System-Specific Complex Interactions Shape Soil Organic Carbon Distribution in Coastal Salt Marshes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Analyses
2.3. Statistical Analyses
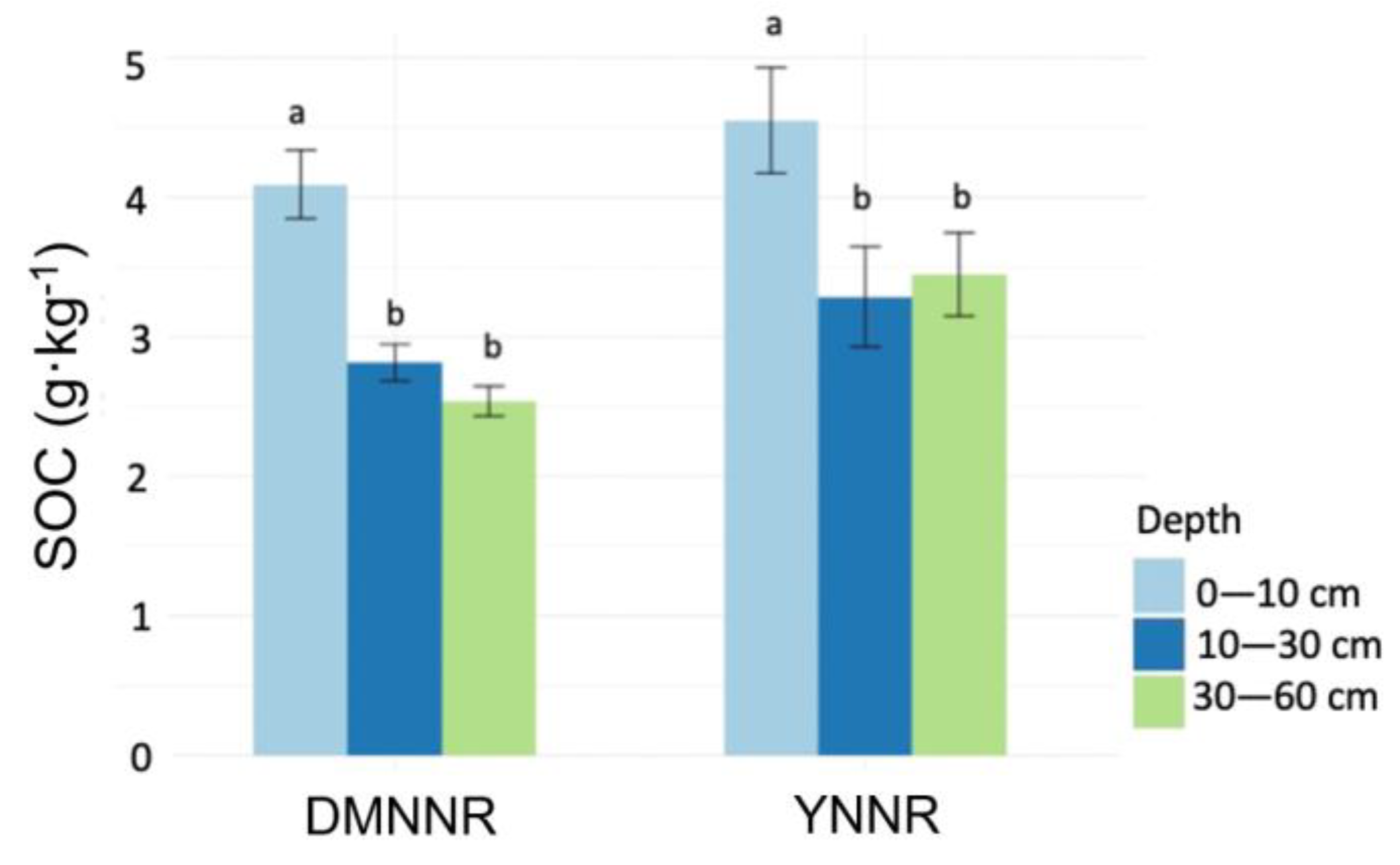
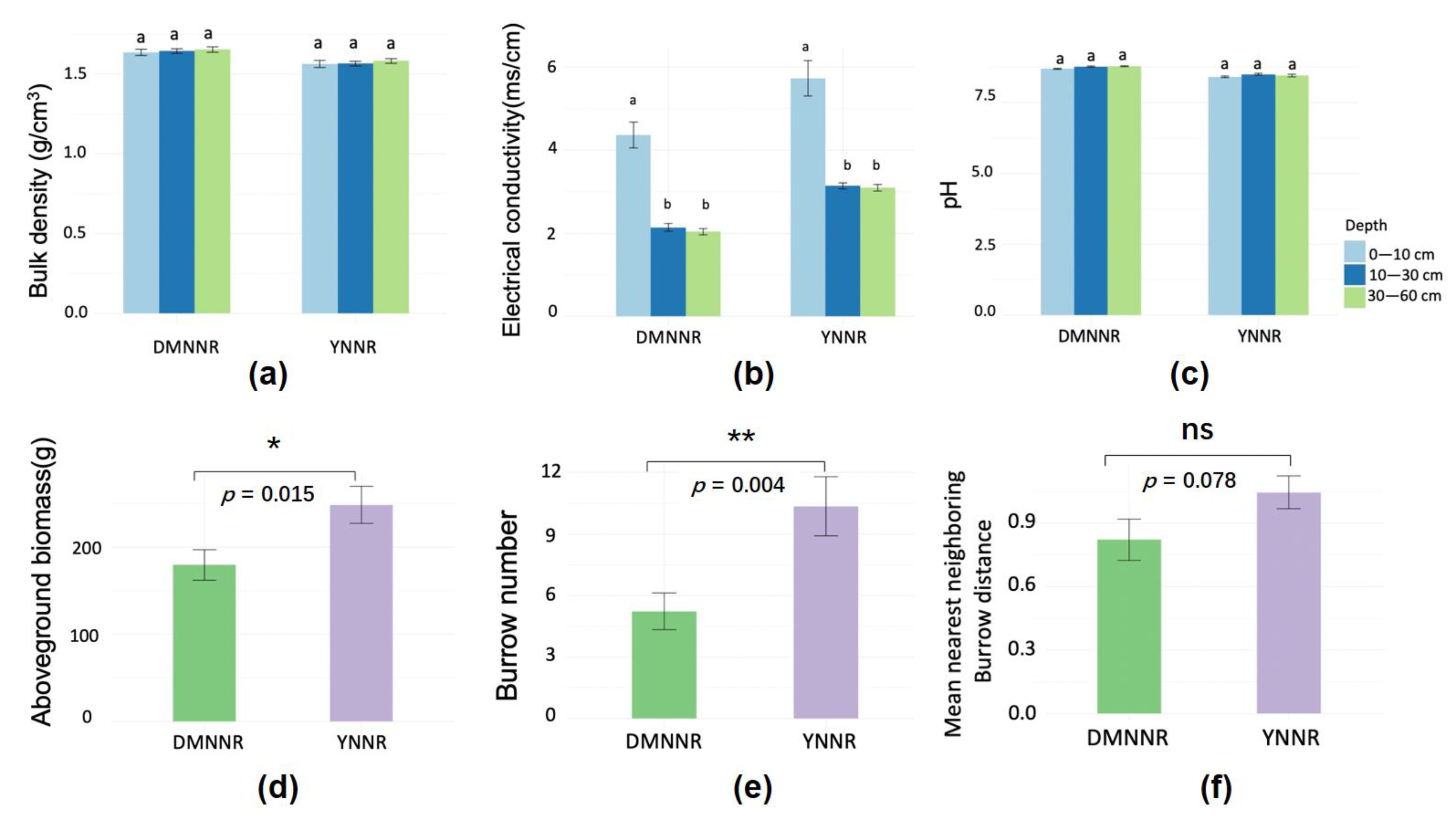
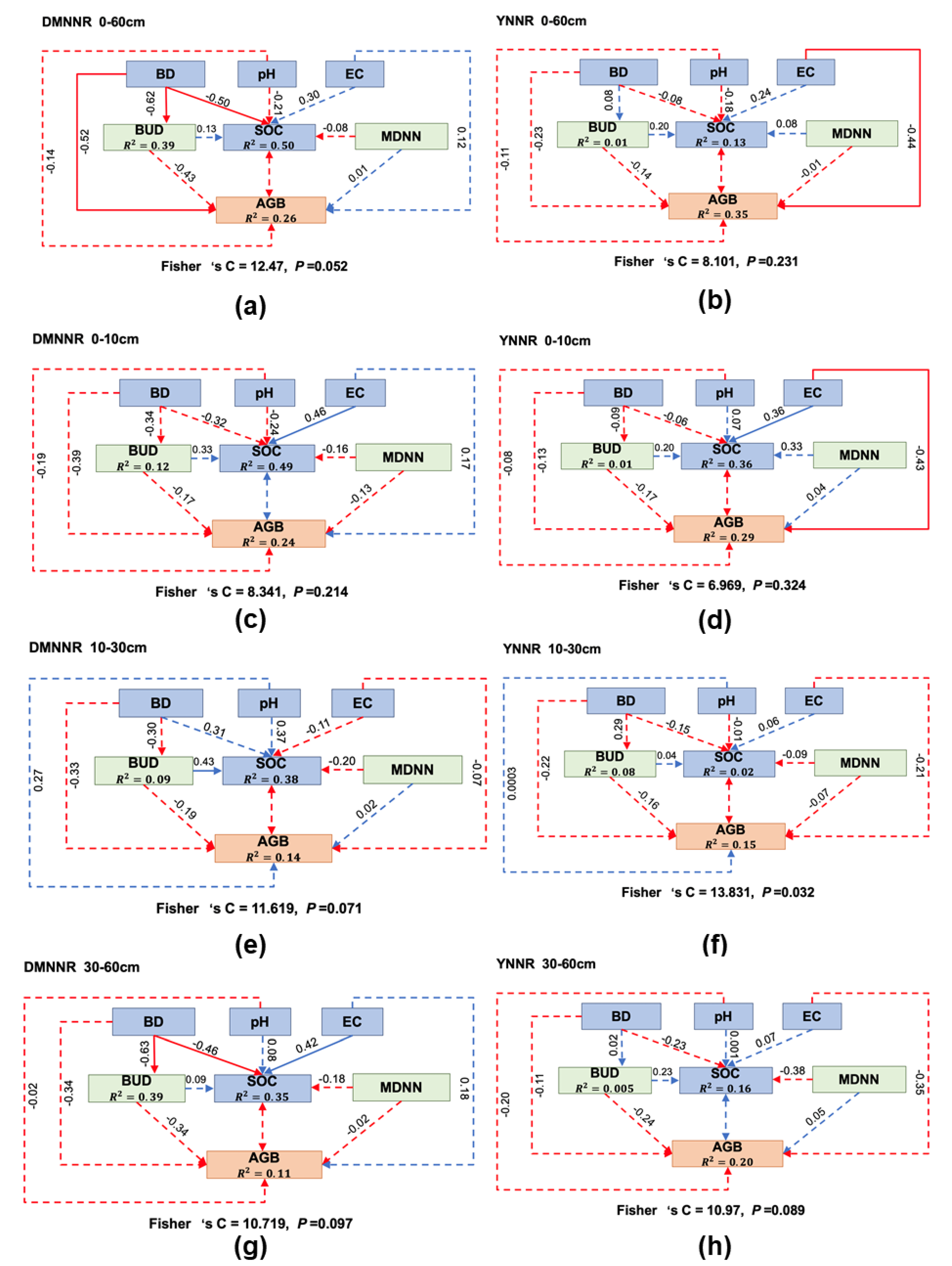
3. Results
3.1. Between-Site Difference in SOC Distribution
3.2. Multiple Regression Analyses
3.3. Inferring Complex Interactions from Structural Equation Modeling
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Millennium Ecosystem Assessment. Ecosystems and Human Well-Being; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Mitra, S.; Wassmann, R.; Vlek, P.L. An appraisal of global wetland area and its organic carbon stock. Curr. Sci. 2005, 88, 25–35. [Google Scholar]
- Mitsch, W.; Gosselink, J. Wetlands, 4th ed.; John Wiley & Sons: New York, NY, USA, 2007. [Google Scholar]
- Macreadie, P.I.; Anton, A.; Raven, J.A.; Beaumont, N.; Connolly, R.M.; Friess, D.A.; Kelleway, J.J.; Kennedy, H.; Kuwae, T.; Lavery, P.S. The future of blue carbon science. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivak, A.C.; Sanderman, J.; Bowen, J.L.; Canuel, E.A.; Hopkinson, C.S. Global-change controls on soil-carbon accumulation and loss in coastal vegetated ecosystems. Nat. Geosci. 2019, 12, 685–692. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, X.; Lee, S. Updated estimates of carbon accumulation rates in coastal marsh sediments. Biogeosciences 2014, 11, 5057–5071. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Dong, H.; Li, Y.; Wu, H.; Guan, B.; Gao, Y.; Zhou, D.; Wang, Y. Spatiotemporal distribution characteristics of soil organic carbon in newborn coastal wetlands of the Yellow River Delta Estuary. CLEAN Soil Air Water 2014, 42, 311–318. [Google Scholar] [CrossRef]
