Effects of Sulfamethoxazole and 2-Ethylhexyl-4-Methoxycinnamate on the Dissimilatory Nitrate Reduction Processes and N2O Release in Sediments in the Yarlung Zangbo River


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample Collection and Pretreatment
2.3. Slurry Incubation Experiments for the Removal of SMX and EHMC
2.4. Slurry Incubation Experiments for the Nitrate Reduction Processes
2.5. Slurry Incubation Experiments of N2O Release
2.6. Gene Expression Assay
2.7. Statistical Analysis
3. Results and Discussion
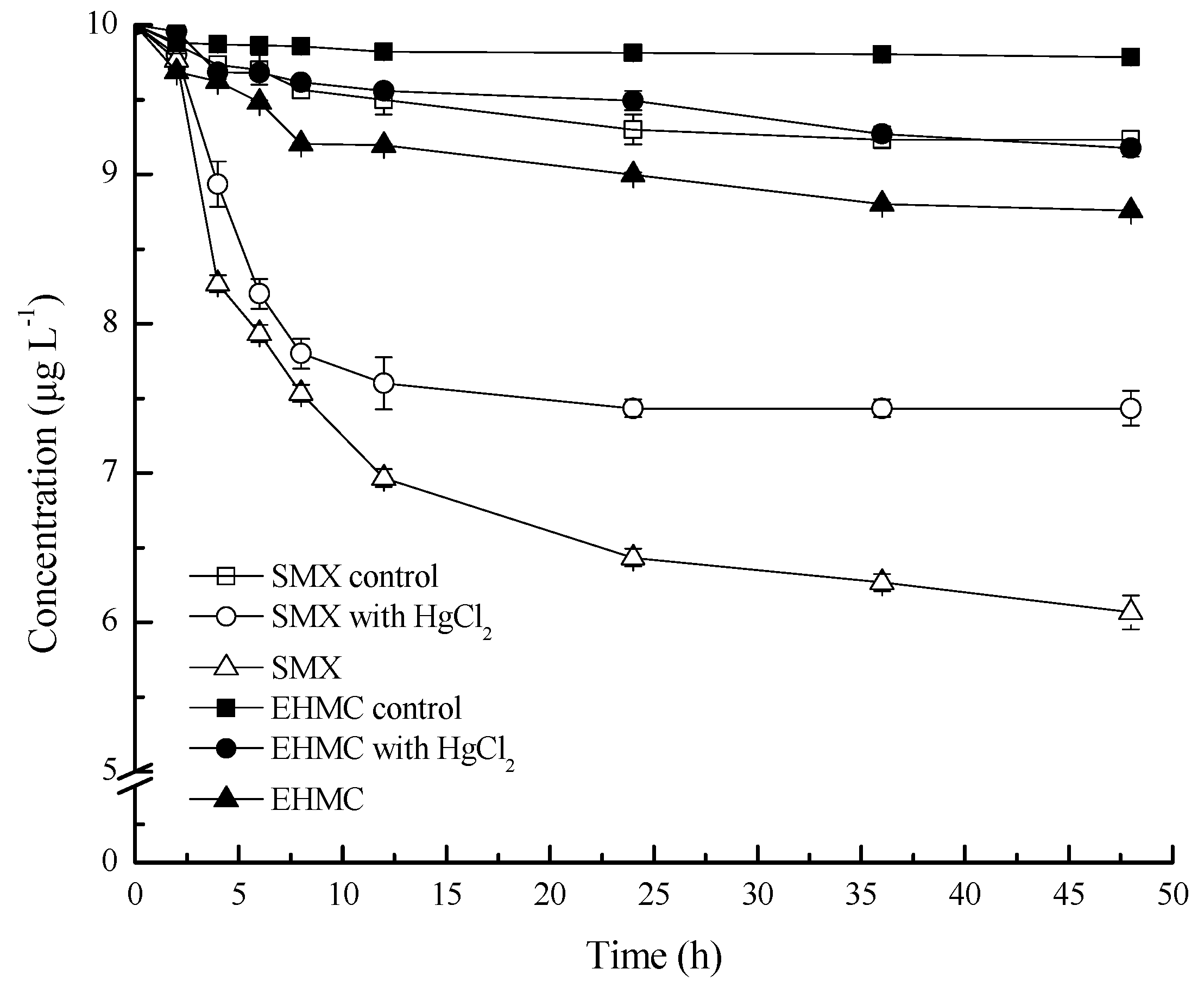
3.1. Adsorption and Degradation of SMZ and EHMC in the Slurries
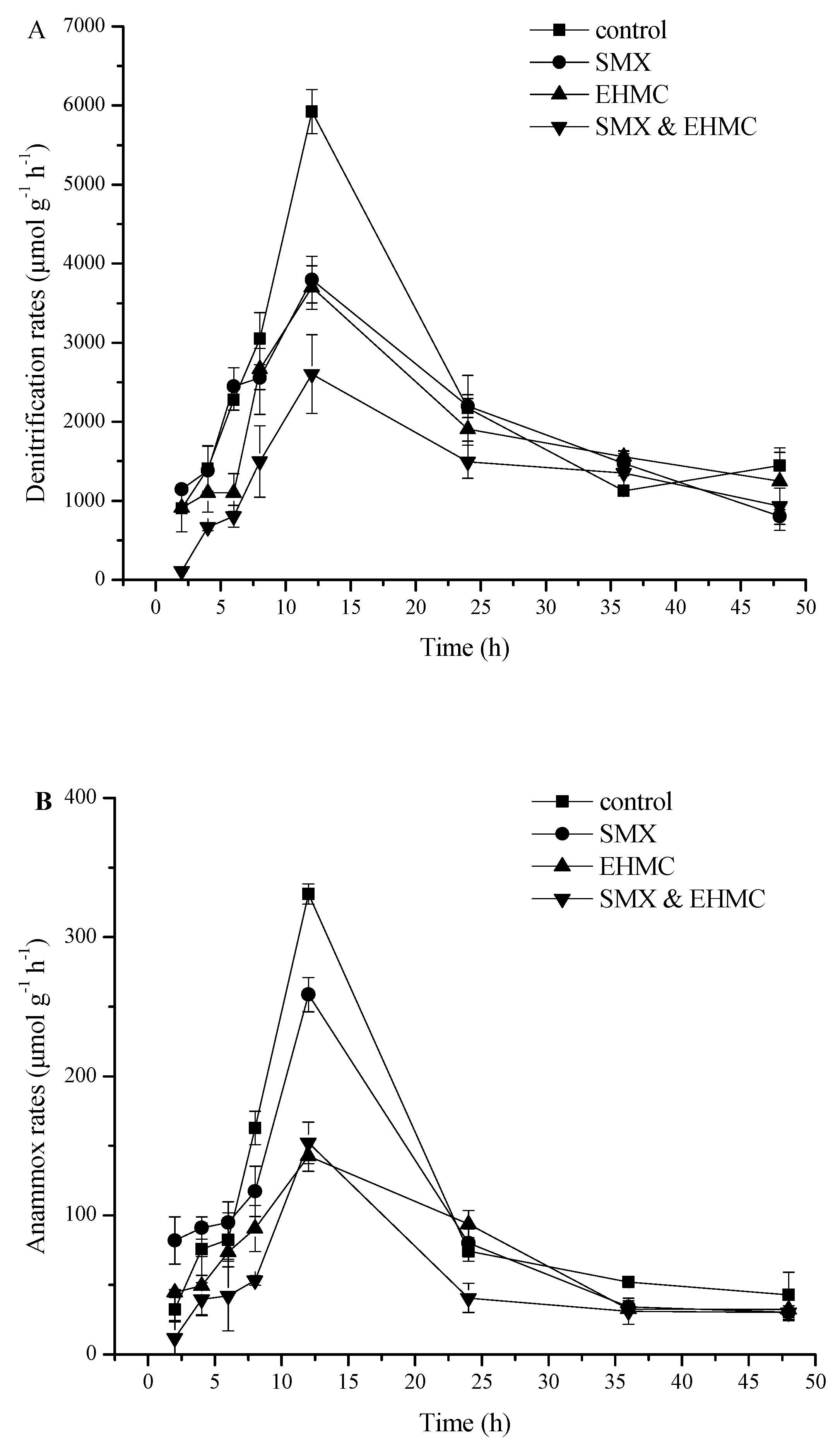
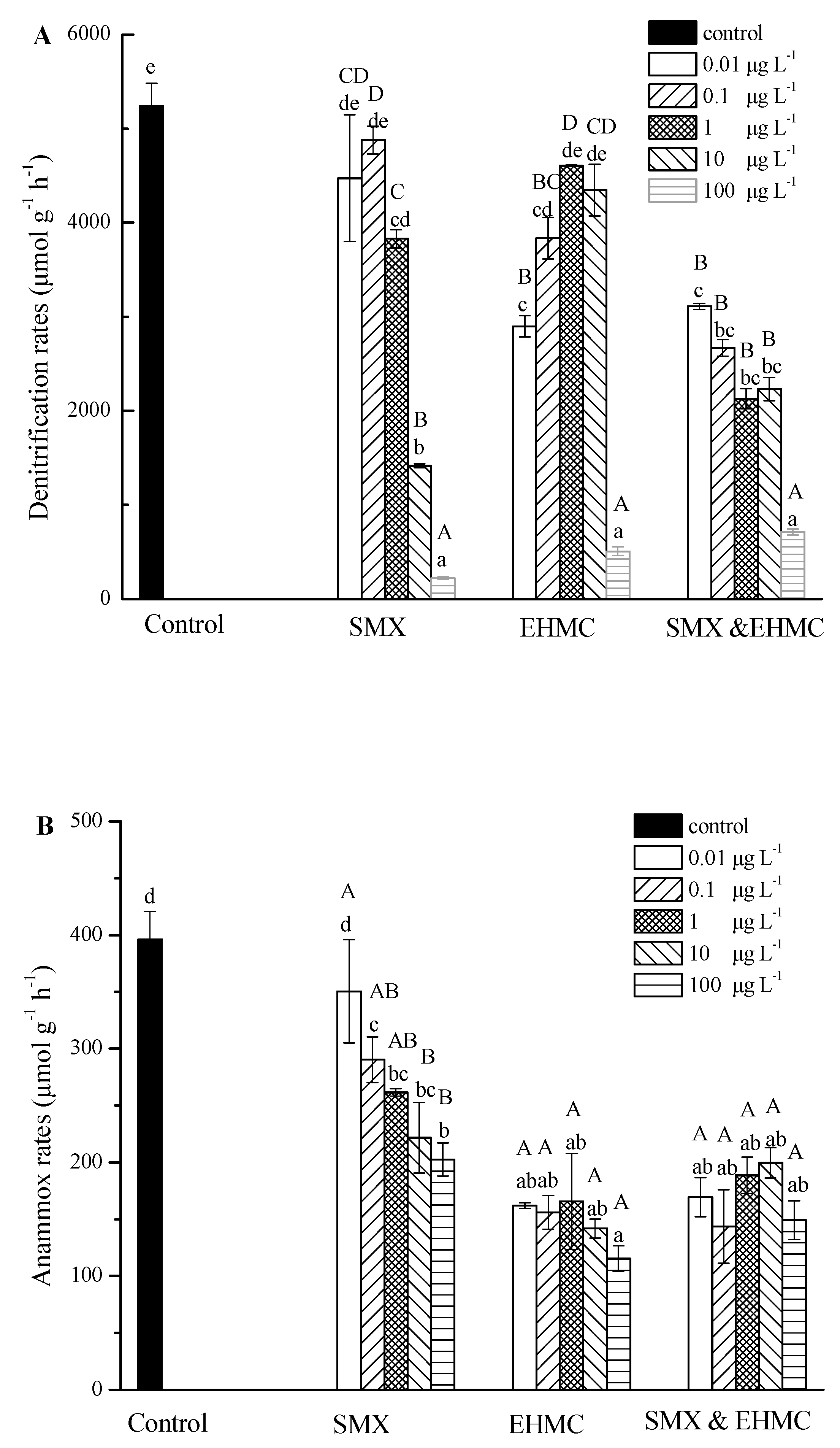
3.2. Influence of SMX and EHMC on the Nitrate Reduction Processes
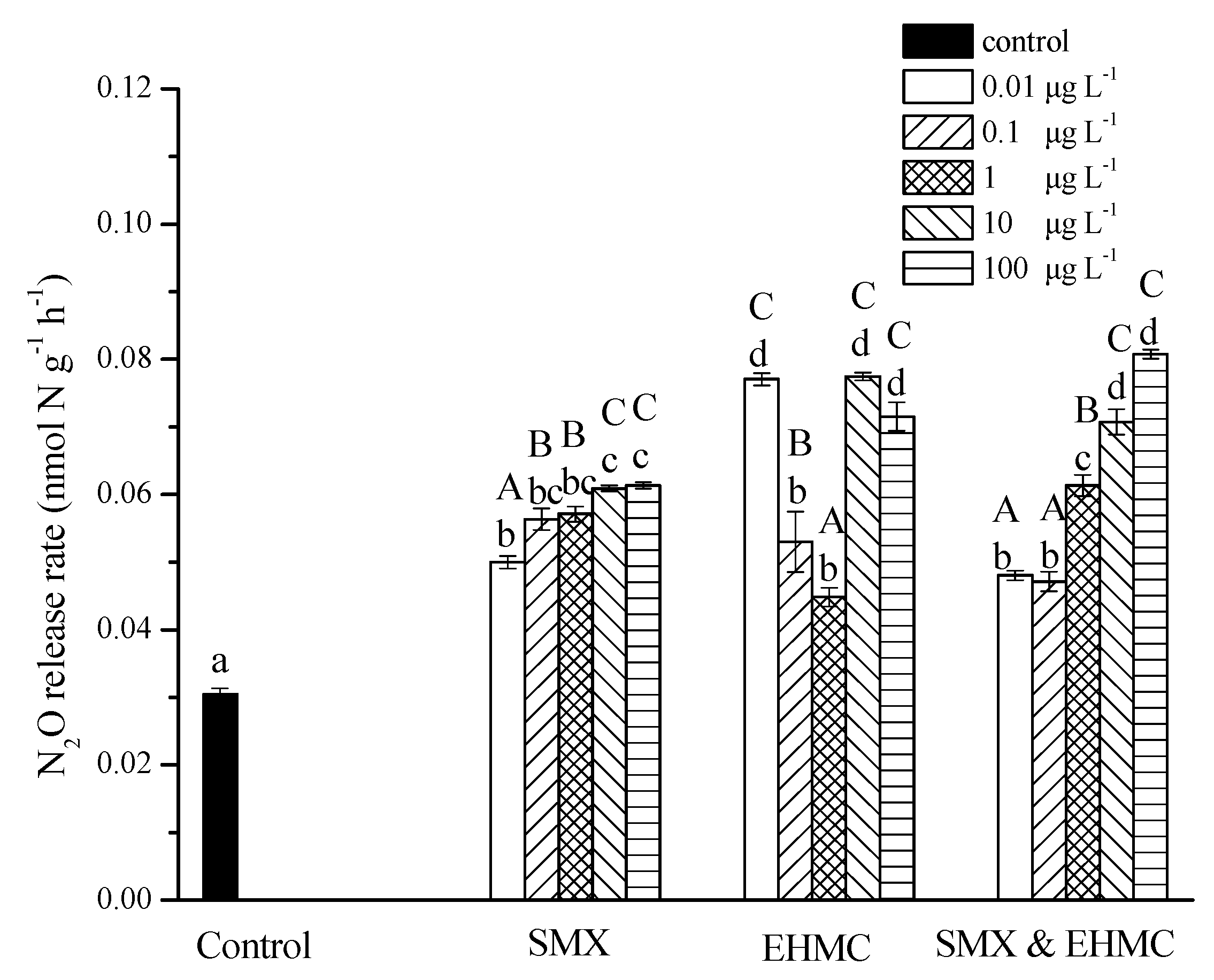
3.3. Influence of SMX and EHMC on N2O Release
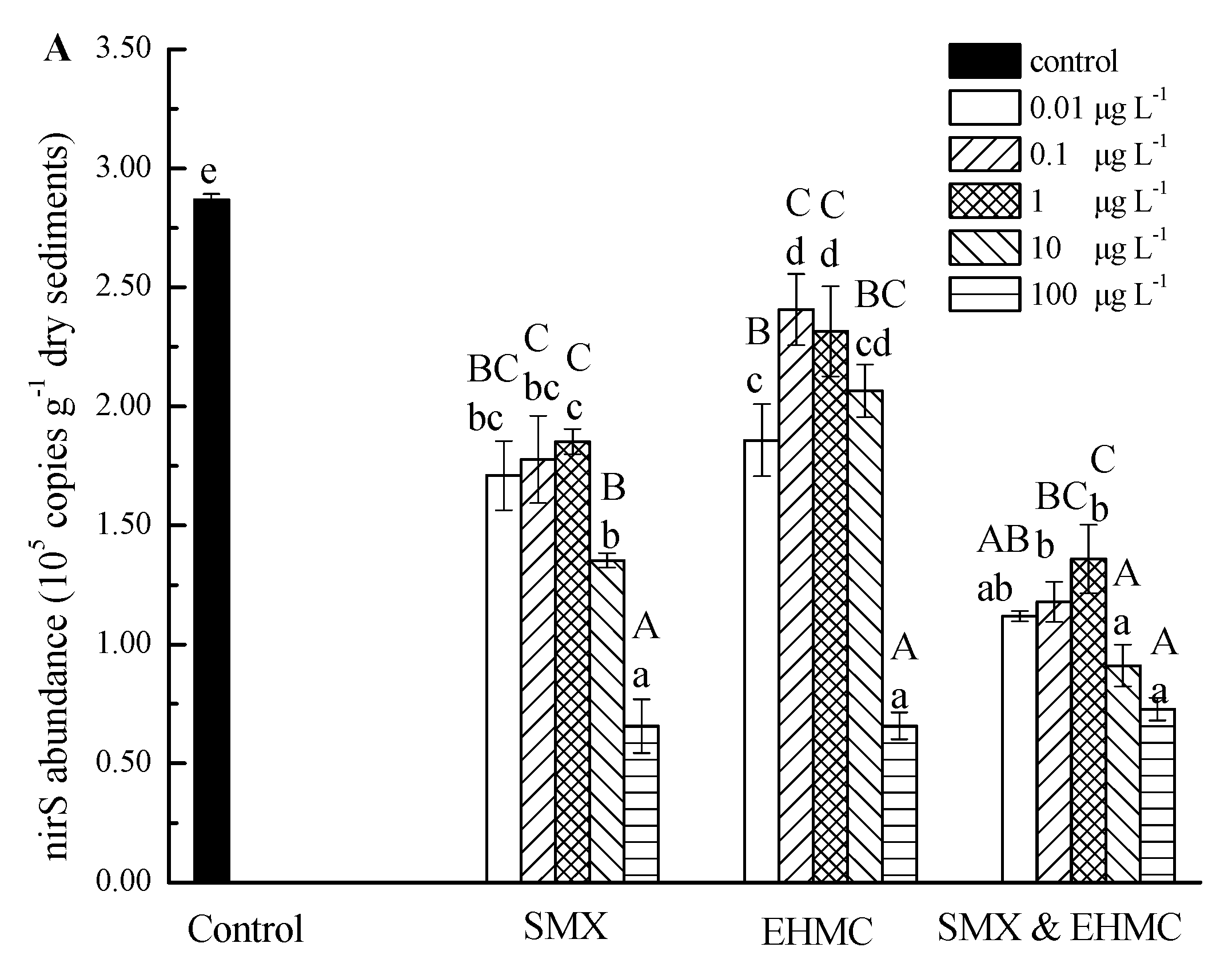
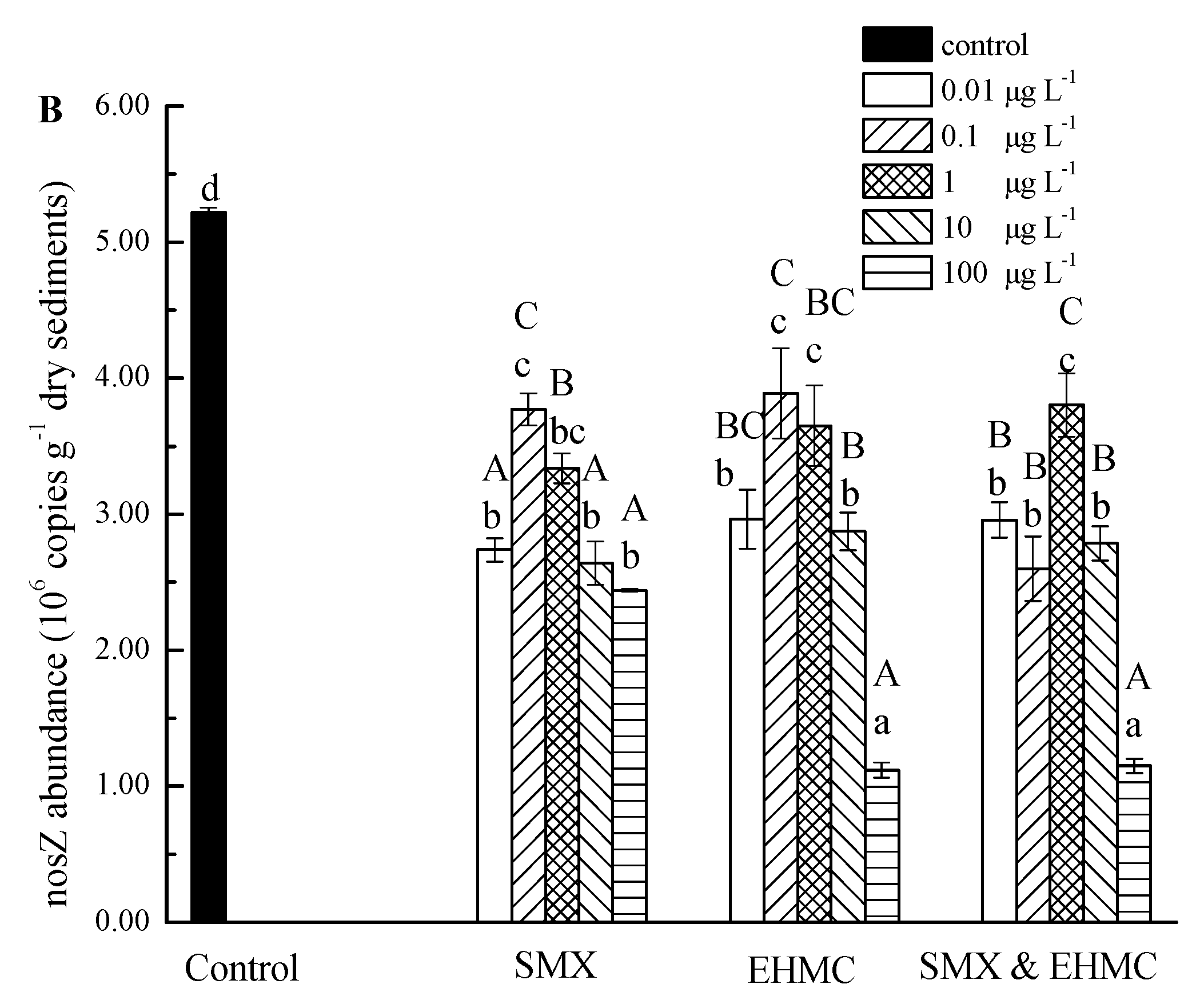
3.4. The Influence of SMX and EHMC on the Abundance of NirS and NosZ
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Canfield, D.E.; Glazer, A.N.; Falkowski, P.G. The evolution and future of earth’s nitrogen cycle. Science 2010, 330, 192–196. [Google Scholar] [CrossRef] [Green Version]
- Lapointe, B.E.; Brewton, R.A.; Herren, L.W.; Porter, J.W.; Hu, C. Nitrogen enrichment, altered stoichiometry, and coral reef decline at Looe Key, Florida Keys, USA: A 3-decade study. Mar. Biol. 2019, 8, 166. [Google Scholar] [CrossRef]
- Zhang, W.; Jin, X.; Liu, D.; Lang, C.; Shan, B. Temporal and spatial variation of nitrogen and phosphorus and eutrophication assessment for a typical arid river—Fuyang River in northern China. J. Environ. Sci. 2017, 55, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yao, L.; Jiang, X.; Guo, L.; Liu, G. Sediment denitrification in Yangtze lakes is mainly influenced by environmental conditions but not biological communities. Sci. Total Environ. 