Geographical Patterns of Algal Communities Associated with Different Urban Lakes in China

Abstract
:1. Introduction
2. Materials and Methods
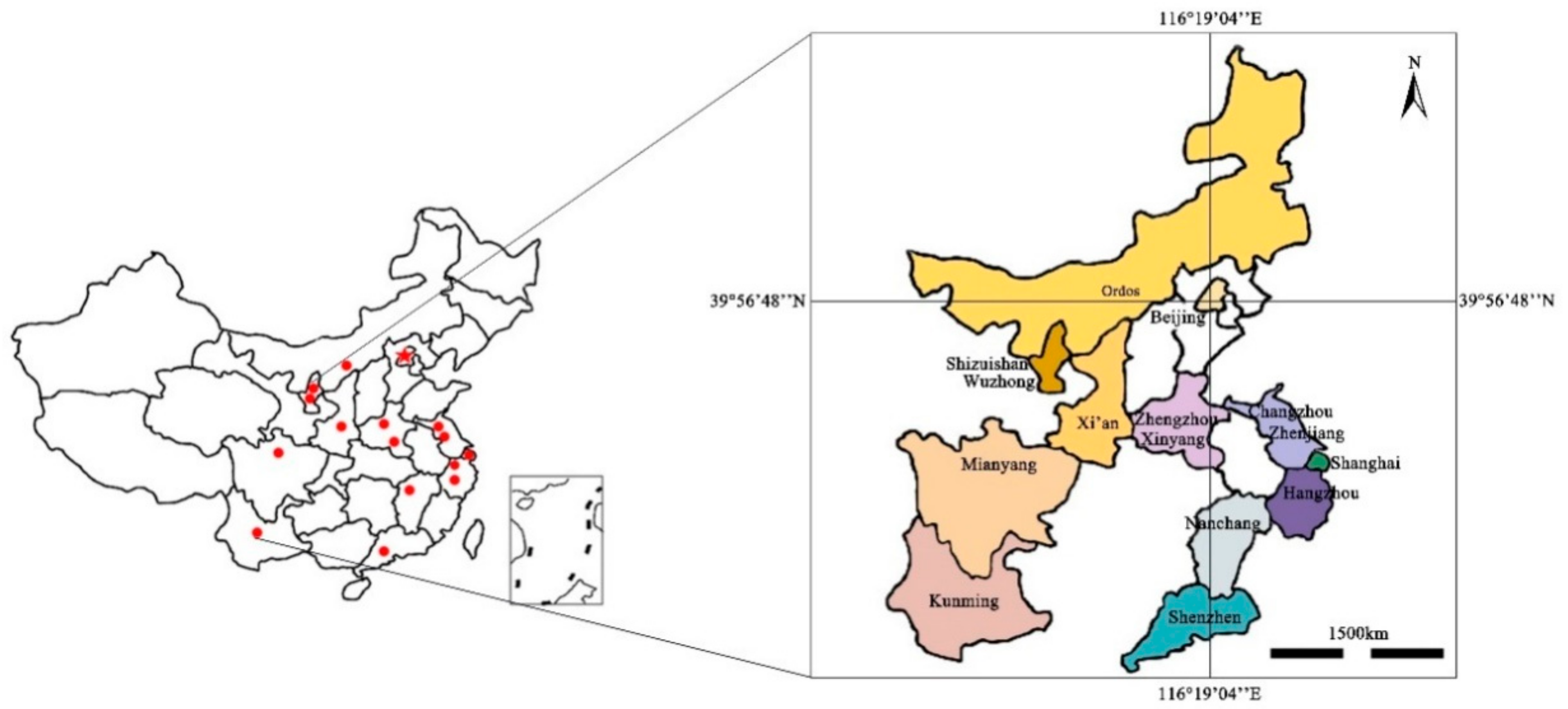
2.1. Site Description and Field Sampling
2.2. Water Physicochemical Analysis
2.3. Algal Cell Concentration and Community
2.4. Network Construction
2.5. Statistical Analysis
3. Results and Discussion
3.1. Water Quality Parameters
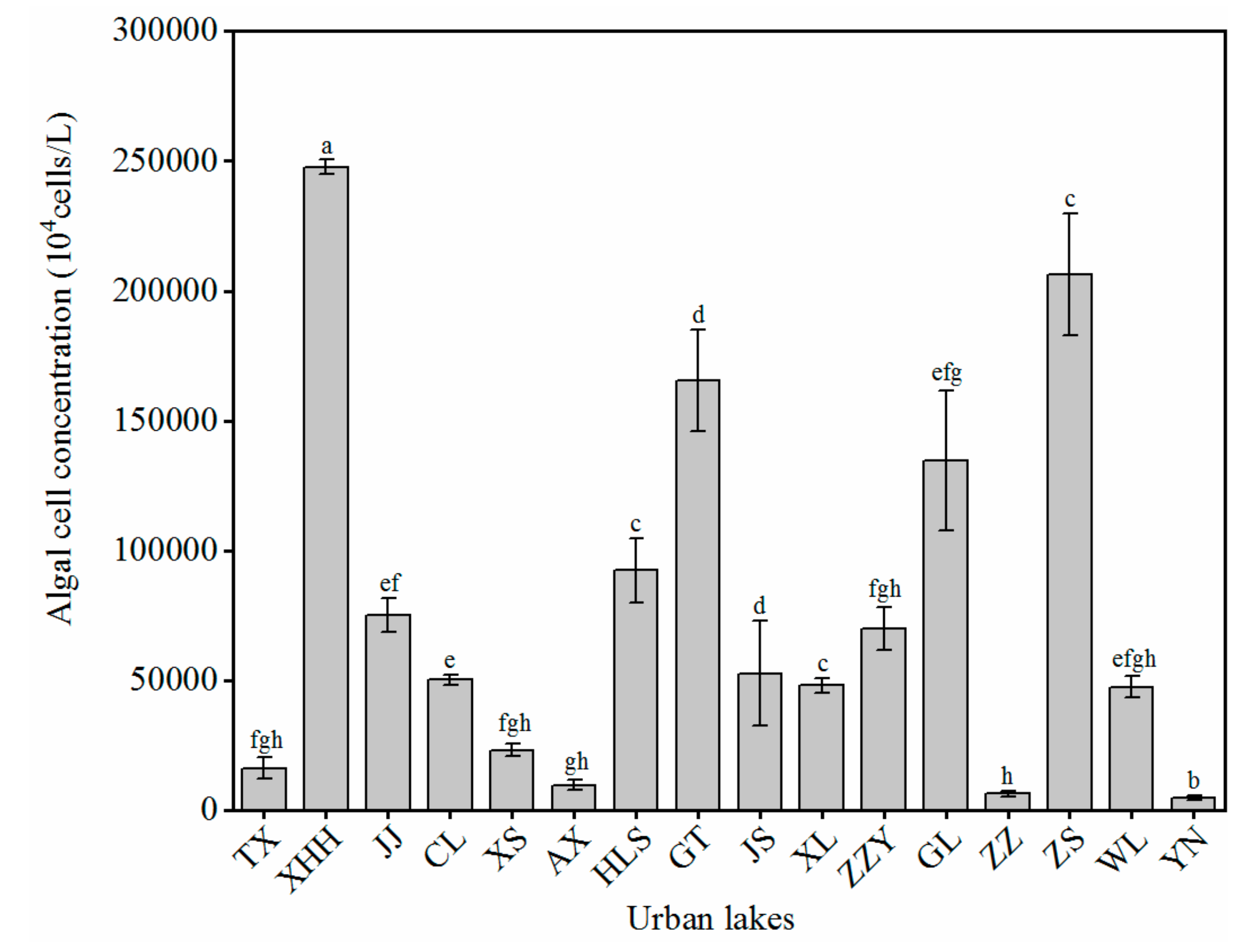
3.2. Algal Cell Concentration
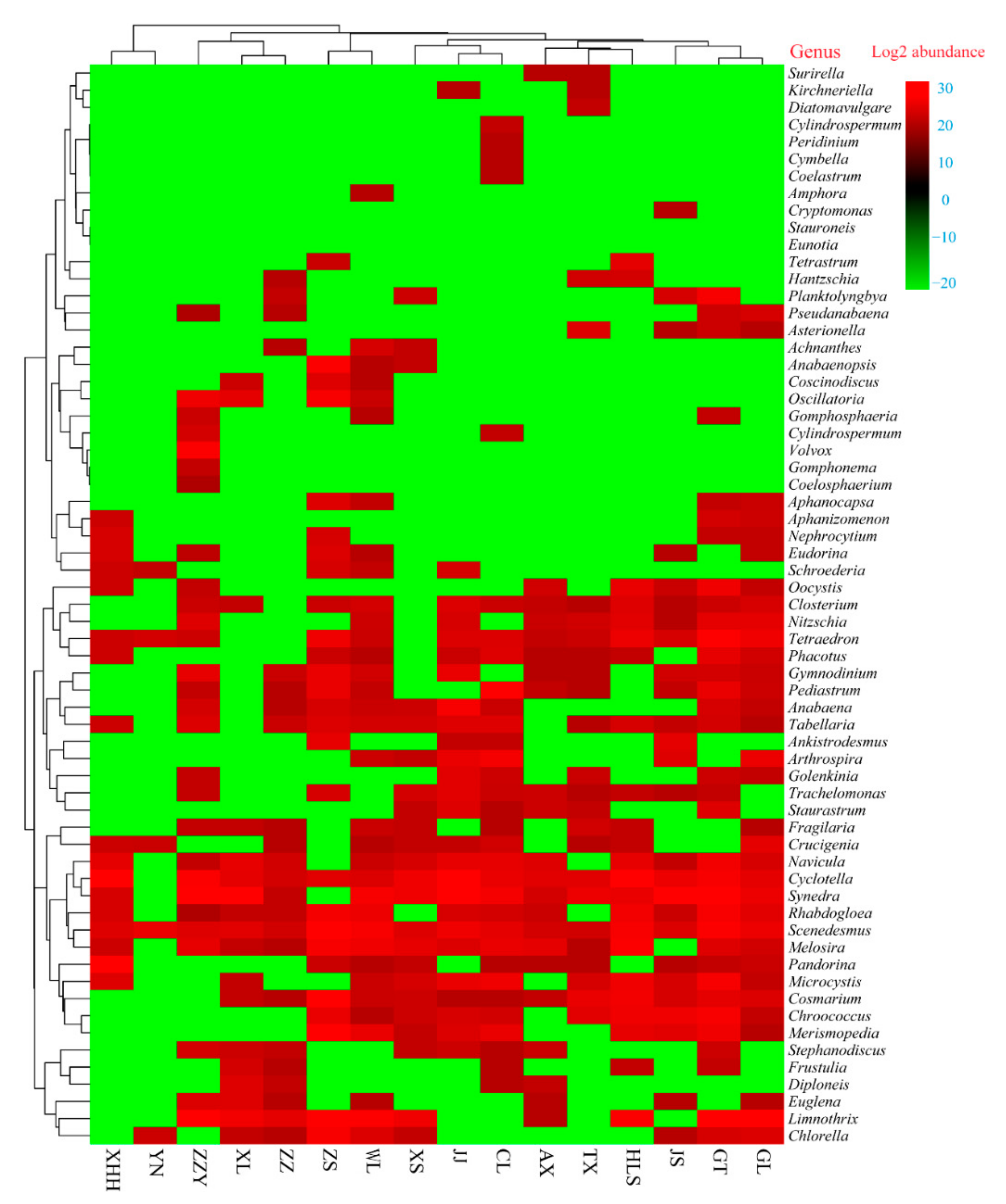
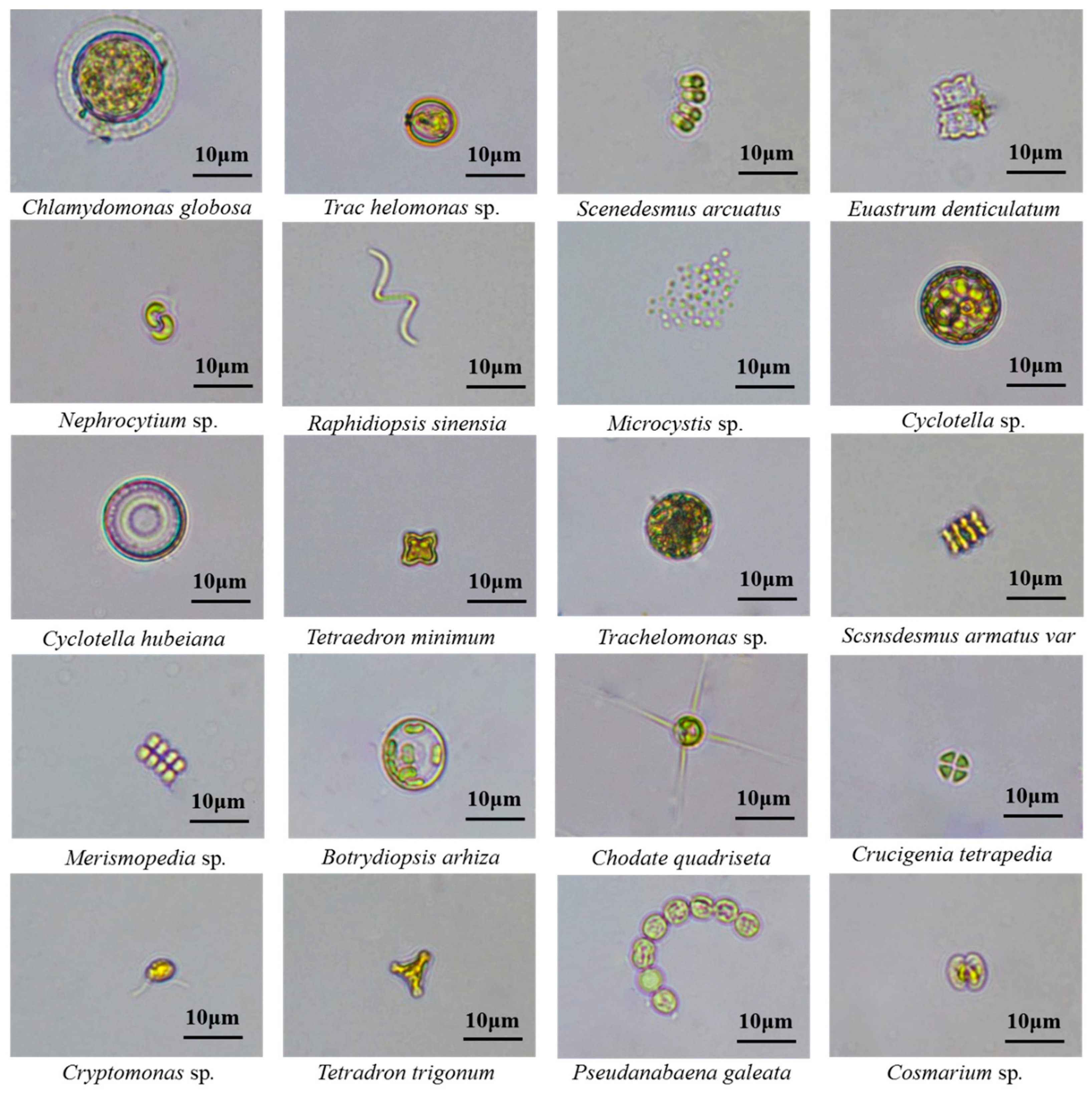
3.3. Geographical Patterns of Algal Community Composition and Typical Cell Morphology
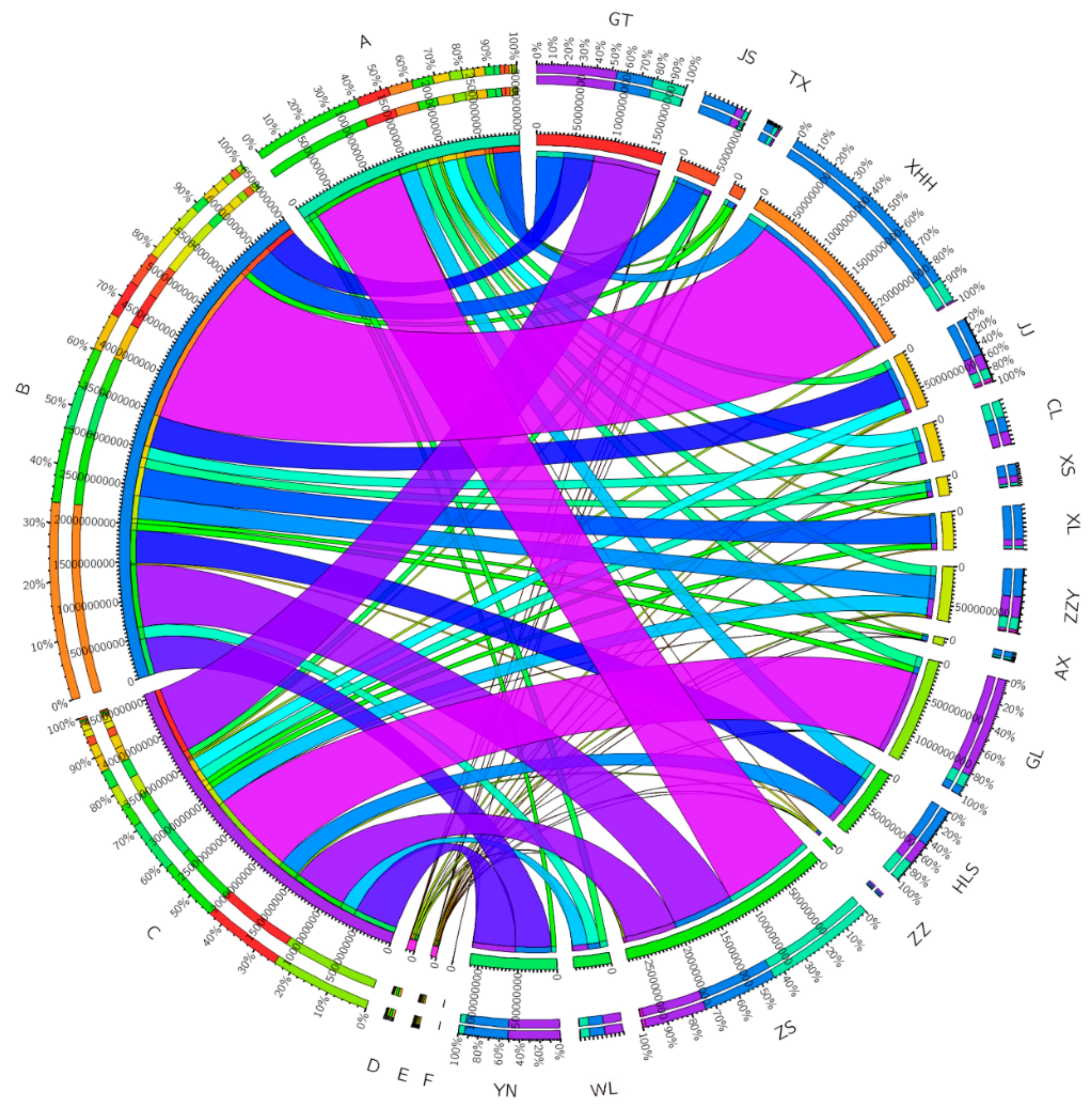
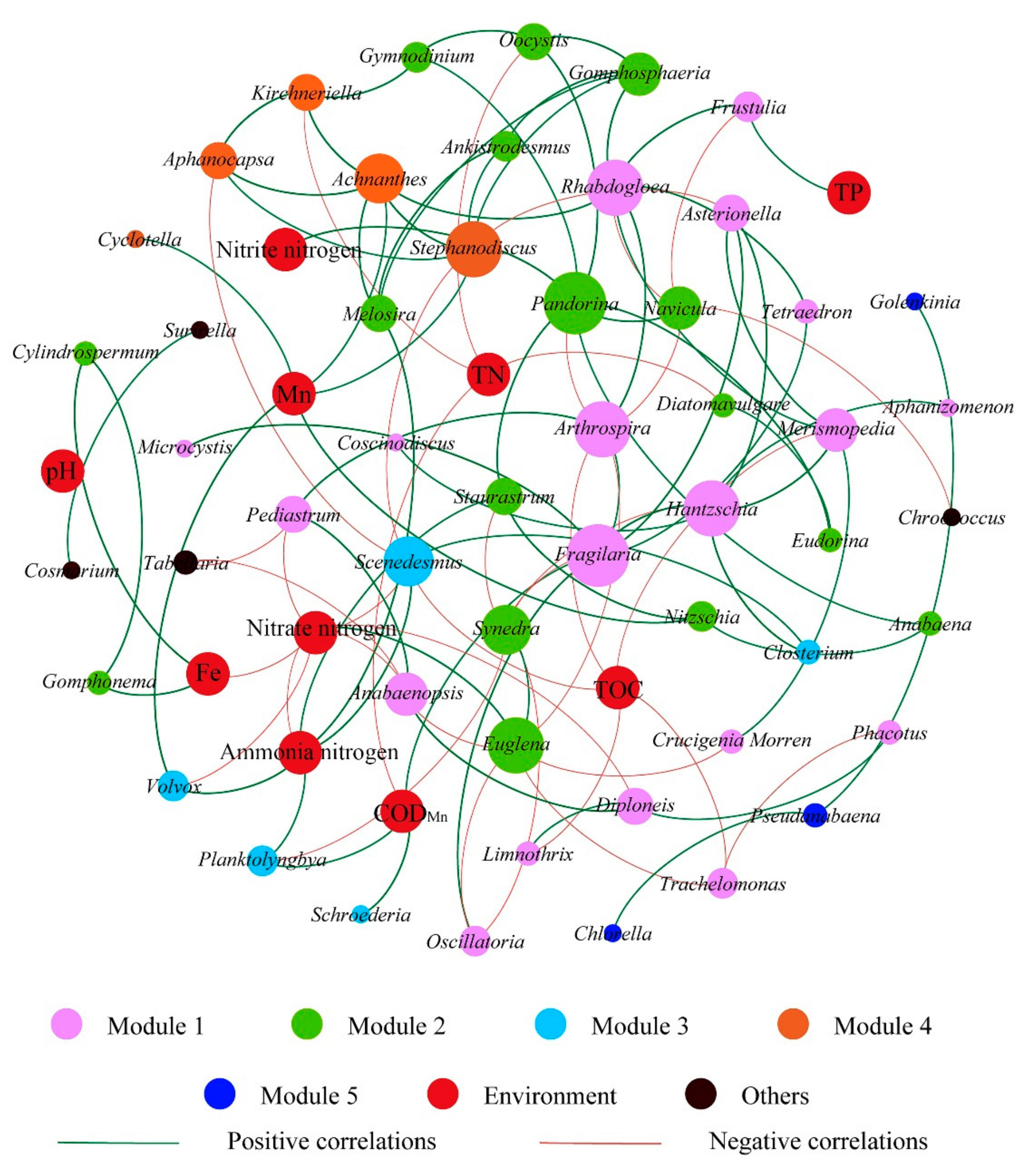
3.4. Co-Occurrence Network of Algae
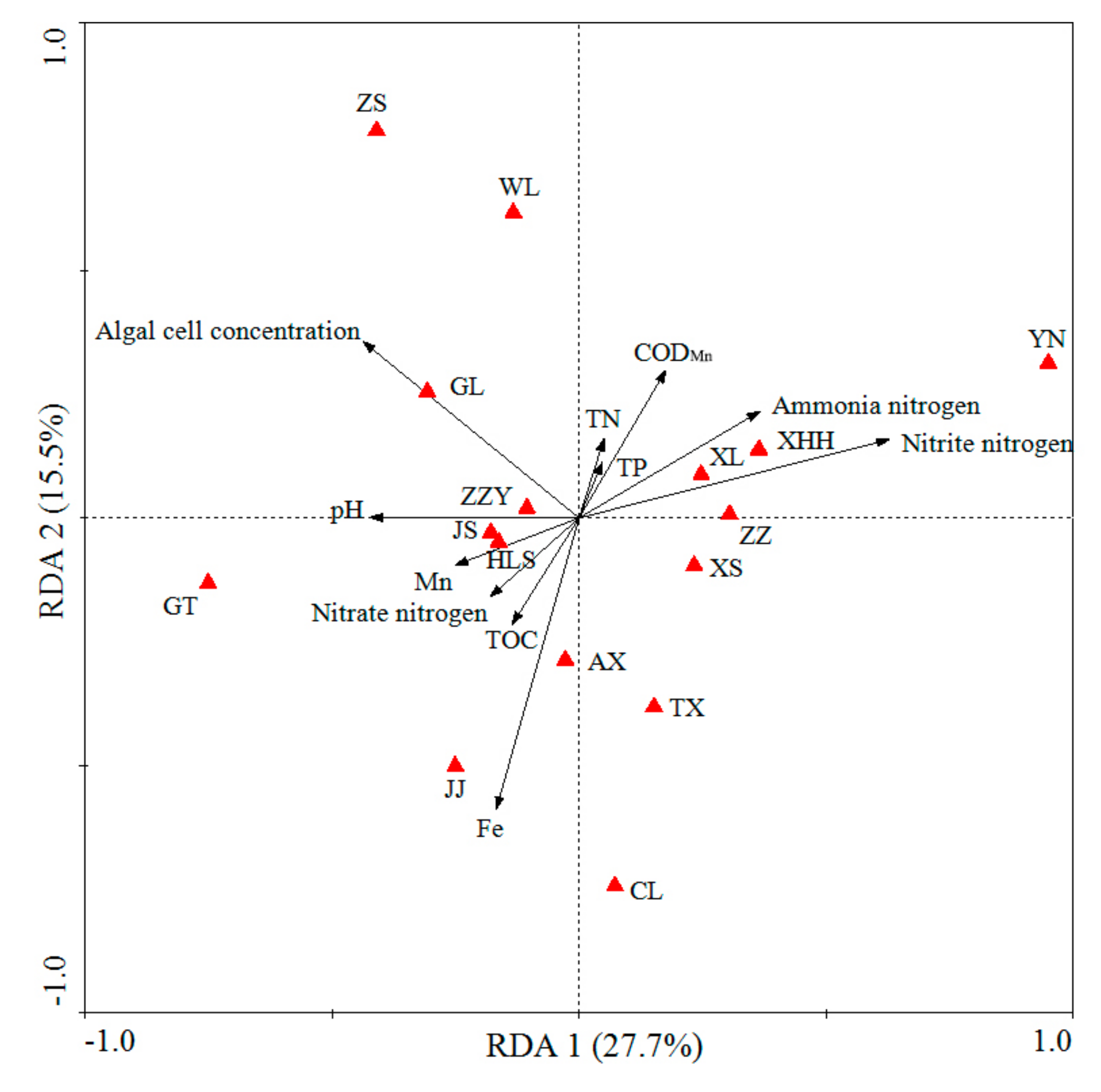
3.5. Relationship between Algal Communities and Water Quality
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Garner, E.; Inyang, M.; Garvey, E.; Parks, J.; Glover, C.; Grimaldi, A.; Dickenson, E.; Sutherland, J.; Salveson, A.; Edwards, M.A.; et al. Impact of blending for direct potable reuse on premise plumbing microbial ecology and regrowth of opportunistic pathogens and antibiotic resistant bacteria. Water Res. 2019, 151, 75–86. [Google Scholar] [CrossRef]
- Su, J.F.; Gao, C.Y.; Huang, T.L.; Gao, Y.C.; Bai, X.C.; He, L. Characterization and mechanism of the Cd(II) removal by anaerobic denitrification bacterium Pseudomonas sp. H117. Chemosphere 2019, 222, 970–979. [Google Scholar] [CrossRef]
- Yang, J.; Yang, Y.; Chen, R.S.; Meng, X.Z.; Xu, J.; Qadeer, A.; Liu, M. Modeling and evaluating spatial variation of polycyclic aromatic hydrocarbons in urban lake surface sediments in Shanghai. Environ. Pollut. 2018, 235, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Metson, G.S.; Lin, J.; Harrison, J.A.; Compton, J.E. Linking terrestrial phosphorus inputs to riverine export across the United States. Water Res. 2017, 124, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Lompe, K.M.; Mohseni, M.; Bérubé, P.R.; Sauvé, S.; Barbeau, B. Biological ion exchange as an alternative to biological activated carbon for drinking water treatment. Water Res. 2020, 168, 115148. [Google Scholar] [CrossRef] [PubMed]
- Eckert, E.M.; Di Cesare, A.; Kettner, M.T.; Arias-Andres, M.; Fontaneto, D.; Grossart, H.-P.; Corno, G. Microplastics increase impact of treated wastewater on freshwater microbial community. Environ. Pollut. 2018, 234, 495–502. [Google Scholar] [CrossRef]
- Jeppesen, E.; Kronvang, B.; Meerhoff, M.; Sondergaard, M.; Hansen, K.M.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Beklioglu, M.; Ozen, A.; et al. Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J. Environ. Qual. 2009, 38, 1930–1941. [Google Scholar] [CrossRef]
- Yin, Q.; Fu, B.; Li, B.; Shi, X.; Inagaki, F.; Zhang, X.H. Spatial variations in microbial community composition in surface seawater from the Ultra-Oligotrophic center to rim of the south Pacific Gyre. PLoS ONE 2013, 8, e55148. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Yan, Q.; Feng, W. Spatiotemporal heterogeneity of plankton communities in Lake Donghu, China, as revealed by PCR–denaturing gradient gel electrophoresis and its relation to biotic and abiotic factors. FEMS Microbiol. Ecol. 2008, 63, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Scherer, P.I.; Millard, A.D.; Miller, A.; Schoen, R.; Raeder, U.; Geist, J.; Zwirglmaier, K. Temporal dynamics of the microbial community composition with a focus on toxic cyanobacteria and toxin presence during harmful algal blooms in two south German Lakes. Front. Microbiol. 2017, 8, 2387. [Google Scholar] [CrossRef]