- Chmura, G.L. What do we need to assess the sustainability of the tidal salt marsh carbon sink? Ocean Coast Manage. 2013, 83, 25–31. [Google Scholar] [CrossRef]
- Li, Y.-L.; Wang, L.; Zhang, W.-Q.; Zhang, S.-P.; Wang, H.-L.; Fu, X.-H.; Le, Y.-Q. Variability of soil carbon sequestration capability and microbial activity of different types of salt marsh soils at Chongming Dongtan. Ecol. Eng. 2010, 36, 1754–1760. [Google Scholar] [CrossRef]
- Eswaran, H.; Van Den Berg, E.; Reich, P. Organic carbon in soils of the world. Soil. Sci. Soc. Am. J. 1993, 57, 192–194. [Google Scholar] [CrossRef] [Green Version]
- Bradley, R.; Milne, R.; Bell, J.; Lilly, A.; Jordan, C.; Higgins, A. A soil carbon and land use database for the United Kingdom. Soil Use Manage. 2005, 21, 363–369. [Google Scholar] [CrossRef]
- Meersmans, J.; De Ridder, F.; Canters, F.; De Baets, S.; Van Molle, M. A multiple regression approach to assess the spatial distribution of Soil Organic Carbon (SOC) at the regional scale (Flanders, Belgium). Geoderma 2008, 143, 1–13. [Google Scholar] [CrossRef]
- Morrissey, E.M.; Gillespie, J.L.; Morina, J.C.; Franklin, R.B. Salinity affects microbial activity and soil organic matter content in tidal wetlands. Global Change Biol. 2014, 20, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Osland, M.J.; Gabler, C.A.; Grace, J.B.; Day, R.H.; McCoy, M.L.; McLeod, J.L.; From, A.S.; Enwright, N.M.; Feher, L.C.; Stagg, C.L. Climate and plant controls on soil organic matter in coastal wetlands. Global Change Biol. 2018, 24, 5361–5379. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Wang, J.; Yan, D.; Gao, H.; Xiao, R.; Shao, H.; Ding, Q. Spatial and temporal distributions of soil organic carbon and total nitrogen in two marsh wetlands with different flooding frequencies of the Yellow River Delta, China. CLEAN Soil. Air. Water. 