2018, 616, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Salahudeen, J.H.; Reshmi, R.R.; Krishnan, K.A.; Ragi, M.S.; Vincent, S.G.T. Denitrification rates in estuarine sediments of Ashtamudi, Kerala, India. Environ. Monit. Assess. 2018, 190, 323. [Google Scholar] [CrossRef] [PubMed]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, M.J.; Ren, H.; Li, J.; Musenze, R.S. Seasonal variations of nitrous oxide fluxes and soil denitrification rates in subtropical freshwater and brackish tidal marshes of the min river estuary. Sci. Total Environ. 2018, 616–617, 1404–1413. [Google Scholar] [CrossRef]
- Murray, R.H.; Erler, D.V.; Eyre, B.D. Nitrous oxide fluxes in estuarine environments: Response to global change. Global Change Biol. 2015, 21, 3219–3245. [Google Scholar] [CrossRef]
- Descloux, S.; Chanudet, V.; Serça, D.; Guérin, F. Methane and nitrous oxide annual emissions from an old eutrophic temperate reservoir. Sci. Total Environ. 2017, 598, 959–972. [Google Scholar] [CrossRef]
- Peng, X.; Fawcett, S.E.; Oostende, N.V.; Wolf, M.J.; Ward, B.B. Nitrogen uptake and nitrification in the subarctic North Atlantic Ocean: N uptake and nitrification in subarctic Atlantic. Limnol. Oceanogr. 2018, 63, 1462–1487. [Google Scholar] [CrossRef]
- Jahangir, M.M.R.; Fenton, O.; Mueller, C.; Harrington, R.; Johnston, P.; Richards, K.G. In situ denitrification and DNRA rates in groundwater beneath an integrated constructed wetland. Water Res. 2017, 111, 254–264. [Google Scholar] [CrossRef]
- Yao, X.; Zhang, L.; Zhang, Y.; Xu, H.; Jiang, H. Denitrification occurring on suspended sediment in a large, shallow, subtropical lake (Poyang Lake, China). Environ. Pollut. 2016, 219, 501–511. [Google Scholar] [CrossRef]