- Sabater, S. Diatom communities as indicators of environmental stress in the Guadiamar River, S-W. Spain, following a major mine tailings spill. J. Appl. Phycol. 2000, 12, 113–124. [Google Scholar] [CrossRef]
- Jelena, R.N.; Henner, H. Phytoplankton community and chlorophyll a as trophic state indices of Lake Skadar (Montenegro, Balkan). Environ. Sci. Pollut. Res. 2005, 3, 146–152. [Google Scholar]
- Su, X.; Steinman, A.D.; Tang, X.; Xue, Q.; Zhao, Y.; Xie, L. Response of bacterial communities to cyanobacterial harmful algal blooms in Lake Taihu, China. Harmful Algae 2017, 68, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.G.; Li, A.J.; Qin, J.; Li, Q.; Ho, J.G.; Li, H.S. Seasonal patterns of water quality and phytoplankton dynamics in surface waters in Guangzhou and Foshan, China. Sci. Total Environ. 2017, 590–591, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yang, J.; Yu, X.; Chen, G.; Yu, Z. Patterns in the composition of microbial communities from a subtropical river: Effects of environmental, spatial and temporal factors. PLoS ONE 2013, 8, e81232. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.J.; He, W.; Liu, W.X.; Qin, N.; Ouyang, H.L.; Wang, Q.M.; Kong, X.Z.; He, Q.S.; Yang, C.; Yang, B.; et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu). Ecol. Indic. 2014, 40, 58–67. [Google Scholar] [CrossRef]
- Dodds, W.K.; Smith, V.H.; Lohman, K. Nitrogen and phosphorus relationships to benthic algal biomass in temperate streams. Can. J. Fish. Aquat. Sci. 2002, 59, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Peura, S.; Eiler, A.; Hiltunen, M.; Nykanen, H.; Tiirola, M.; Jones, R.I. Bacterial and phytoplankton responses to nutrient amendments in a boreal lake differ according to season and to taxonomic resolution. PLoS ONE 2012, 7, e38552. [Google Scholar] [CrossRef]
- Vakkilainen, K.; Kairesalo, T.; Hietala, J.; Balayla, D.M.; Becares, E.; Van de Bund, W.J.; Van Donk, E.; Fernandez-Alaez, M.; Gyllstrom, M.; Hansson, L.A.; et al. Response of zooplankton to nutrient enrichment and fish in shallow lakes: A pan-European mesocosm experiment. Freshw. Biol. 2004, 49, 1619–1632. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.H.; Chen, S.N.; Huang, T.L.; Ma, W.X.; Xu, J.L.; Sun, X. Vertical distribution of bacterial community diversity and water quality during the Reservoir thermal stratification. Int. J. Environ. Res. Public Health 2015, 12, 6933–6945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulshrestha, S.K.; Adholia, U.N.; Khan, A.A.; Bhatnagar, A.; Saxena, M.; Baghail, M. Community structure of planktons and macrozoobenthos, with special reference to pollution in the River Khan (India). Int. J. Environ. Stud. 1989, 35, 83–96. [Google Scholar] [CrossRef]