2012, 40, 1137–1144. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, G.; Zhao, Q.; Lu, Q.; Jia, J.; Cui, B.; Liu, X. Depth-Distribution patterns and control of soil organic carbon in coastal salt marshes with different plant covers. Sci. Rep. 2016, 6, 34835. [Google Scholar] [CrossRef]
- Yu, J.; Wang, Y.; Li, Y.; Dong, H.; Zhou, D.; Han, G.; Wu, H.; Wang, G.; Mao, P.; Gao, Y. Soil organic carbon storage changes in coastal wetlands of the modern Yellow River Delta from 2000 to 2009. Biogeosciences 2012, 9, 2325–2331. [Google Scholar] [CrossRef] [Green Version]
- Van der Putten, W.H.; de Ruiter, P.C.; Bezemer, T.M.; Harvey, J.A.; Wassen, M.; Wolters, V. Trophic interactions in a changing world. Basic Appl. Ecol. 2004, 5, 487–494. [Google Scholar] [CrossRef]
- He, Q.; Silliman, B.R.; Liu, Z.; Cui, B. Natural enemies govern ecosystem resilience in the face of extreme droughts. Ecol. Lett. 2017, 20, 194–201. [Google Scholar] [CrossRef]
- Silliman, B.R.; He, Q. Physical stress, consumer control, and new theory in ecology. Trends Ecol. Evol. 2018, 33, 492–503. [Google Scholar] [CrossRef]
- Liu, Q.-X.; Doelman, A.; Rottschäfer, V.; de Jager, M.; Herman, P.M.; Rietkerk, M.; van de Koppel, J. Phase separation explains a new class of self-organized spatial patterns in ecological systems. Proc. Natl. Acad. Sci. USA 2013, 110, 11905–11910. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.-X.; Herman, P.M.; Mooij, W.M.; Huisman, J.; Scheffer, M.; Olff, H.; Van De Koppel, J. Pattern formation at multiple spatial scales drives the resilience of mussel bed ecosystems. Nat. Commun. 2014, 5, 5234. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.-X.; Xu, C.; Ge, Z.-M.; van de Koppel, J.; Liu, Q.-X. The shaping role of self-organization: Linking vegetation patterning, plant traits and ecosystem functioning. Proc. R Soc. B. 2019, 286, 20182859. [Google Scholar] [CrossRef]