- Wang, Y.J.H.; Wang, R.; Guo, S. Responses of nitrification and denitrification to nitrogen and phosphorus fertilization: Does the intrinsic soil fertility matter? Plant Soil 2019, 440, 443–456. [Google Scholar] [CrossRef]
- Li, F.; Xu, Z.; Liu, W.; Zhang, Y. The impact of climate change on runoff in the Yarlung Tsangpo River basin in the Tibetan Plateau. Stoch. Env. Res. Risk Assess. 2014, 28, 517–526. [Google Scholar] [CrossRef]
- Wang, P.; Wang, X.; Wang, C.; Miao, L.; Hou, J.; Yuan, Q. Shift in bacterioplankton diversity and structure: Influence of anthropogenic disturbances along the Yarlung Tsangpo River on the Tibetan Plateau, China. Sci. Rep. 2017, 7, 12529. [Google Scholar] [CrossRef] [PubMed]
- Immerzeel, W.W.; van Beek, L.P.H.; Bierkens, M.F.P. Climate change will affect the Asian water towers. Science 2010, 328, 1382–1385. [Google Scholar] [CrossRef]
- Richmond, E.K.; Grace, M.R.; Kelly, J.J.; Reisinger, A.J.; Walters, D.M. Pharmaceuticals and personal care products (PPCPs) are ecological disrupting compounds (EcoDC). Elementa. Sci. Anthrop. 2017, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Ram, B.; Honda, R.; Poopipattana, C.; Canh, V.D.; Chaminda, T.; Furumai, H. Concurrence of antibiotic resistant bacteria (ARB), viruses, pharmaceuticals and personal care products (PPCPs) in ambient waters of Guwahati, India: Urban vulnerability and resilience perspective. Sci. Total Environ. 2019, 693, 133640. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Yin, G.; Liu, M.; Zhou, J.; Zheng, Y.; Gao, J.; Zong, H.; Yang, Y.; Gao, L.; Tong, C. Effects of sulfamethazine on denitrification and the associated N2O release in estuarine and coastal sediments. Environ. Sci. Technol. 2015, 49, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Hou, L.; Liu, M.; Zheng, Y.; Li, X.; Lin, X.; Gao, J.; Jiang, X. Effects of thiamphenicol on nitrate reduction and N2O release in estuarine and coastal sediments. Environ. Pollut. 