- Zhang, H.H.; Chen, S.N.; Huang, T.L.; Shang, P.L.; Yang, X.; Ma, W.X. Indoor heating drives water bacterial growth and community metabolic profile changes in building tap pipes during the winter season. Int. J. Environ. Res. Public Health 2015, 12, 13649–13661. [Google Scholar] [CrossRef] [PubMed]
- Diehl, S.; Berger, S.; Ptacnik, R.; Wild, A. Phytoplankton, light, and nutrients in a gradient of mixing depths: Field experiments. Ecology 2002, 83, 399–411. [Google Scholar] [CrossRef]
- Maestrini, S.Y.; Berland, B.R.; Bréret, M.; Béchemin, C.; Poletti, R.; Rinaldi, A. Nutrients limiting the algal growth potential (AGP) in the Po River plume and an adjacent area, Northwest Adriatic Sea: Enrichment bioassays with the test algaeNitzschia closterium and Thalassiosira pseudonana. Estuaries 1997, 20, 416–429. [Google Scholar] [CrossRef]
- Habib, O.A.; Tippett, R.; Murphy, K.J. Seasonal changes in phytoplankton community structure in relation to physico-chemical factors in Loch Lomond, Scotland. Hydrobiologia 1997, 350, 63–79. [Google Scholar] [CrossRef]
- Zhang, H.H.; Feng, J.; Chen, S.N.; Zhao, Z.F.; Li, B.Q.; Wang, Y.; Jia, J.Y.; Li, S.L.; Wang, Y.; Yan, M.M.; et al. Geographical patterns of nirS gene abundance and nirS-Type denitrifying bacterial community associated with activated sludge from different wastewater treatment plants. Microb. Ecol. 2019, 77, 304–316. [Google Scholar] [CrossRef]
- Liu, L.; Chen, H.; Liu, M.; Yang, J.R.; Xiao, P.; Wilkinson, D.M.; Yang, J. Response of the eukaryotic plankton community to the cyanobacterial biomass cycle over 6 years in two subtropical reservoirs. ISME J. 2019, 13, 2196–2208. [Google Scholar] [CrossRef] [Green Version]
- Berdjeb, L.; Parada, A.; Needham, D.M.; Fuhrman, J.A. Short-term dynamics and interactions of marine protist communities during the spring–summer transition. ISME J. 2018, 12, 1907–1917. [Google Scholar] [CrossRef]
- Morrien, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; de Hollander, M.; Soto, R.L.; Bouffaud, M.L.; Buee, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, Y.; Luo, F.; He, Z.; Tu, Q.; Zhi, X. Functional molecular ecological networks. mBio 2010, 1, e00169-10–e00169-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bascompte, J. Networks in ecology. Basic Appl. Ecol. 2007, 8, 485–490. [Google Scholar] [CrossRef]
- Montoya, J.M.; Pimm, S.L.; Solé, R.V. Ecological networks and their fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; He, H.Y.; Chen, S.N.; Huang, T.T.; Lu, K.Y.; Zhang, Z.H.; Wang, R.; Zhang, X.Y.; Li, H.L. Abundance of antibiotic resistance genes and their association with bacterial communities in activated sludge of wastewater treatment plants: Geographical distribution and network analysis. J. Environ. Sci. 2019, 82, 24–38. [Google Scholar] [CrossRef]
- Kang, P.L.; Hang, T.L.; Zhang, H.H.; Chen, S.N.; Shang, P.L.; Feng, J.; Jia, J.Y. Water quality and diversity of denitrifier community structure of typical scenic water bodies in Xi’an. Environ. Sci. Pollut. Res. 2017, 38, 5174–5183. [Google Scholar]
- Shi, J.C.; Huang, T.L.; Wen, G.; Liu, F.; Qiu, X.P.; Wang, B.S. The variation characteristic of sulfides and VOSc in a source water reservoir and its control using a water-lifting aerator. Int. J. Environ. Res. Public Health 2016, 13, 427. [Google Scholar] [CrossRef] [Green Version]
