
A Review on Bacterial Contribution to Lignocellulose Breakdown into Useful Bio-Products




Abstract
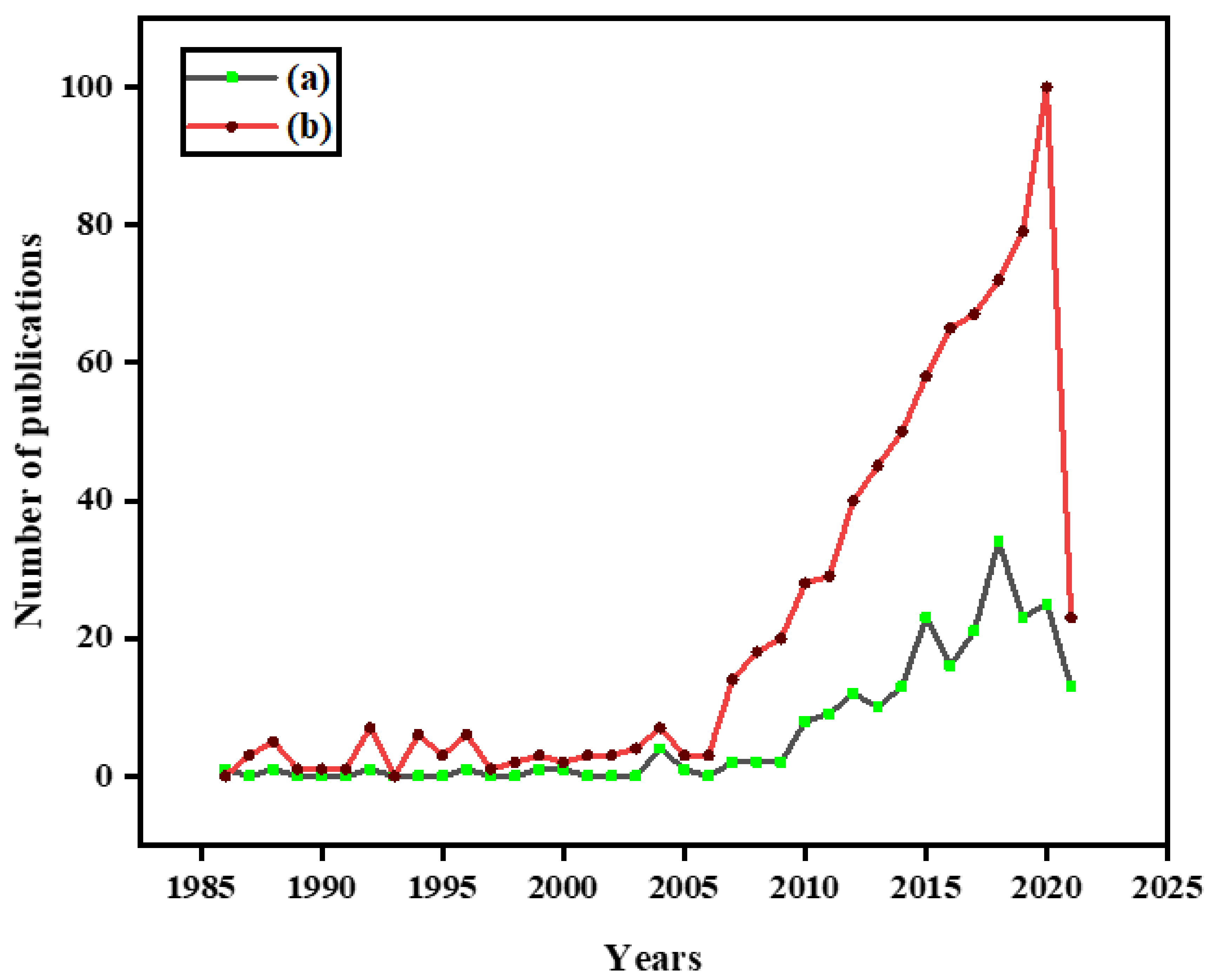
:1. Introduction
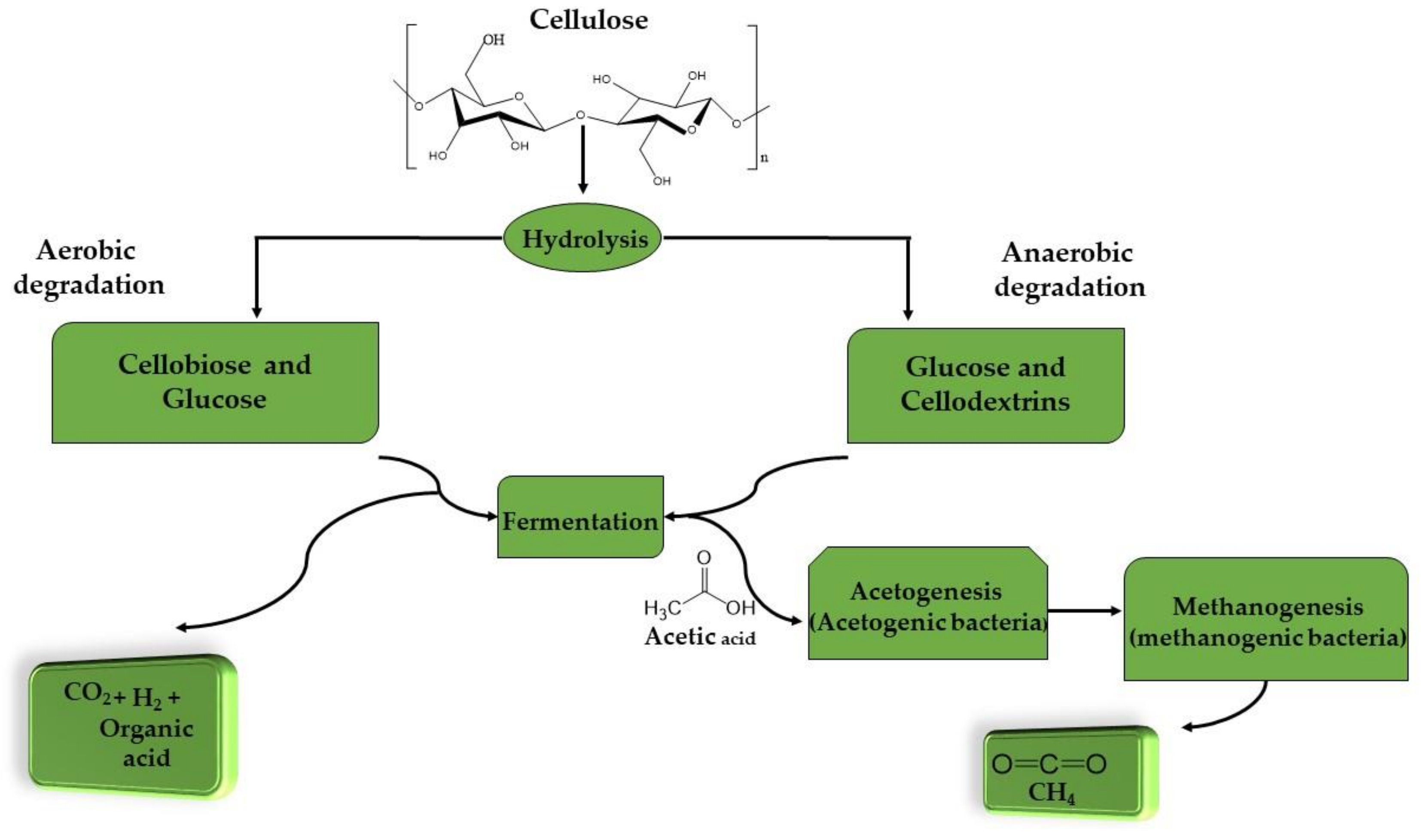
2. Bacteria Mechanism for Delignification and Cellulose Degradation
- Cellulose being oxidized to form CO2 and protons being concurrently reduced to H2;
- H2 being oxidized as CO2 is reduced to acetate and then methane, sulfate to hydrogen sulphide or nitrate reduced to ammonia.