- Xu, C.; Van Nes, E.H.; Holmgren, M.; Kéfi, S.; Scheffer, M. Local facilitation may cause tipping points on a landscape level preceded by early-warning indicators. Am. Nat. 2015, 186, E81–E90. [Google Scholar] [CrossRef]
- Xu, C.; Holmgren, M.; Van Nes, E.H.; Maestre, F.T.; Soliveres, S.; Berdugo, M.; Kéfi, S.; Marquet, P.A.; Abades, S.; Scheffer, M. Can we infer plant facilitation from remote sensing? A test across global drylands. Ecol. Appl. 2015, 25, 1456–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Silliman, B.R. Consumer control as a common driver of coastal vegetation worldwide. Ecol. Monogr. 2016, 86, 278–294. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Zhu, H.; Qin, P.; Wang, H. Functional group classification and target species selection for Yancheng Nature Reserve, China. Biodivers. Conserv. 2004, 13, 1335–1353. [Google Scholar] [CrossRef]
- Li, S.; Cui, B.; Xie, T.; Bai, J.; Wang, Q.; Shi, W. What drives the distribution of crab burrows in different habitats of intertidal salt marshes, Yellow River Delta, China. Ecol. Indic. 2018, 92, 99–106. [Google Scholar] [CrossRef]
- Andreetta, A.; Fusi, M.; Cameldi, I.; Cimò, F.; Carnicelli, S.; Cannicci, S. Mangrove carbon sink. Do burrowing crabs contribute to sediment carbon storage? Evidence from a Kenyan mangrove system. J. Sea Res. 2014, 85, 524–533. [Google Scholar] [CrossRef] [Green Version]