- Nazeer, M.; Wong, M.S.; Nichol, J.E. A new approach for the estimation of phytoplankton cell counts associated with algal blooms. Sci. Total Environ. 2017, 590–591, 125–138. [Google Scholar] [CrossRef]
- Shen, Y.R.; Zhang, Z.S.; Gong, X.J.; Gu, M.R.; Shi, Z.X.; Wei, Y.X. Modern Biomonitoring Techniques Using Freshwater Microbiota; China Architecture and Building Press: Beijing, China, 1990. [Google Scholar]
- Zhang, Z.S.; Huang, X.F. Research Methods of Freshwater Plankton; Science Press: Beijing, China, 1991. [Google Scholar]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China; Science and Technology Press: Beijing, China, 2006. [Google Scholar]
- Newman, M.E.J. Communities, modules and large-scale structure in networks. Nat. Phys. 2012, 8, 25–31. [Google Scholar] [CrossRef]
- Sun, W.; Krumins, V.; Dong, Y.; Gao, P.; Ma, C.; Hu, M.; Li, B.; Xia, B.; He, Z.; Xiong, S. A combination of stable isotope probing, Illumina Sequencing, and co-occurrence network to investigate thermophilic Acetate and lactate utilizing bacteria. Microb. Ecol. 2018, 75, 113–122. [Google Scholar] [CrossRef]
- Blondel, V.D.; Guillaume, J.L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. Theory Equip. 2008, 2008, 10008. [Google Scholar] [CrossRef] [Green Version]
- Jacomy, M.; Venturini, T.; Heymann, S.; Bastian, M. ForceAtlas2, a continuous graph layout algorithm for handy network visualization designed for the Gephi Software. PLoS ONE 2014, 9, e98679. [Google Scholar] [CrossRef] [PubMed]
- Beisser, D.; Klau, G.W.; Dandekar, T.; Müller, T.; Dittrich, M.T. Binet: An R-Package for the functional analysis of biological networks. Bioinformatics 2010, 26, 1129–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parulekar, N.; Kolekar, P.; Jenkins, A.; Kleiven, S.; Utkilen, H.; Johansen, A.; Sawant, S.; Kulkarni-Kale, U.; Kale, M.; Sæbø, M. Characterization of bacterial community associated with phytoplankton bloom in a eutrophic lake in South Norway using 16S rRNA gene amplicon sequence analysis. PLoS ONE 2017, 12, e0173408. [Google Scholar] [CrossRef]
- Legendre, P.; Oksanen, J.; ter Braak, C.; Legendre, P.; Oksanen, J.; Braak, C.J.F. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Yang, J.; Jiang, H.; Wu, G.; Liu, W.; Zhang, G. Distinct factors shape aquatic and sedimentary microbial community structures in the lakes of western China. Front. Microbiol. 2016, 7, 1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.; Liu, Y.; Lin, X.; Zhang, H.; Zeng, J.; Hou, J.; Yang, Y.; Yao, T.; Knight, R.; Chu, H. Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environ. Microbiol. 2012, 14, 2457–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logares, R.; Lindstrom, E.S.; Langenheder, S.; Logue, J.B.; Paterson, H.; Laybourn-Parry, J.; Rengefors, K.; Tranvik, L.; Bertilsson, S. Biogeography of bacterial communities exposed to progressive long-term environmental change. ISME J. 2013, 7, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Souffreau, C.; Van der Gucht, K.; van Gremberghe, I.; Kosten, S.; Lacerot, G.; Lobao, L.M.; de Moraes Huszar, V.L.; Roland, F.; Jeppesen, E.; Vyverman, W.; et al. Environmental rather than spatial factors structure bacterioplankton communities in shallow lakes along a > 6000 km latitudinal gradient in South America. Environ. Microbiol. 2015, 17, 2336–2351. [Google Scholar] [CrossRef]