2016, 214, 265–272. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, T.; Cheng, P.; AO, J.J.; Wang, W.; Zhang, H. Organic UV filters inhibit multixenobiotic resistance (MXR) activity in Tetrahymena Thermophila: Investigations by the Rhodamine 123 accumulation assay and molecular docking. Ecotoxicology 2016, 25, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, P.L.; Yang, Q.X. Occurrence and diversity of antibiotic resistance inuntreated hospital wastewater. Sci. Total Environ. 2018, 621, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Wu, Y.; Zhang, W.; Zhong, J.; Lou, Q.; Yang, P.; Fang, Y. Occurrence, distribution, and risk assessment of antibiotics in the surface water of Poyang Lake, the largest freshwater lake in China. Chemosphere 2017, 184, 137–147. [Google Scholar] [CrossRef]
- Kafaei, R.; Papari, F.; Seyedabadi, M.; Sahebi, S.; Tahmasebi, R.; Ahmadi, M.; Sorial, G.A.; Asgari, G.; Ramavandi, B. Occurrence, distribution, and potential sources of antibiotics pollution in the water-sediment of the northern coastline of the Persian Gulf, Iran. Sci. Total Environ. 2018, 627, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Huwe, B. Effect of pH and soil structure on transport of sulphonamide antibiotics in agricultural soils. Environ. Pollut. 2016, 213, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Ekpeghere, K.I.; Kim, U.J.; Sung-Hee, O.; Kim, H.Y.; Oh, J.E. Distribution and seasonal occurrence of UV filters in rivers and wastewater treatment plants in Korea. Sci Total Environ. 2016, 542, 121–128. [Google Scholar] [CrossRef]
- Peng, X.; Xiong, S.; Ou, W.; Wang, Z.; Tan, J.; Jin, J.; Tang, C.; Liu, J.; Fan, Y. Persistence, temporal and spatial profiles of ultraviolet absorbents and phenolic personal care products in riverine and estuarine sediment of the Pearl River catchment, China. J. Hazard. Mater. 2016, 323, 139–146. [Google Scholar] [CrossRef]