3. Distribution of Lignocellulolytic Bacteria
4. Classification and Production of Bacterial Lignocellulases
5. Common Substrates for Lignocellulose Based Bio-Refineries
6. Bio-Refinery Products
7. Molecular Approaches of Bacterial Delignification and Cellulolytic Production
8. Challenges and Future Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Ge, X.; Chang, C.; Zhang, L.; Cui, S.; Luo, X.; Hu, S.; Qin, Y.; Li, Y. Conversion of Lignocellulosic Biomass Into Platform Chemicals for Biobased Polyurethane Application. In Advances in Bioenergy; Elsevier: Amsterdam, The Netherlands, 2018; Volume 3, pp. 161–213. [Google Scholar]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: A sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.H.; Xia, X.; Lin, C.X.; Tong, D.S.; Beltramini, J. Catalytic conversion of lignocellulosic biomass to fine chemicals and fuels. Chem. Soc. Rev. 2011, 40, 5588–5617. [Google Scholar] [CrossRef] [PubMed]
- Qian, E.W. Pretreatment and Saccharification of Lignocellulosic Biomass. In Research Approaches to Sustainable Biomass Systems; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 181–204. ISBN 9780124046092. [Google Scholar]
- Kumar, R.; Singh, S.; Singh, O.V. Bioconversion of lignocellulosic biomass: Biochemical and molecular perspectives. J. Ind. Microbiol. Biotechnol. 2008, 35, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Chukwuma, O.B.; Rafatullah, M.; Tajarudin, H.A.; Ismail, N. Lignocellulolytic Enzymes in Biotechnological and Industrial Processes: A Review. Sustainability 2020, 12, 7282. [Google Scholar] [CrossRef]
- Wongwilaiwalin, S.; Rattanachomsri, U.; Laothanachareon, T.; Eurwilaichitr, L.; Igarashi, Y.; Champreda, V. Analysis of a thermophilic lignocellulose degrading microbial consortium and multi-species lignocellulolytic enzyme system. Enzyme Microb. Technol. 2010, 47, 283–290. [Google Scholar] [CrossRef]
- Prakash, D.; Nawani, N.; Prakash, M.; Bodas, M.; Mandal, A.; Khetmalas, M.; Kapadnis, B. Actinomycetes: A repertory of green catalysts with a potential revenue resource. Biomed Res. Int. 2013, 2013, 264020. [Google Scholar] [CrossRef]
- Pathak, V.M.; Navneet. Review on the current status of polymer degradation: A microbial approach. Bioresour. Bioprocess. 2017, 4, 15. [Google Scholar] [CrossRef]
- Muaaz-Us-Salam, S.; Cleall, P.J.; Harbottle, M.J. Application of enzymatic and bacterial biodelignification systems for enhanced breakdown of model lignocellulosic wastes. Sci. Total Environ. 2020, 728, 138741. [Google Scholar] [CrossRef]
- DeAngelis, K.M.; Sharma, D.; Varney, R.; Simmons, B.; Isern, N.G.; Markilllie, L.M.; Nicora, C.; Norbeck, A.D.; Taylor, R.C.; Aldrich, J.T.; et al. Evidence supporting dissimilatory and assimilatory lignin degradation in Enterobacter lignolyticus SCF1. Front. Microbiol. 2013, 4, 280. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.A.D.; Kerr, R.G.; Haltli, B.A.; Tinto, W.F. Temperature and pH effect on glucose production from pretreated bagasse by a novel species of Citrobacter and other bacteria. Heliyon 2018, 4, e00657. [Google Scholar] [CrossRef]
- Hu, J.; Xue, Y.; Guo, H.; Gao, M.T.; Li, J.; Zhang, S.; Tsang, Y.F. Design and composition of synthetic fungal-bacterial microbial consortia that improve lignocellulolytic enzyme activity. Bioresour. Technol. 2017, 227, 247–255. [Google Scholar] [CrossRef]
- Arnling Bååth, J.; Mazurkewich, S.; Knudsen, R.M.; Poulsen, J.C.N.; Olsson, L.; Lo Leggio, L.; Larsbrink, J. Biochemical and structural features of diverse bacterial glucuronoyl esterases facilitating recalcitrant biomass conversion. Biotechnol. Biofuels 2018, 11, 213. [Google Scholar] [CrossRef]
- Cortes-Tolalpa, L.; Norder, J.; van Elsas, J.D.; Falcao Salles, J. Halotolerant microbial consortia able to degrade highly recalcitrant plant biomass substrate. Appl. Microbiol. Biotechnol. 2018, 102, 2913–2927. [Google Scholar] [CrossRef] [Green Version]
- López-Mondéjar, R.; Algora, C.; Baldrian, P. Lignocellulolytic systems of soil bacteria: A vast and diverse toolbox for biotechnological conversion processes. Biotechnol. Adv. 2019, 37, 107374. [Google Scholar] [CrossRef]
- Rosnow, J.J.; Anderson, L.N.; Nair, R.N.; Baker, E.S.; Wright, A.T. Profiling microbial lignocellulose degradation and utilization by emergent omics technologies. Crit. Rev. Biotechnol. 2017, 37, 626–640. [Google Scholar] [CrossRef]
- DeAngelis, K.M.; Allgaier, M.; Chavarria, Y.; Fortney, J.L.; Hugenholtz, P.; Simmons, B.; Sublette, K.; Silver, W.L.; Hazen, T.C. Characterization of Trapped Lignin-Degrading Microbes in Tropical Forest Soil. PLoS ONE 2011, 6, e19306. [Google Scholar] [CrossRef] [Green Version]
- Malgas, S.; Thoresen, M.; van Dyk, J.S.; Pletschke, B.I. Time dependence of enzyme synergism during the degradation of model and natural lignocellulosic substrates. Enzyme Microb. Technol. 2017, 103, 1–11. [Google Scholar] [CrossRef]