- Crab, R.; Avnimelech, Y.; Defoirdt, T.; Bossier, P.; Verstraete, W. Nitrogen removal techniques in aquaculture for a sustainable production. Aquaculture 2007, 270, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Tan, H.; Luo, G.; Liang, W. The effects of Bacillus subtilis on nitrogen recycling from aquaculture solid waste using heterotrophic nitrogen assimilation in sequencing batch reactors. Bioresour. Technol. 2012, 124, 180–185. [Google Scholar] [CrossRef]
- Green, C.T.; Bekins, B.A.; Kalkhoff, S.J.; Hirsch, R.M.; Liao, L.; Barnes, K.K. Decadal surface water quality trends under variable climate, land use, and hydrogeochemical setting in Iowa, USA. Water Resour. Res. 2014, 50, 2425–2443. [Google Scholar] [CrossRef]
- Sarier, N. Specific features of adsorption of azo dyes on fly ash. Russ. Chem. Bull. 2007, 56, 566–569. [Google Scholar] [CrossRef]
- Andersson, M.G.I.; Berga, M.; Lindström, E.S.; Langenheder, S. The spatial structure of bacterial communities is influenced by historical environmental conditions. Ecology 2014, 95, 1134–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, G.P.; Shang, J.C. Spatial and temporal variations of eutrophication in western Chaohu Lake, China. Environ. Monit. Assess. 2007, 130, 99–109. [Google Scholar] [CrossRef]
- Yin, K.D.; Qian, P.Y.; Wu, M.C.S.; Chen, J.C.; Huang, L.M.; Song, X.Y.; Jian, W.J. Shift from P to N limitation of phytoplankton growth across the Pearl River estuarine plume during summer. Mar. Ecol. Prog. Ser. 2001, 221, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Huber, V.; Adrian, R.; Gerten, D. Phytoplankton response to climate warming modified by trophic state. Limnol. Oceanogr. 2008, 53, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Greenaway, A.M.; Gordon-Smith, D.A. The effects of rainfall on the distribution of inorganic nitrogen and phosphorus in Discovery Bay, Jamaica. Limnol. Oceanogr. 2006, 51, 2206–2220. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yu, X.; Liu, L.; Zhang, W.; Guo, P. Algae community and trophic state of subtropical reservoirs in southeast Fujian, China. Environ. Sci. Pollut. Res. 2012, 19, 1432–1442. [Google Scholar] [CrossRef] [Green Version]
- Moustaka-Gouni, M.; Vardaka, E.; Tryfon, E. Phytoplankton species succession in a shallow Mediterranean lake (L. Kastoria, Greece): Steady-state dominance of Limnothrix redekei, Microcystis aeruginosa and Cylindrospermopsis raciborskii. Hydrobiologia 2007, 575, 129–140. [Google Scholar] [CrossRef]
- Atkins, R.; Rose, T.; Brown, R.S.; Robb, M. The Microcystis cyanobacteria bloom in the Swan River February 2000. Water Sci. Technol. 2001, 43, 107–114. [Google Scholar] [CrossRef]
- Wagner, C.; Adrian, R. Cyanobacteria dominance: Quantifying the effects of climate change. Limnol. Oceanogr. 2009, 54, 2460–2468. [Google Scholar] [CrossRef]
