intercellular CO
2 decreased in all genotypes with increasing salinity, indicating that reductions in stomatal conductance were more impactful than direct effects on photosynthetic capacity. Reduced transpiration, however, may be beneficial in concurrently limiting salt translocation to sensitive tissues [
39]. Willow genotypes CAM-2 and STL-2 both exhibited the strictest stomatal control and also exhibited the smallest decreases in biomass with salinity, suggesting that these two genotypes excelled at limiting water loss in saline conditions, and also restricted salts from accumulating in toxic quantities within their photosynthetic tissues.
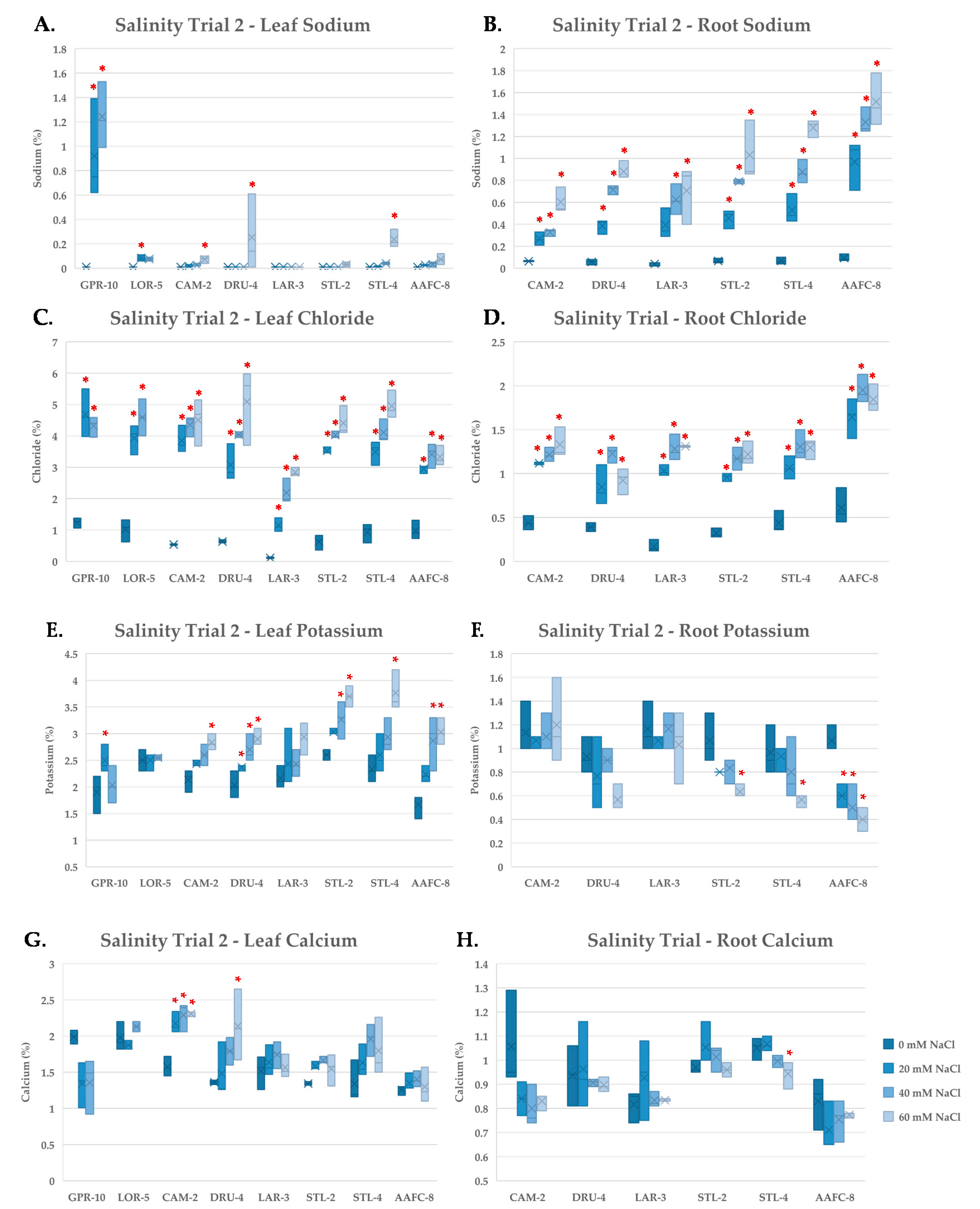
Sodium ions exert toxic effects through their competition with K
+ binding sites, degradation of membranes, evolution of reactive oxygen species, and inhibition of photosynthesis [
3,
40,
41,
42]. In the native and hybrid willow genotypes, Na
+ was limited in leaf tissues and accumulated primarily in root tissues. Balsam poplar genotype GPR-10 failed to restrict Na
+ movement to leaves at 20 mM NaCl and was also the most salt-sensitive genotype examined, however, native willow genotypes DRU-4 and STL-4 amassed Na
+ in their leaves at the severe salinity treatment and experienced significant biomass loss and senescence, suggesting that that control of Na
+ translocation is crucial for salt tolerance within these species.