- Daniel, G.; Nilsson, T. Developments in the study of soft rot and bacterial decay. In Forest Products Biotechnology; CRC Press: Boca Raton, FL, USA, 1998; pp. 37–62. ISBN 9783540321385. [Google Scholar]
- Vijayalaxmi, S.; Anu Appaiah, K.A.; Jayalakshmi, S.K.; Mulimani, V.H.; Sreeramulu, K. Production of bioethanol from fermented sugars of sugarcane bagasse produced by lignocellulolytic enzymes of exiguobacterium sp. VSG-1. Appl. Biochem. Biotechnol. 2013, 171, 246–260. [Google Scholar] [CrossRef]
- Ho, B.K.X.; Azahari, B.; Yhaya, M.F.B.; Talebi, A.; Ng, C.W.C.; Tajarudin, H.A.; Ismail, N. Green Technology Approach for Reinforcement of Calcium Chloride Cured Sodium Alginate Films by Isolated Bacteria from Palm Oil Mill Effluent (POME). Sustainability 2020, 12, 9468. [Google Scholar] [CrossRef]
- Taha, M.; Kadali, K.K.; Khalid, A.H.; Smith, A.T.; Ball, A.S.; Adetutu, E.M. An effective microplate method (Biolog MT2) for screening native lignocellulosic-straw-degrading bacteria. Ann. Microbiol. 2015, 65, 2053–2064. [Google Scholar] [CrossRef] [Green Version]
- McDonald, J.E.; Rooks, D.J.; McCarthy, A.J. Methods for the isolation of cellulose-degrading microorganisms. In Methods in Enzymology; Academic Press Inc.: Cambridge, MA, USA, 2012; Volume 510, pp. 349–374. [Google Scholar]
- Liao, J.J.; Latif, N.H.A.; Trache, D.; Brosse, N.; Hussin, M.H. Current advancement on the isolation, characterization and application of lignin. Int. J. Biol. Macromol. 2020, 162, 985–1024. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, M.; Bae, J.-H.; Sohn, J.-H.; Sung, B.H. Bacterial Valorization of Lignin: Strains, Enzymes, Conversion Pathways, Biosensors, and Perspectives. Front. Bioeng. Biotechnol. 2019, 7, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Berg, B.; Laskowski, R. Decomposers: Soil Microorganisms and Animals. Adv. Ecol. Res. 2005, 38, 73–100. [Google Scholar]
- Xie, S.; Ragauskas, A.J.; Yuan, J.S. Lignin Conversion: Opportunities and Challenges for the Integrated Biorefinery. Ind. Biotechnol. 2016, 12, 161–167. [Google Scholar] [CrossRef]
- Xu, R.; Zhang, K.; Liu, P.; Han, H.; Zhao, S.; Kakade, A.; Khan, A.; Du, D.; Li, X. Lignin depolymerization and utilization by bacteria. Bioresour. Technol. 2018, 269, 557–566. [Google Scholar] [CrossRef]
- Ljungdahl, L.G.; Eriksson, K.-E. Ecology of Microbial Cellulose Degradation. In Advances in Microbial Ecology; Springer: Berlin, Germany, 1985; pp. 237–299. [Google Scholar] [CrossRef]
- Ezeilo, U.R.; Zakaria, I.I.; Huyop, F.; Wahab, R.A. Enzymatic breakdown of lignocellulosic biomass: The role of glycosyl hydrolases and lytic polysaccharide monooxygenases. Biotechnol. Biotechnol. Equip. 2017, 31, 647–662. [Google Scholar] [CrossRef] [Green Version]
- Mohee, R.; Unmar, G.D.; Mudhoo, A.; Khadoo, P. Biodegradability of biodegradable/degradable plastic materials under aerobic and anaerobic conditions. Waste Manag. 2008, 28, 1624–1629. [Google Scholar] [CrossRef]
- Singhvi, M.S.; Gokhale, D.V. Lignocellulosic biomass: Hurdles and challenges in its valorization. Appl. Microbiol. Biotechnol. 2019, 103, 9305–9320. [Google Scholar] [CrossRef]
- Hassan, S.S.; Williams, G.A.; Jaiswal, A.K. Moving towards the second generation of lignocellulosic biorefineries in the EU: Drivers, challenges, and opportunities. Renew. Sustain. Energy Rev. 2019, 101, 590–599. [Google Scholar] [CrossRef] [Green Version]
- Beaton, D.; Pelletier, P.; Goulet, R.R. Microbial degradation of cellulosic material and gas generation: Implications for the management of low-and intermediate-level radioactive waste. Front. Microbiol. 2019, 10, 204. [Google Scholar] [CrossRef] [Green Version]
- Panda, A.K.; Bisht, S.S.; De Mandal, S.; Kumar, N.S. Microbial Diversity of Thermophiles through the Lens of Next Generation Sequencing. In Microbial Diversity in the Genomic Era; Elsevier: Amsterdam, The Netherlands, 2019; pp. 217–226. [Google Scholar]
- Chang, T.; Yao, S. Thermophilic, lignocellulolytic bacteria for ethanol production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2011, 92, 13–27. [Google Scholar] [CrossRef]
- Escuder-Rodríguez, J.-J.; DeCastro, M.-E.; Cerdán, M.-E.; Rodríguez-Belmonte, E.; Becerra, M.; González-Siso, M.-I. Cellulases from Thermophiles Found by Metagenomics. Microorganisms 2018, 6, 66. [Google Scholar] [CrossRef] [Green Version]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial Diversity in Extreme Marine Habitats and Their Biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Hemati, A.; Aliasgharzad, N.; Khakvar, R. Biocatalysis and Agricultural Biotechnology In vitro evaluation of lignocellulolytic activity of thermophilic bacteria isolated from di ff erent composts and soils of Iran. Biocatal. Agric. Biotechnol. 2018, 14, 424–430. [Google Scholar] [CrossRef]
- Schiraldi, C.; De Rosa, M. Mesophilic Organisms. In Encyclopedia of Membranes; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–2. [Google Scholar]