- Carvalho, L.; Miller, C.A.; Scott, E.M.; Codd, G.A.; Davies, P.S.; Tyler, A.N. Cyanobacterial blooms: Statistical models describing risk factors for national-scale lake assessment and lake management. Sci. Total Environ. 2011, 409, 5353–5358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, H.B.; Lee, Y.J.; Lee, B.D.; Knappe, D.R.U. Effectiveness of coagulants and coagulant aids for the removal of filter-clogging Synedra. J. Water Supply Res. 2001, 50, 135–148. [Google Scholar] [CrossRef]
- Henderson, R.; Parsons, S.A.; Jefferson, B. The impact of algal properties and pre-oxidation on solid–liquid separation of algae. Water Res. 2008, 42, 1827–1845. [Google Scholar] [CrossRef] [PubMed]
- Bracht, B.B.; Stone, J.R.; Fritz, S.C. A diatom record of late Holocene climate variation in the northern range of Yellowstone National Park, USA. Quatern. Int. 2008, 188, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Saros, J.E.; Anderson, N.J. The ecology of the planktonic diatom Cyclotella and its implications for global environmental change studies. Biol. Rev. 2015, 90, 522–541. [Google Scholar] [CrossRef]
- Barberan, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.H.; Zhao, Z.F.; Chen, S.N.; Wang, Y.; Feng, J.; Jia, J.Y.; Kang, P.L.; Li, S.L. Geographical patterns of denitrifying bacterial communities associated with different urban lakes. RSC Adv. 2018, 8, 17079–17090. [Google Scholar] [CrossRef] [Green Version]
- Figueredo, C.C.; von Rückert, G.; Cupertino, A.; Pontes, M.A.; Fernandes, L.A.; Ribeiro, S.G.; Maran, N.R.C. Lack of nitrogen as a causing agent of Cylindrospermopsis raciborskii intermittent blooms in a small tropical reservoir. FEMS Microbiol. Ecol. 2014, 87, 557–567. [Google Scholar] [CrossRef] [Green Version]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calijuri, M.C.; Dos Santos, A.C.A.; Jati, S. Temporal changes in the phytoplankton community structure in a tropical and eutrophic reservoir (Barra Bonita, S.P.—Brazil). J. Plankton Res. 2002, 24, 617–634. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Yang, J.; Lv, H.; Yu, X.; Wilkinson, D.M.; Yang, J. Phytoplankton communities exhibit a stronger response to environmental changes than bacterioplankton in three subtropical reservoirs. Environ. Sci. Technol. 2015, 49, 10850–10858. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Sakamoto, S.; Yamaguchi, M. Nutrition and growth kinetics in nitrogen and phosphorus limited cultures of the novel red tide flagellate Chattonella ovata (Raphidophyceae). Harmful Algae 2008, 7, 26–32. [Google Scholar] [CrossRef]
- Gallardo Rodríguez, J.J.; Sánchez Mirón, A.; Cerón García, M.D.C.; Belarbi, E.H.; García Camacho, F.; Chisti, Y.; Molina Grima, E. Macronutrients requirements of the dinoflagellate Protoceratium reticulatum. Harmful Algae 2009, 8, 239–246. [Google Scholar] [CrossRef]
- Qiu, X.C.; Zhao, H.X.; Sun, X.X. Studies on relationship of phytoplankton and water environmental factors in Shahu Lake. J. Environ. Sci. China 2012, 33, 2265–2271. [Google Scholar]
- Shi, D.; Xu, Y.; Morel, F.M.M. Effects of the pH/pCO2 control method on medium chemistry and phytoplankton growth. Biogeosciences 2009, 6, 1199–1207. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Marzolf, E.R.; Goldman, C.R. Phosphorus and nitrogen limitation of phytoplankton growth in the freshwaters of north america: A review and critique of experimental enrichments. Can. J. Fish. Aquat. Sci. 1990, 47, 1468–1477. [Google Scholar] [CrossRef]