Sodium and potassium are highly competitive for binding sites on membrane-bound proteins and cytosolic enzymes, and plants have evolved highly specialized systems to maintain a high K
+:Na
+ ratio [
43]. We observed an increase in K
+ concentration in leaves with increasing salinity in willow genotypes, but not in balsam poplar. Roots experienced a decline in K
+ with salinity, suggesting translocation to leaf tissues to help maintain metabolic function. A favourable K
+:Na
+ ratio was apparent in willow leaves, and likely contributes to normal enzyme function, preventing apoptosis and necrotic lesions from forming. Calcium was seen to increase in willow leaves, significantly at all salinity levels in the salt-tolerant CAM-2 genotype. Calcium acts as a secondary messenger in the salt stress response and contributes to membrane stabilization [
44,
45].
Chloride accumulated to levels as high as 5% by dry weight in leaves, and to far lower concentrations in roots. It has been demonstrated that Cl
− facilitates stomatal closure [
46]. Furthermore, it may act as a readily available ion used in internal osmotic adjustment for maintenance of turgor and facilitating water uptake. Chloride is toxic in tolerant species at tissues concentrations of 15–50 mg/g of dry weight, and was likely exerting toxic effects on the salt-treated trees [
47].
Plants can produce internal osmotic adjustments via synthesis of a wide variety of compatible solutes including sugars, sugar alcohols, and charged metabolites [
48]. We measured the concentrations of soluble carbohydrates in response to increasing salinity and found an increase of stachyose in leaf tissues and myo-inositol and raffinose in roots. Both stachyose and raffinose have been shown to act as osmoprotectants and free-radical scavengers, which may aid in anti-oxidant defense as well as promote normal enzyme functions [
49,
50]. It should be noted that the 357% increase in stachyose content may be misleading, as the concentrations measured were under 5mg per gram of tissue. Sucrose content was elevated in the roots of native willow genotypes, but not in balsam poplar, and may have contributed to tolerance via mild osmotic adjustments, via the production of stachyose and raffinose, and metabolization to yield energy [
51]. Despite the variety of metabolites examined in this study, little change was observed, suggesting that these metabolites likely contribute only marginally to overall salinity tolerance.
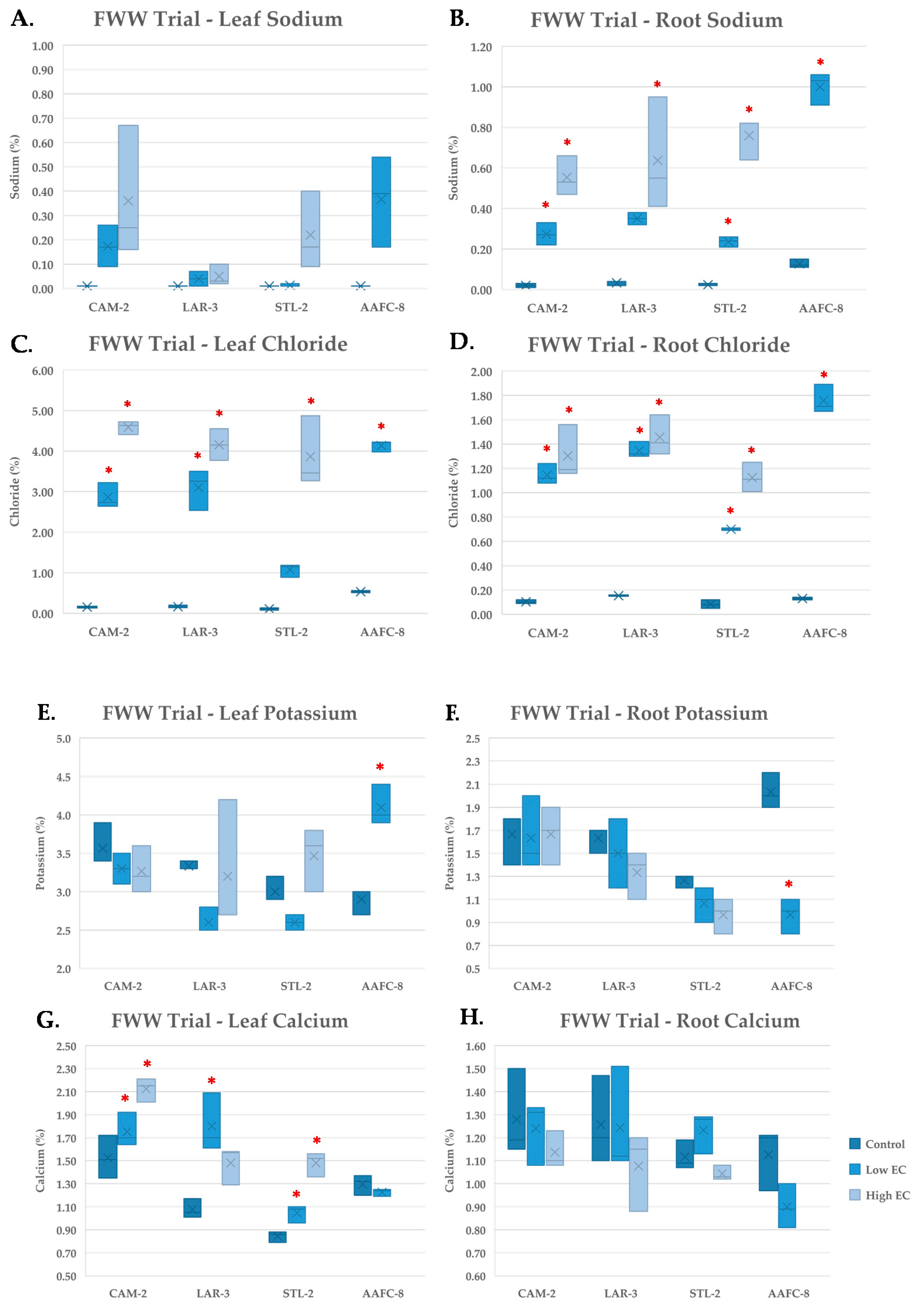
Four genotypes displayed a variety of traits that could be advantageous for industrial reclamation and phytoremediation: CAM-2, LAR-3, STL-2, and AAFC-8. Fracking wastewater is a complex mixture of highly concentrated salts, as well as surfactants, lubricants, and biocides, and the composition changes with industrial site and process [
52]. Subjecting these four tolerant genotypes to FWW treatments allowed us to not only test their growth on an industrial pollutant, but also observe and compare their physiological stress response to that of saline water.
Numerous differences were witnessed with FWW compared to salinity treatments. Despite good performance in the first two salinity trials, the hybrid willow genotype failed to survive after eight weeks of fracking wastewater treatment. This genotype experienced a 44.2% loss of total dry biomass at the low FWW treatment compared to a 25.8% decrease at salinity treatments with similar EC. Although the ECs of the severe FWW treatment and 60 mM NaCl solutions were similar, the actual Na