- Varma, V.S.; Das, S.; Sastri, C.V.; Kalamdhad, A.S. Microbial degradation of lignocellulosic fractions during drum composting of mixed organic waste. Sustain. Environ. Res. 2017, 27, 265–272. [Google Scholar] [CrossRef]
- Kanokratana, P.; Wongwilaiwalin, S.; Mhuantong, W.; Tangphatsornruang, S.; Eurwilaichitr, L.; Champreda, V. Characterization of cellulolytic microbial consortium enriched on Napier grass using metagenomic approaches. J. Biosci. Bioeng. 2018, 125, 439–447. [Google Scholar] [CrossRef]
- Zhu, D.; Adebisi, W.A.; Ahmad, F.; Sethupathy, S.; Danso, B.; Sun, J. Recent Development of Extremophilic Bacteria and Their Application in Biorefinery. Front. Bioeng. Biotechnol. 2020, 8, 483. [Google Scholar] [CrossRef]
- Björdal, C.G.; Dayton, P.K. First evidence of microbial wood degradation in the coastal waters of the Antarctic. Sci. Rep. 2020, 10, 12774. [Google Scholar] [CrossRef]
- Kong, X.; Du, J.; Ye, X.; Xi, Y.; Jin, H.; Zhang, M.; Guo, D. Enhanced methane production from wheat straw with the assistance of lignocellulolytic microbial consortium TC-5. Bioresour. Technol. 2018, 263, 33–39. [Google Scholar] [CrossRef]
- Tomazetto, G.; Pimentel, A.C.; Wibberg, D.; Dixon, N.; Squina, F.M. Multi-omic Directed Discovery of Cellulosomes, Polysaccharide Utilization Loci, and Lignocellulases from an Enriched Rumen Anaerobic Consortium. Appl. Environ. Microbiol. 2020, 86, e00199-20. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Mishra, J.; Arora, N.; Mishra, P.; Li, H.; O’Hair, J.; Bhatti, S.; Zhou, S. Microbial cellulolytic enzymes: Diversity and biotechnology with reference to lignocellulosic biomass degradation. Rev. Environ. Sci. Bio Technol. 2020, 19, 621–648. [Google Scholar] [CrossRef]
- Duarah, P.; Haldar, D.; Purkait, M.K. Technological advancement in the synthesis and applications of lignin-based nanoparticles derived from agro-industrial waste residues: A review. Int. J. Biol. Macromol. 2020, 163, 1828–1843. [Google Scholar] [CrossRef] [PubMed]
- Datta, R.; Kelkar, A.; Baraniya, D.; Molaei, A.; Moulick, A.; Meena, R.S.; Formanek, P. Enzymatic degradation of lignin in soil: A review. Sustainability 2017, 9, 1163. [Google Scholar] [CrossRef] [Green Version]
- Baramee, S.; Uke, A.; Tachaapaikoon, C.; Waeonukul, R.; Pason, P.; Ratanakhanokchai, K.; Kosugi, A. Draft genome sequence data of Paenbacillus curdlanolyticus B-6 possessing a unique xylanolytic-cellulolytic multienzyme system. Data Br. 2020, 32, 106213. [Google Scholar] [CrossRef] [PubMed]
- Kausar, H.; Ismail, M.R.; Saud, H.M.; Habib, S.H.; Othman, R.; Miah, G. A novel lignocellulolytic bacterium for bioconversion of rice straw. Pakistan J. Agric. Sci. 2016, 53, 523–533. [Google Scholar] [CrossRef]
- Khelil, O.; Choubane, S.; Cheba, B.A. Polyphenols content of spent coffee grounds subjected to physico-chemical pretreatments influences lignocellulolytic enzymes production by Bacillus sp. R2. Bioresour. Technol. 2016, 211, 769–773. [Google Scholar] [CrossRef]
- Xu, J.; Gao, Z.; Wu, B.; He, B. Lactose-inducted production of a complete lignocellulolytic enzyme system by a novel bacterium Bacillus sp. BS-5 and its application for saccharification of alkali-pretreated corn cob. Cellulose 2017, 24, 2059–2070. [Google Scholar] [CrossRef]
- Gong, G.; Kim, S.; Lee, S.M.; Woo, H.M.; Park, T.H.; Um, Y. Complete genome sequence of Bacillus sp. 275, producing extracellular cellulolytic, xylanolytic and ligninolytic enzymes. J. Biotechnol. 2017, 254, 59–62. [Google Scholar] [CrossRef]
- Suriya, J.; Bharathiraja, S.; Manivasagan, P.; Kim, S.K. Enzymes from Rare Actinobacterial Strains. In Advances in Food and Nutrition Research; Academic Press Inc.: Cambridge, MA, USA, 2016; Volume 79, pp. 67–98. [Google Scholar]
- Conway, J.M.; Crosby, J.R.; Hren, A.P.; Southerland, R.T.; Lee, L.L.; Lunin, V.V.; Alahuhta, P.; Himmel, M.E.; Bomble, Y.J.; Adams, M.W.W.; et al. Novel multidomain, multifunctional glycoside hydrolases from highly lignocellulolytic Caldicellulosiruptor species. AIChE J. 2018, 64, 4218–4228. [Google Scholar] [CrossRef]
- Hassan, S.S.; Williams, G.A.; Jaiswal, A.K. Emerging technologies for the pretreatment of lignocellulosic biomass. Bioresour. Technol. 2018, 262, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Arevalo-Gallegos, A.; Ahmad, Z.; Asgher, M.; Parra-Saldivar, R.; Iqbal, H.M.N. Lignocellulose: A sustainable material to produce value-added products with a zero waste approach—A review. Int. J. Biol. Macromol. 2017, 99, 308–318. [Google Scholar] [CrossRef]
- Arora, R.; Behera, S.; Kumar, S. Bioprospecting thermophilic/thermotolerant microbes for production of lignocellulosic ethanol: A future perspective. Renew. Sustain. Energy Rev. 2015, 51, 699–717. [Google Scholar] [CrossRef]
- Ghosh, S.; Chowdhury, R.; Bhattacharya, P. Sustainability of cereal straws for the fermentative production of second generation biofuels: A review of the efficiency and economics of biochemical pretreatment processes. Appl. Energy 2017, 198, 284–298. [Google Scholar] [CrossRef]