- Kim, L.H.; Choi, E.; Stenstrom, M.K. Sediment characteristics, phosphorus types and phosphorus release rates between river and lake sediments. Chemosphere 2003, 50, 53–61. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Urban Lakes | Provinces | Cities | Latitude | Longitude | Average Monthly Temperature (°C) | Surface Area(m2) | Urban Population | Built Year |
|---|---|---|---|---|---|---|---|---|
| TieXi (TX) | Inner Mongolia | Ordos | 39°49′16″ N | 109°58′07″ E | 15.3 | 1.7 × 104 | 2.0 × 106 | 2005 |
| XinHaiHu (XHH) | Ningxia | Shizuishan | 38°59′32″ N | 106°24′22″ E | 16.7 | 2.0 × 107 | 7.9 × 105 | 2004 |
| JinJi (JJ) | Ningxia | Wuzhong | 37°56′10″ N | 106°08′36″ E | 17.3 | 2.0 × 108 | 1.3 × 106 | Qin and Han Dynasties |
| ChangLe (CL) | Shaanxi | Xi’an | 34°16′04″ N | 109°00′00″ E | 24.7 | 2.2 × 105 | 8.8 × 106 | 1956 |
| XiangShan (XS) | Henan | Xinyang | 31°34′27″ N | 114°55′00″ E | 26.0 | 1.1 × 107 | 6.4 × 106 | 1969 |
| AiXi (AX) | Jiangxi | Nanchang | 28°42′56″ N | 115°59′21″ E | 29.7 | 4.5 × 106 | 5.5 × 107 | 2007 |
| HuiLongShan (HLS) | Jiangsu | Zhenjiang | 32°09′24″ N | 119°27′07″ E | 22.0 | 1.3 × 107 | 3.1 × 106 | 1977 |
| GaoTie (GT) | Jiangsu | Changzhou | 31°51′21″ N | 119°58′07″ E | 26.3 | 1.0 × 105 | 3.8 × 106 | 2017 |
| JinSha (JS) | Zhejiang | Hangzhou | 30°18′52″ N | 120°20′00″ E | 28.3 | 3.1 × 104 | 9.2 × 106 | 2018 |
| XiLiu (XL) | Henan | Zhengzhou | 34°46′00″ N | 113°34′36″ E | 28.7 | 4.6 × 108 | 1.1 × 107 | 2012 |
| ZiZhuYuan (ZZY) | Beijing | Beijing | 39°56′48″ N | 116°19′04″ E | 23.3 | 1.9 × 105 | 2.2 × 107 | 1953 |
| GuiLong (GL) | Yunnan | Kunming | 25°04′10″ N | 102°42′53″ E | 24.0 | 1.6 × 105 | 6.7 × 106 | 2006 |
| ZhuZhai (ZZ) | Shanghai | Shanghai | 31°12′53″ N | 121°17′36″ E | 27.3 | 3.5 × 104 | 1.4 × 107 | 2004 |
| ZhongShan (ZS) | Shenzhen | Shenzhen | 31°13′27″ N | 121°25′23″ E | 33.3 | 3.5 × 104 | 1.1 × 107 | 2004 |
| West lake (WL) | Zhejiang | Hangzhou | 30°13′14″ N | 120°06′30″ E | 29.0 | 6.4 × 106 | 9.2 × 106 | Qin and Han Dynasties |
| Yunv (YN) | Sichuan | Mianyang | 31°29′54″ N | 104°44′14″ E | 25.3 | 2.4 × 105 | 5.4 × 106 | 1986 |
| Urban Lakes | pH | TN | NO3--N | NO2--N | NH4+-N | TP | COD Mn | Fe | Mn | TOC |
|---|---|---|---|---|---|---|---|---|---|---|
| (mg/L) | ||||||||||
| TieXi (TX) | 9.30 ± 0.24a | 0.44 ± 0.06l | 0.07 ± 0.01ef | 0.01 ± 0.00b | 0.03 ± 0.00j | 0.01 ± 0.00h | 5.79 ± 0.87c | 0.03 ± 0.00bcd | 0.01 ± 0.00bc | 5.74 ± 0.28bc |
| XinHaiHu (XHH) | 8.48 ± 0.06b | 3.84 ± 0.33a | 0.39 ± 0.09a | 0.04 ± 0.03a | 0.30 ± 0.05c | 0.21 ± 0.01a | 9.01 ± 0.31a | 0.02 ± 0.00de | 0.01 ± 0.00a | 9.77 ± 0.71a |
| JinJi (JJ) | 8.04 ± 0.14de | 0.85 ± 0.26gh | 0.06 ± 0.01fg | 0.01 ± 0.00b | 0.15 ± 0.07ef | 0.05 ± 0.00cd | 5.08 ± 0.10de | 0.04 ± 0.01bc | 0.01 ± 0.00a | 5.69 ± 0.56d |
| ChangLe (CL) | 7.72 ± 0.43de | 0.57 ± 0.11jk | 0.04 ± 0.01fg | 0.04 ± 0.01a | 0.10 ± 0.00ghi | 0.06 ± 0.00b | 4.70 ± 1.44f | 0.04 ± 0.01a | 0.00 ± 0.00c | 1.09 ± 0.20f |
| XiangShan (XS) | 7.45 ± 0.04gh | 1.02 ± 0.07g | 0.16 ± 0.03c | 0.01 ± 0.00b | 0.01 ± 0.00j | 0.02 ± 0.00g | 3.60 ± 0.25f | 0.02 ± 0.00ef | 0.01 ± 0.00ab | 4.07 ± 0.55d |