- Gackowska, A.; Studzinski, W.; Kudlek, E.; Dudziak, M.; Gaca, J. Estimation of physicochemical properties of 2-ethylhexyl-4-methoxycinnamate (EHMC) degradation products and their toxicological evaluation. Environ. Sci. Pollut. Res. 2018, 25, 16037–16049. [Google Scholar] [CrossRef] [Green Version]
- Brandt, K.K.; Amezquita, A.; Backhaus, T.; Boxall, A.; Coors, A.; Heberer, T.; Lawrence, J.R.; Lazorchak, J.; Schonfeld, J.; Snape, J.R.; et al. Ecotoxicological assessment of antibiotics: A call for improved consideration of microorganisms. Environ. Int. 2015, 85, 189–205. [Google Scholar] [CrossRef]
- Shan, J.; Yang, P.; Rahman, M.M.; Shang, X.; Yan, X. Tetracycline and sulfamethazine alter dissimilatory nitrate reduction processes and increase N2O release in rice fields. Environ. Pollut. 2018, 242, 788–796. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Zhong, Z.; Guo, C.; Li, L.; He, Y.; Fan, W.; Chen, Y. Degradation of sulfonamides antibiotics in lake water and sediment. Environ. Sci. Pollut. Res. 2013, 20, 2372–2380. [Google Scholar] [CrossRef] [PubMed]
- McTigue, N.D.; Gardner, W.S.; Dunton, K.H.; Hardison, A.K. Biotic and abiotic controls on co-occurring nitrogen cycling processes in shallow Arctic shelf sediments. Nat. Commun. 2016, 7, 13145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Lu, G.; Yan, Z.; Liu, J.; Dong, H.; Jiang, R.; Zhou, R.; Zhang, P.; Sun, Y.; Matthew, N. Occurrence, spatial-temporal distribution and ecological risks of pharmaceuticals and personal care products response to water diversion across the rivers in Nanjing, China. Environ. Pollut. 2019, 255, 113132. [Google Scholar] [CrossRef] [PubMed]
- Deng, F.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Li, X.; Lin, X.; Chen, F.; Gao, J.; Jiang, X. Dissimilatory nitrate reduction processes and associated contribution to nitrogen removal in sediments of the Yangtze Estuary. J. Geophys. Res.-Biogeo. 2015, 120, 1521–1531. [Google Scholar] [CrossRef]