- Deng, Y.J.; Wang, S.Y. Synergistic growth in bacteria depends on substrate complexity. J. Microbiol. 2016, 54, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A.S.; Bhattacharya, A.; Pletschke, B.I. Synergism of fungal and bacterial cellulases and hemicellulases: A novel perspective for enhanced bio-ethanol production. Biotechnol. Lett. 2015, 37, 1117–1129. [Google Scholar] [CrossRef]
- Levin, D.B.; Verbeke, T.J.; Munir, R.; Islam, R.; Ramachandran, U.; Lal, S.; Schellenberg, J.; Sparling, R. Omics Approaches for Designing Biofuel Producing Cocultures for Enhanced Microbial Conversion of Lignocellulosic Substrates. In Direct Microbial Conversion of Biomass to Advanced Biofuels; Elsevier: Amsterdam, The Netherlands, 2015; pp. 335–363. ISBN 9780444595928. [Google Scholar]
- Sander, K.; Yeary, M.; Mahan, K.; Whitham, J.; Giannone, R.J.; Brown, S.D.; Rodriguez, M.; Graham, D.E.; Hankoua, B. Expression of benzoyl-CoA metabolism genes in the lignocellulolytic host Caldicellulosiruptor bescii. AMB Express 2019, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.J.; Wang, S.Y. Complex carbohydrates reduce the frequency of antagonistic interactions among bacteria degrading cellulose and xylan. FEMS Microbiol. Lett. 2017, 364, fnx019. [Google Scholar] [CrossRef] [Green Version]
- Chin, K.L.; Nurliyana, M.Y.; H’ng, P.S.; Lee, C.L.; Go, W.Z.; Khoo, P.S.; Raja Nazrin, R.A.; Ashikin, S.N. Effects of Bacterial Bio-augmentation on the Methane Potential from Facultative Digestion of Palm Oil Mill Effluent and Empty Fruit Bunch. Waste Biomass Valoriz. 2020, 11, 3407–3418. [Google Scholar] [CrossRef]
- Zheng, G.; Yin, T.; Lu, Z.; yannick benz Boboua, S.; Li, J.; Zhou, W. Degradation of rice straw at low temperature using a novel microbial consortium LTF-27 with efficient ability. Bioresour. Technol. 2020, 304, 123064. [Google Scholar] [CrossRef]
- Qi, X.; Zheng, Y.; Tang, N.; Zhou, J.; Sun, S. Bioconversion of citrus peel wastes into bioflocculants and their application in the removal of microcystins. Sci. Total Environ. 2020, 715, 136885. [Google Scholar] [CrossRef]
- Yao, Q.; Huang, M.; Bu, Z.; Zeng, J.; Wang, X.; Liu, Z.; Ma, J.; Zhang, K.; Liu, X.; Zhu, Y. Identification and characterization of a novel bacterial carbohydrate esterase from the bacterium Pantoea ananatis Sd-1 with potential for degradation of lignocellulose and pesticides. Biotechnol. Lett. 2020, 42, 1479–1488. [Google Scholar] [CrossRef] [PubMed]
- Taha, M.; Shahsavari, E.; Al-Hothaly, K.; Mouradov, A.; Smith, A.T.; Ball, A.S.; Adetutu, E.M. Enhanced Biological Straw Saccharification Through Coculturing of Lignocellulose-Degrading Microorganisms. Appl. Biochem. Biotechnol. 2015, 175, 3709–3728. [Google Scholar] [CrossRef] [PubMed]
- Buraimoh, O.M.; Ilori, M.O.; Amund, O.O.; Michel, F.C.; Grewal, S.K. Assessment of bacterial degradation of lignocellulosic residues (sawdust) in a tropical estuarine microcosm using improvised floating raft equipment. Int. Biodeterior. Biodegrad. 2015, 104, 186–193. [Google Scholar] [CrossRef]
- Setiawati, T.C.; Mandala, M.; Avief. The activity of Potassium and Phosphate Solubilizing bacteria from sugarcane rhizosphere on Some Bagasse Condition media inoculated byLignocellulolytic bacteria. In IOP Conference Series: Earth and Environmental Science, Proceedings of the 6th International Conference on Sustainable Agriculture, Food and Energy, Manila, The Philippines, 18–21 October 2018; Institute of Physics Publishing: Bristol, UK, 2019; Volume 347. [Google Scholar]
- Neoh, C.H.; Lam, C.Y.; Ghani, S.M.; Ware, I.; Sarip, S.H.M.; Ibrahim, Z. Bioremediation of high-strength agricultural wastewater using Ochrobactrum sp. strain SZ1. 3 Biotech 2016, 6, 143. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Zhang, K.; Huang, M.; Hector, S.B.; Liu, B.; Tong, C.; Liu, Q.; Zeng, J.; Gao, Y.; Xu, T.; et al. Involvement of Fenton chemistry in rice straw degradation by the lignocellulolytic bacterium Pantoea ananatis Sd-1. Biotechnol. Biofuels 2016, 9, 211. [Google Scholar] [CrossRef] [Green Version]
- Gong, G.; Lee, S.M.; Woo, H.M.; Park, T.H.; Um, Y. Influences of Media Compositions on Characteristics of Isolated Bacteria Exhibiting Lignocellulolytic Activities from Various Environmental Sites. Appl. Biochem. Biotechnol. 2017, 183, 931–942. [Google Scholar] [CrossRef]
- Yang, Y.J.; Lin, W.; Singh, R.P.; Xu, Q.; Chen, Z.; Yuan, Y.; Zou, P.; Li, Y.; Zhang, C. Genomic, transcriptomic and enzymatic insight into lignocellulolytic system of a plant pathogen dickeya sp. WS52 to digest sweet pepper and tomato stalk. Biomolecules 2019, 9, 753. [Google Scholar] [CrossRef] [Green Version]
- Nutongkaew, T.; Prasertsan, P.; O-Thong, S.; Chanthong, S.; Suyotha, W. Improved Methane Production Using Lignocellulolytic Enzymes from Trichoderma koningiopsis TM3 Through Co-digestion of Palm Oil Mill Effluent and Oil Palm Trunk Residues. Waste Biomass Valoriz. 2020, 11, 5123–5136. [Google Scholar] [CrossRef]