| AiXi (AX) | 7.40 ± 0.09gh | 0.57 ± 0.16ij | 0.12 ± 0.03d | 0.01 ± 0.00b | 0.13 ± 0.07e | 0.03 ± 0.01f | 5.13 ± 0.86cd | 0.04 ± 0.012ab | 0.01 ± 0.002a | 5.61 ± 0.17bc |
| HuiLongShan (HLS) | 7.88 ± 0.10def | 0.52 ± 0.02f | 0.06 ± 0.01fg | 0.01 ± 0.00b | 0.07 ± 0.01f | 0.03 ± 0.01cd | 4.43 ± 0.18ef | 0.02 ± 0.00de | 0.01 ± 0.00a | 4.66 ± 0.59b |
| GaoTie (GT) | 8.20 ± 0.19bc | 0.35 ± 0.11kl | 0.04 ± 0.02fg | 0.01 ± 0.00b | 0.07 ± 0.01i | 0.05 ± 0.01ef | 4.21 ± 0.11f | 0.02 ± 0.00ef | 0.01 ± 0.00a | 3.78 ± 0.54a |
| JinSha (JS) | 8.15 ± 0.19cd | 2.08 ± 0.22m | 0.04 ± 0.00fg | 0.01 ± 0.00b | 0.15 ± 0.03hi | 0.05 ± 0.01cd | 4.35 ± 0.80f | 0.02 ± 0.01ef | 0.01 ± 0.00a | 9.33 ± 0.45a |
| XiLiu (XL) | 7.79 ± 0.02ef | 0.77 ± 0.10b | 0.37 ± 0.06g | 0.04 ± 0.01a | 0.41 ± 0.01f | 0.05 ± 0.01cd | 4.42 ± 0.30ef | 0.02 ± 0.00de | 0.01 ± 0.00a | 3.34 ± 0.19b |
| ZiZhuYuan (ZZY) | 7.95 ± 0.02de | 0.32 ± 0.03hi | 0.22 ± 0.03a | 0.01 ± 0.00b | 0.11 ± 0.01b | 0.05 ± 0.01cd | 5.67 ± 0.80c | 0.01 ± 0.01ef | 0.01 ± 0.00a | 3.17 ± 0.62e |
| GuiLong (GL) | 7.27 ± 0.27hi | 1.23 ± 0.40m | 0.19 ± 0.23b | 0.01 ± 0.00b | 0.16 ± 0.03gh | 0.05 ± 0.00d | 6.23 ± 1.12cd | 0.02 ± 0.01ced | 0.01 ± 0.00a | 5.77 ± 0.26b |
| ZhuZhai (ZZ) | 7.28 ± 0.30i | 0.70 ± 0.06ij | 0.13 ± 0.05de | 0.01 ± 0.00b | 0.01 ± 0.00j | 0.03 ± 0.01ef | 8.35 ± 0.10a | 0.01 ± 0.00efg | 0.01 ± 0.002a | 4.76 ± 0.14d |
| ZhongShan (ZS) | 7.45 ± 0.03gh | 1.37 ± 0.04e | 0.05 ± 0.02fg | 0.01 ± 0.00b | 0.25 ± 0.00d | 0.05 ± 0.00cd | 7.1 ± 0.27b | 0.02 ± 0.00ef | 0.01 ± 0.002a | 4.31 ± 0.41e |
| West Lake (WL) | 7.94 ± 0.04de | 1.05 ± 0.47d | 0.15 ± 0.03cd | 0.01 ± 0.00b | 0.11 ± 0.01g | 0.04 ± 0.00e | 4.41 ± 0.81f | 0.00 ± 0.00fg | 0.01 ± 0.000a | 2.82 ± 0.49e |
| YuNv (YN) | 7.61 ± 0.05fg | 2.08 ± 0.10c | 0.11 ± 0.01de | 0.10 ± 0.00b | 0.50 ± 0.04a | 0.06 ± 0.01b | 8.33 ± 0.08a | 0.00 ± 0.00g | 0.01 ± 0.002a | 5.59 ± 0.86b |
| One-way ANOVA | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Parameters | Number |
|---|---|
| Avg. weighted degree | 0.548 |
| Network diameter | 7 |
| Graph density | 0.057 |
| Modularity | 1.446 |
| Connected components | 6 |
| Avg. clustering coefficient | 0.209 |
| Avg. path length | 3.231 |
| Nodes | 62 |
| Edges | 108 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; He, H.; Zong, R.; Liu, K.; Miao, Y.; Yan, M.; Xu, L. Geographical Patterns of Algal Communities Associated with Different Urban Lakes in China. Int. J. Environ. Res. Public Health 2020, 17, 1009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17031009
Chen S, He H, Zong R, Liu K, Miao Y, Yan M, Xu L. Geographical Patterns of Algal Communities Associated with Different Urban Lakes in China. International Journal of Environmental Research and Public Health. 2020; 17(3):1009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17031009
Chicago/Turabian StyleChen, Shengnan, Huiyan He, Rongrong Zong, Kaiwen Liu, Yutian Miao, Miaomiao Yan, and Lei Xu. 2020. "Geographical Patterns of Algal Communities Associated with Different Urban Lakes in China" International Journal of Environmental Research and Public Health 17, no. 3: 1009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17031009