- Long, X.E.; Shen, J.P.; Wang, J.T.; Zhang, L.M.; He, J.Z. Contrasting response of two grassland soils to N addition and moisture levels: N2O emission and functional gene abundance. J. Soil Sediment. 2016, 17, 1–9. [Google Scholar] [CrossRef]
- Thompson, K.A.; Deen, B.; Dunfield, K.E. Soil denitrifier community size changes with land use change to perennial bioenergy cropping systems. Soil 2016, 2, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Foshtomi, M.Y.; Leliaert, F.; Derycke, S.; Willems, A.; Vanaverbeke, J. The effect of bio-irrigation by the polychaete lanice conchilega on active denitrifiers: Distribution, diversity and composition of nosZ gene. PLoS ONE 2018, 13, e0192391. [Google Scholar]
- Srinivasan, P.; Sarmah, A.K.; Manley-Harris, M. Co-contaminants and factors affecting the sorption behaviour of two sulfonamides in pasture soils. Environ. Pollut. 2013, 180, 165–172. [Google Scholar] [CrossRef]
- Ahmed, M.B.; Zhou, J.; Ngo, H.H.; Guo, W.; Johir, M.A.H.; Sornalingam, K. Single and competitive sorption properties and mechanism of functionalized biochar for removing sulfonamide antibiotics from water. Chem. Eng. J. 2017, 311, 348–358. [Google Scholar] [CrossRef]
- Shao, S.C.; Hu, Y.Y.; Cheng, J.H.; Chen, Y.C. Effects of carbon source, nitrogen source, and natural algal powder-derived carbon source on biodegradation of tetracycline (TEC). Bioresour. Technol. 2019, 288, 12156. [Google Scholar] [CrossRef]
- Ding, C.; He, J. Effect of antibiotics in the environment on microbial populations. Appl. Microbiol. Biotechnol. 2010, 87, 925–941. [Google Scholar] [CrossRef] [PubMed]