- Dulermo, T.; Coze, F.; Virolle, M.J.; Méchin, V.; Baumberger, S.; Froissard, M. Bioconversion of agricultural lignocellulosic residues into branched-chain fatty acids using Streptomyces lividans. OCL 2016, 23, A202. [Google Scholar] [CrossRef] [Green Version]
- Detman, A.; Bucha, M.; Simoneit, B.R.T.; Mielecki, D.; Piwowarczyk, C.; Chojnacka, A.; Błaszczyk, M.K.; Jędrysek, M.O.; Marynowski, L.; Sikora, A. Lignite biodegradation under conditions of acidic molasses fermentation. Int. J. Coal Geol. 2018, 196, 274–287. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Yang, J.; Ji, L.; Liu, J.; Yang, Y.; Yuan, H. Metagenomic and metaproteomic analyses of a corn stover-adapted microbial consortium EMSD5 reveal its taxonomic and enzymatic basis for degrading lignocellulose. Biotechnol. Biofuels 2016, 9, 243. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Hong, C.; Zheng, B.; Lu, F.; Jiang, D.; Qin, W. Bioflocculants’ production in a biomass-degrading bacterium using untreated corn stover as carbon source and use of bioflocculants for microalgae harvest. Biotechnol. Biofuels 2017, 10, 306. [Google Scholar] [CrossRef] [Green Version]
- Bibra, M.; Kumar, S.; Wang, J.; Bhalla, A.; Salem, D.R.; Sani, R.K. Single pot bioconversion of prairie cordgrass into biohydrogen by thermophiles. Bioresour. Technol. 2018, 266, 232–241. [Google Scholar] [CrossRef]
- Saratale, G.D.; Saratale, R.G.; Ghodake, G.S.; Jiang, Y.Y.; Chang, J.S.; Shin, H.S.; Kumar, G. Solid state fermentative lignocellulolytic enzymes production, characterization and its application in the saccharification of rice waste biomass for ethanol production: An integrated biotechnological approach. J. Taiwan Inst. Chem. Eng. 2017, 76, 51–58. [Google Scholar] [CrossRef]
- Jain, A.; Pelle, H.S.; Baughman, W.H.; Henson, J.M. Conversion of ammonia-pretreated switchgrass to biofuel precursors by bacterial–fungal consortia under solid-state and submerged-state cultivation. J. Appl. Microbiol. 2017, 122, 953–963. [Google Scholar] [CrossRef]
- Medouni-Haroune, L.; Zaidi, F.; Medouni-Adrar, S.; Roussos, S.; Azzouz, S.; Desseaux, V.; Kecha, M. Selective isolation and screening of actinobacteria strains producing lignocellulolytic enzymes using olive pomace as substrate. Iran. J. Biotechnol. 2017, 15, 74–77. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Chen, H.; Fan, L.; Linklater, A.; Zheng, B.; Jiang, D.; Qin, W. Enzymes produced by biomass-degrading bacteria can efficiently hydrolyze algal cell walls and facilitate lipid extraction. Renew. Energy 2017, 109, 195–201. [Google Scholar] [CrossRef]
- Guo, H.; Wu, Y.; Hong, C.; Chen, H.; Chen, X.; Zheng, B.; Jiang, D.; Qin, W. Enhancing digestibility of Miscanthus using lignocellulolytic enzyme produced by Bacillus. Bioresour. Technol. 2017, 245, 1008–1015. [Google Scholar] [CrossRef]
- Solikhah, F.; Assavalapsakul, W.; Zulaika, E. Peat Endogenous Lignocellulolytic Bacteria for Humic Waste Decomposition. In Journal of Physics: Conference Series, Proceedings of the Mathematics, Informatics, Science and Education International Conference, Surabaya, Indonesia, 21 July 2018; Institute of Physics Publishing: Bristol, UK, 2018; Volume 1108. [Google Scholar]
- Yeager, C.M.; Gallegos-Graves, L.V.; Dunbar, J.; Hesse, C.N.; Daligault, H.; Kuske, C.R. Polysaccharide degradation capability of Actinomycetales soil isolates from a semiarid grassland of the Colorado Plateau. Appl. Environ. Microbiol. 2017, 83, e03020-16. [Google Scholar] [CrossRef] [Green Version]
- Aini, F.N.; Laconi, E.B.; Suhartono, M.T. The potential of agricultural by-products as a carbon source in the growth of Paenibacillus sp. In AIP Conference Proceedings; American Institute of Physics Inc.: College Park, MD, USA, 2018; Volume 2021, p. 030004. [Google Scholar]
- Nevita, T.; Sharma, G.D.; Pandey, P. Differences in rice rhizosphere bacterial community structure by application of lignocellulolytic plant-probiotic bacteria with rapid composting traits. Ecol. Eng. 2018, 120, 209–221. [Google Scholar] [CrossRef]
- Apiwatanapiwat, W.; Vaithanomsat, P.; Thanapase, W.; Ratanakhanokchai, K.; Kosugi, A. Xylan supplement improves 1,3-propanediol fermentation by Clostridium butyricum. J. Biosci. Bioeng. 2018, 125, 662–668. [Google Scholar] [CrossRef]
- Zhang, K.; Huang, M.; Ma, J.; Liu, Z.; Zeng, J.; Liu, X.; Xu, T.; Wang, X.; Liu, Y.; Bu, Z.; et al. Identification and characterization of a novel bacterial pyranose 2-oxidase from the lignocellulolytic bacterium Pantoea ananatis Sd-1. Biotechnol. Lett. 2018, 40, 871–880. [Google Scholar] [CrossRef]
- Bohra, V.; Dafale, N.A.; Purohit, H.J. Paenibacillus polymyxa ND25: Candidate genome for lignocellulosic biomass utilization. 3 Biotech 2018, 8, 248. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Q.; Zhang, Y.; Chen, J.; Zhang, D.; Tong, J. Changes in fibrolytic enzyme activity during vermicomposting of maize stover by an anecic earthworm Amynthas hupeiensis. Polym. Degrad. Stab. 2015, 120, 169–177. [Google Scholar] [CrossRef]