+ content of the severe FWW treatment was calculated to be 72.8 mM, which may be too toxic for the hybrid willow to withstand, or the other toxic constituents in the FWW may have contributed to its mortality. However, CAM-2, LAR-3, and STL-2 survived eight weeks of the severe FWW treatment and mirrored the growth of earlier salinity trials. Both CAM-2 and STL-2 exhibited a more pronounced reduction in biomass compared to the previous salinity trials, but also displayed fewer necrotic lesions. This suggests that energy that may normally be used for growth could instead be allocated to active transport processes and antioxidant defenses [
53].
Net photosynthetic rates were heavily reduced in all genotypes with FWW treatment, except in LAR-3. Negative photosynthetic rates were recorded in STL-2 indicating that respiration outpaced carbon assimilation, and supports the idea that energy is being allocated to active stress responses, or that additional stressors are interfering with metabolic processes [
54]. Transpiration paralleled the previous salinity trials, but WUE increased and intercellular CO
2 was unchanged between FWW treatments. This indicates that there was no significant stomatal restriction to CO
2, rather the stressors in the FWW directly impacted the photosynthetic capacity [
53,
55,
56].
Sodium and chloride both followed analogous patterns of accumulation in leaf and root tissues treated with FWW compared with the previous salinity trial. The hybrid willow accumulated more leaf Na+ at the low FWW treatment than it had under the previous 60 mM NaCl treatment, indicating that the additional stressors present in the FWW may be disrupting normal Na+ compartmentalization. Potassium remained unchanged in response to FWW treatments but was present in higher concentrations than in the salinity trials, likely due its presence in the FWW in addition to the applied fertilizer. Calcium also remained unchanged despite its greater abundance in the FWW solution, suggesting that it may exist in a biologically unavailable form. Copper, lead, and nickel were found to be present in low concentrations in the stock FWW solution, but were not found in detectable quantities in the plant tissues examined.
A growing body of work has documented the production of antioxidants, phytochelators, and various charged metabolites in response to salinity stresses, but non-structural carbohydrates have been poorly studied. Roots accumulated sucrose in response to FWW treatment which may be used to fuel respiration and provide ATP for active transport and production of other osmolytes [
51,
54]. Fructose and raffinose both decreased in leaf tissues with FWW treatment, but were unaffected in the salinity trials; similar decreases were found in cucumbers grown near an industrial copper production site [
57]. The willow hybrid exhibited a 50% decrease in myo-inositol and glucose as well as an 80% drop in raffinose with the low FWW treatment, while CAM-2 and STL-2 maintained greater carbohydrate concentrations at the severe FWW treatment.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