- Karkman, A.; Parnanen, K.; Larsson, D.G.J. Fecal pollution can explain antibiotic resistance gene abundances in anthropogenically impacted environments. Nat. Commun. 2019, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. Fems. Microbiol. Rev. 2018, 42, 68–80. [Google Scholar] [CrossRef]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Buergmann, H.; Sorum, H.; Norstrom, M.; Pons, M.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Yang, H.; Dong, H.; Ma, B.; Sun, H.; Pan, T.; Jiang, R.; Zhou, R.; Shen, J.; Liu, J.; et al. Occurrence and ecological risk assessment of organic micropollutants in the lower reaches of the Yangtze River, China: A case study of water diversion. Environ. Pollut. 2018, 239, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Yao, L.; Guo, L.; Liu, G.; Liu, W. Multi-scale factors affecting composition, diversity, and abundance of sediment denitrifying microorganisms in Yangtze lakes. Appl. Microbiol. Biotechnol. 2017, 101, 8015–8027. [Google Scholar] [CrossRef] [PubMed]
- DeVries, S.L.; Loving, M.; Li, X.; Zhang, P. The effect of ultralow-dose antibiotics exposure on soil nitrate and N2O flux. Sci. Rep. 2015, 5, 16818. [Google Scholar] [CrossRef] [PubMed]
- Conkle, J.L.; White, J.R. An initial screening of antibiotic effects on microbial respiration in wetland soils. J. Environ. Sci. Health A 2012, 47, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Hou, L.; Liu, M.; Zheng, Y.; Li, X.; Lin, X.; Gao, J.; Jiang, X.; Wang, R.; Yu, C. Effects of multiple antibiotics exposure on denitrification process in the Yangtze Estuary sediments. Chemosphere 2017, 171, 118–125. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter in Water | Value | Parameter in Sediment | Value |
|---|---|---|---|
| Temperature (°C) | 13.7 | TN (mg kg−1) | 354.87 |
| NH4+ (mg L−1) | 0.34 | NH4+ (mg kg−1) | 4.69 |
| NO3− (mg L−1) | 0.45 | NO3− (mg kg−1) | 2.37 |
| NO2− (mg L−1) | 0.07 | NO2− (mg kg−1) | 0.21 |
| SMX (ng L−1) | 2.3 | SMX (ng g−1) | 4.36 |