- Thiyageshwari, S.; Gayathri, P.; Krishnamoorthy, R.; Anandham, R.; Paul, D. Exploration of rice husk compost as an alternate organic manure to enhance the productivity of blackgram in typic haplustalf and typic rhodustalf. Int. J. Environ. Res. Public Health 2018, 15, 358. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Gu, W.; Xu, H.Y.; Yang, G.L.; Shan, X.F.; Chen, G.; Wang, C.F.; Qian, A.D. Complete genome sequence of Bacillus velezensis 157 isolated from Eucommia ulmoides with pathogenic bacteria inhibiting and lignocellulolytic enzymes production by SSF. 3 Biotech 2018, 8, 465. [Google Scholar] [CrossRef]
- Ozbayram, E.G.; Akyol, Ç.; Ince, B.; Karakoç, C.; Ince, O. Rumen bacteria at work: Bioaugmentation strategies to enhance biogas production from cow manure. J. Appl. Microbiol. 2018, 124, 491–502. [Google Scholar] [CrossRef]
- Wongwilaiwalin, S.; Mhuantong, W.; Champreda, V.; Tangphatsornruang, S.; Panichnumsin, P.; Ratanakhanokchai, K.; Tachaapaikoon, C. Structural and metabolic adaptation of cellulolytic microcosm in co-digested Napier grass-swine manure and its application in enhancing thermophilic biogas production. RSC Adv. 2018, 8, 29806–29815. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Zhang, H.; Sang, Y.; Cai, P.; Zhang, G.; Chen, Q. Feasibility of weeds-based compost-cultivated Agaricus bisporus. Chinese J. Appl. Environ. Biol. 2018, 24, 1275–1282. [Google Scholar] [CrossRef]
- Raut, M.P.; Couto, N.; Karunakaran, E.; Biggs, C.A.; Wright, P.C. Deciphering the unique cellulose degradation mechanism of the ruminal bacterium Fibrobacter succinogenes S85. Sci. Rep. 2019, 9, 16542. [Google Scholar] [CrossRef] [PubMed]
- Bohra, V.; Dafale, N.A.; Purohit, H.J. Understanding the alteration in rumen microbiome and CAZymes profile with diet and host through comparative metagenomic approach. Arch. Microbiol. 2019, 201, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Tapadia-Maheshwari, S.; Pore, S.; Engineer, A.; Shetty, D.; Dagar, S.S.; Dhakephalkar, P.K. Illustration of the microbial community selected by optimized process and nutritional parameters resulting in enhanced biomethanation of rice straw without thermo-chemical pretreatment. Bioresour. Technol. 2019, 289, 121639. [Google Scholar] [CrossRef]
- Wu, Y.; Guo, H.; Zhang, J.; Chen, X.; Wu, M.; Qin, W. Multiple Applications of Enzymes Induced by Algal Biomasses from a New Bacillus Isolate to Saccharify Algae and Degrade Chemical Dyes. Waste Biomass Valoriz. 2019, 10, 2517–2526. [Google Scholar] [CrossRef]
- Straub, C.T.; Khatibi, P.A.; Otten, J.K.; Adams, M.W.W.; Kelly, R.M. Lignocellulose solubilization and conversion by extremely thermophilic Caldicellulosiruptor bescii improves by maintaining metabolic activity. Biotechnol. Bioeng. 2019, 116, 1901–1908. [Google Scholar] [CrossRef]
- Joy, S.; Rahman, P.K.S.M.; Khare, S.K.; Sharma, S. Production and characterization of glycolipid biosurfactant from Achromobacter sp. (PS1) isolate using one-factor-at-a-time (OFAT) approach with feasible utilization of ammonia-soaked lignocellulosic pretreated residues. Bioprocess Biosyst. Eng. 2019, 42, 1301–1315. [Google Scholar] [CrossRef]
- Luo, C.; Li, Y.; Chen, Y.; Fu, C.; Long, W.; Xiao, X.; Liao, H.; Yang, Y. Bamboo lignocellulose degradation by gut symbiotic microbiota of the bamboo snout beetle Cyrtotrachelus buqueti. Biotechnol. Biofuels 2019, 12, 70. [Google Scholar] [CrossRef]
- Guo, W.; Guo, X.J.; Zhu, B.C.; Guo, Y.Y.; Zhou, X. In situ degradation, ruminal fermentation, and the rumen bacterial community of cattle fed corn stover fermented by lignocellulolytic microorganisms. Anim. Feed Sci. Technol. 2019, 248, 10–19. [Google Scholar] [CrossRef]
- Neelkant, K.S.; Shankar, K.; Jayalakshmi, S.K.; Sreeramulu, K. Optimization of conditions for the production of lignocellulolytic enzymes by Sphingobacterium sp. ksn-11 utilizing agro-wastes under submerged condition. Prep. Biochem. Biotechnol. 2019, 49, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Zabidi, N.A.M.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Rahim, R.A. Enhancement of versatile extracellular cellulolytic and hemicellulolytic enzyme productions by Lactobacillus plantarum RI 11 isolated from Malaysian food using renewable natural polymers. Molecules 2020, 25, 2607. [Google Scholar] [CrossRef]
- Arias, D.E.; Veluchamy, C.; Dunfield, K.E.; Habash, M.B.; Gilroyed, B.H. Hygienization and microbial metabolic adaptation during anaerobic co-digestion of swine manure and corn stover. Bioresour. Technol. 2020, 306, 123168. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.S.; Han, Y.; Wang, X.C.; Wen, J.; Cao, S.; Zhang, K.; Li, Q.; Yuan, H. Persistent action of cow rumen microorganisms in enhancing biodegradation of wheat straw by rumen fermentation. Sci. Total Environ. 2020, 715, 136529. [Google Scholar] [CrossRef] [PubMed]