- Tao, T.; Abades, S.; Teng, S.; Huang, Z.Y.; Reino, L.; Chen, B.J.; Zhang, Y.; Xu, C.; Svenning, J.-C. Macroecological factors shape local-scale spatial patterns in agriculturalist settlements. Proc. R Soc. B 2017, 284, 20172003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldridge, D.J.; Delgado-Baquerizo, M.; Quero, J.L.; Ochoa, V.; Gozalo, B.; García-Palacios, P.; Escolar, C.; García-Gómez, M.; Prina, A.; Bowker, M.A. Surface indicators are correlated with soil multifunctionality in global drylands. J. Appl. Ecol. 2020, 57, 424–435. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 3—Chemical. Methods, 3rd ed.; Sparks, D., Page, A., Helmke, P., Loeppert, R., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; Soil Science Society of America—American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Whittingham, M.J.; Stephens, P.A.; Bradbury, R.B.; Freckleton, R.P. Why do we still use stepwise modelling in ecology and behaviour? J. Anim. Ecol. 2006, 75, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, A.; Fujiwara, S.-I.; Niiya, H.; Katsuragi, H. Physical constraints on sand crab burrows: Mechanical properties of wet sand explain the size and spatial distributions of burrows on beaches. PLoS ONE 2019, 14, e0215743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Lefcheck, J.S. PIECEWISESEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- He, Q.; Silliman, B.R.; van de Koppel, J.; Cui, B. Weather fluctuations affect the impact of consumers on vegetation recovery following a catastrophic die-off. Ecology 2019, 100, e02559. [Google Scholar] [CrossRef] [PubMed]
- Sherman, P.M. Effects of land crabs on leaf litter distributions and accumulations in a mainland tropical rain forest. Biotropica 2003, 35, 365–374. [Google Scholar] [CrossRef]
- Milns, I.; Beale, C.M.; Smith, V.A. Revealing ecological networks using Bayesian network inference algorithms. Ecology 2010, 91, 1892–1899. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | DMNNR | YNNR | ||||