| EHMC (ng L−1) | 0.9 | EHMC (ng g−1) | 16.4 |
| Target | Primer | Nucleotide Sequence | Thermal Profile | Reference |
|---|---|---|---|---|
| nirS | cd3aF | GTSAACGTSAAGGARACSGG | 95 °C, 3 min, 1cycle, 95 °C for 30 s, 56 °C for 30 s, 72 °C for 30 s, 40 cycles | [36] |
| R3cd | GASTTCGGRTGSGTCTTGA | |||
| nosZ | nosZ2F | CGCRACGGCAASAAGGTSMSSGT | 95 °C, 3 min, 1 cycle, 95 °C for 30 s, 56 °C for 30 s, 70 °C for 30 s, 40 cycles | [37] |
| nosZ2R | CAKRTGCAKSGCRTGGCAGAA |
| Varibles | Denitrification | Anammox | nirS | nosZ | N2O Release |
|---|---|---|---|---|---|
| Denitrification | 1 | 0.403 ** | 0.684 ** | 0.622 ** | −0.409 ** |
| Anammox | 1 | 0.437 ** | 0.570 ** | −0.519 ** | |
| nirS | 1 | 0.789 ** | −0.514 ** | ||
| nosZ | 1 | −0.657 ** | |||
| N2O release | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Lu, G.; Xue, C. Effects of Sulfamethoxazole and 2-Ethylhexyl-4-Methoxycinnamate on the Dissimilatory Nitrate Reduction Processes and N2O Release in Sediments in the Yarlung Zangbo River. Int. J. Environ. Res. Public Health 2020, 17, 1822. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17061822
Xu H, Lu G, Xue C. Effects of Sulfamethoxazole and 2-Ethylhexyl-4-Methoxycinnamate on the Dissimilatory Nitrate Reduction Processes and N2O Release in Sediments in the Yarlung Zangbo River. International Journal of Environmental Research and Public Health. 2020; 17(6):1822. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17061822
Chicago/Turabian StyleXu, Huiping, Guanghua Lu, and Chenwang Xue. 2020. "Effects of Sulfamethoxazole and 2-Ethylhexyl-4-Methoxycinnamate on the Dissimilatory Nitrate Reduction Processes and N2O Release in Sediments in the Yarlung Zangbo River" International Journal of Environmental Research and Public Health 17, no. 6: 1822. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17061822