|---|---|---|---|---|---|---|
| Soil Depth | 0–10 cm | 10–30 cm | 30–60 cm | 0–10 cm | 10–30 cm | 30–60 cm |
| BD | −0.230.18 | 0.290.21 | −0.51 0.23 * | −0.060.19 | −0.010.25 | −0.210.20 |
| pH | −0.200.16 | −0.110.20 | 0.080.20 | 0.070.19 | 0.020.26 | 0.090.25 |
| EC | 0.430.16 * | 0.400.20 † | 0.450.20 * | 0.350.19 † | −0.120.23 | 0.170.27 |
| AGB | 0.220.16 | −0.060.18 | −0.160.17 | −0.020.20 | −0.160.22 | 0.070.22 |
| BUD | 0.370.17 * | 0.420.20 * | 0.030.25 | 0.200.18 | −0.030.24 | 0.230.22 |
| MDNN | −0.140.16 | −0.200.20 | −0.180.19 | 0.330.18 † | −0.120.26 | −0.440.25 † |
| Adjusted | 0.40 | 0.22 | 0.21 | 0.19 | 0.00 | 0.00 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; Miao, X.-Y.; Wang, B.; Jiang, R.-P.; Wen, T.; Liu, M.-S.; Huang, C.; Xu, C. System-Specific Complex Interactions Shape Soil Organic Carbon Distribution in Coastal Salt Marshes. Int. J. Environ. Res. Public Health 2020, 17, 2037. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17062037
Yang D, Miao X-Y, Wang B, Jiang R-P, Wen T, Liu M-S, Huang C, Xu C. System-Specific Complex Interactions Shape Soil Organic Carbon Distribution in Coastal Salt Marshes. International Journal of Environmental Research and Public Health. 2020; 17(6):2037. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17062037
Chicago/Turabian StyleYang, Dan, Xin-Yu Miao, Bo Wang, Ren-Ping Jiang, Teng Wen, Mao-Song Liu, Cheng Huang, and Chi Xu. 2020. "System-Specific Complex Interactions Shape Soil Organic Carbon Distribution in Coastal Salt Marshes" International Journal of Environmental Research and Public Health 17, no. 6: 2037. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17062037