- Froese, A.G.; Nguyen, T.N.; Ayele, B.T.; Sparling, R. Digestibility of Wheat and Cattail Biomass Using a Co-culture of Thermophilic Anaerobes for Consolidated Bioprocessing. Bioenergy Res. 2020, 13, 325–333. [Google Scholar] [CrossRef]
- González-García, S.; Bacenetti, J. Exploring the production of bio-energy from wood biomass. Italian case study. Sci. Total Environ. 2019, 647, 158–168. [Google Scholar] [CrossRef]
- Gasparatos, A.; Stromberg, P.; Takeuchi, K. Sustainability impacts of first-generation biofuels. Anim. Front. 2013, 3, 12–26. [Google Scholar] [CrossRef]
- Sharma, B.; Larroche, C.; Dussap, C.-G. Comprehensive assessment of 2G bioethanol production. Bioresour. Technol. 2020, 313, 123630. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.N.; Suely, A.; Boyce, A.N.; Faruq, G. Bioethanol production from renewable sources: Current perspectives and technological progress. Renew. Sustain. Energy Rev. 2017, 71, 475–501. [Google Scholar] [CrossRef]
- Carrillo-Nieves, D.; Rostro Alanís, M.J.; de la Cruz Quiroz, R.; Ruiz, H.A.; Iqbal, H.M.N.; Parra-Saldívar, R. Current status and future trends of bioethanol production from agro-industrial wastes in Mexico. Renew. Sustain. Energy Rev. 2019, 102, 63–74. [Google Scholar] [CrossRef]
- Angelidaki, I.; Treu, L.; Tsapekos, P.; Luo, G.; Campanaro, S.; Wenzel, H.; Kougias, P.G. Biogas upgrading and utilization: Current status and perspectives. Biotechnol. Adv. 2018, 36, 452–466. [Google Scholar] [CrossRef] [Green Version]
- Ullah Khan, I.; Hafiz Dzarfan Othman, M.; Hashim, H.; Matsuura, T.; Ismail, A.F.; Rezaei-DashtArzhandi, M.; Wan Azelee, I. Biogas as a renewable energy fuel—A review of biogas upgrading, utilisation and storage. Energy Convers. Manag. 2017, 150, 277–294. [Google Scholar] [CrossRef]
- Negri, C.; Ricci, M.; Zilio, M.; D’Imporzano, G.; Qiao, W.; Dong, R.; Adani, F. Anaerobic digestion of food waste for bio-energy production in China and Southeast Asia: A review. Renew. Sustain. Energy Rev. 2020, 133, 110138. [Google Scholar] [CrossRef]
- Molino, A.; Nanna, F.; Ding, Y.; Bikson, B.; Braccio, G. Biomethane production by anaerobic digestion of organic waste. Fuel 2013, 103, 1003–1009. [Google Scholar] [CrossRef]
- Bouallagui, H.; Haouari, O.; Touhami, Y.; Ben Cheikh, R.; Marouani, L.; Hamdi, M. Effect of temperature on the performance of an anaerobic tubular reactor treating fruit and vegetable waste. Process Biochem. 2004, 39, 2143–2148. [Google Scholar] [CrossRef]
- Tenca, A.; Schievano, A.; Perazzolo, F.; Adani, F.; Oberti, R. Biohydrogen from thermophilic co-fermentation of swine manure with fruit and vegetable waste: Maximizing stable production without pH control. Bioresour. Technol. 2011, 102, 8582–8588. [Google Scholar] [CrossRef]
- Pascualone, M.J.; Gómez Costa, M.B.; Dalmasso, P.R. Fermentative biohydrogen production from a novel combination of vermicompost as inoculum and mild heat-pretreated fruit and vegetable waste. Biofuel Res. J. 2019, 6, 1046–1053. [Google Scholar] [CrossRef] [Green Version]
- Vendruscolo, F. Starch: A potential substrate for biohydrogen production. Int. J. Energy Res. 2015, 39, 293–302. [Google Scholar] [CrossRef]
- Mona, S.; Kumar, S.S.; Kumar, V.; Parveen, K.; Saini, N.; Deepak, B.; Pugazhendhi, A. Green technology for sustainable biohydrogen production (waste to energy): A review. Sci. Total Environ. 2020, 728, 138481. [Google Scholar] [CrossRef]
- Keskin, T.; Abubackar, H.N.; Yazgin, O.; Gunay, B.; Azbar, N. Effect of percolation frequency on biohydrogen production from fruit and vegetable wastes by dry fermentation. Int. J. Hydrogen Energy 2019, 44, 18767–18775. [Google Scholar] [CrossRef]
- Esteban, J.; Ladero, M. Food waste as a source of value-added chemicals and materials: A biorefinery perspective. Int. J. Food Sci. Technol. 2018, 53, 1095–1108. [Google Scholar] [CrossRef]
- Batista-García, R.A.; del Rayo Sánchez-Carbente, M.; Talia, P.; Jackson, S.A.; O’Leary, N.D.; Dobson, A.D.W.; Folch-Mallol, J.L. From lignocellulosic metagenomes to lignocellulolytic genes: Trends, challenges and future prospects. Biofuels Bioprod. Biorefining 2016, 10, 864–882. [Google Scholar] [CrossRef]
- Chien, P.C.T.; Yoo, H.S.; Dykes, G.A.; Lee, S.M.; Chua, P.; Chien, T.; Yoo, H.S.; Dykes, G.A.; Lee, S.M. Isolation and characterization of cellulose degrading ability in Paenibacillus isolates from landfill leachate. Malays. J. Microbiol. 2015, 11, 185–194. [Google Scholar]
- Feng, H.; Sun, Y.; Zhi, Y.; Mao, L.; Luo, Y.; Wei, X.; Zhou, P. Lignocellulose degradation by the isolate of Streptomyces griseorubens JSD-1. Funct. Integr. Genom. 2015, 15, 163–173. [Google Scholar] [CrossRef]
- Porsch, K.; Wirth, B.; Tóth, E.M.; Schattenberg, F.; Nikolausz, M. Characterization of wheat straw-degrading anaerobic alkali-tolerant mixed cultures from soda lake sediments by molecular and cultivation techniques. Microb. Biotechnol. 2015, 8, 801–814. [Google Scholar] [CrossRef]
- De Melo, R.R.; Tomazetto, G.; Persinoti, G.F.; Sato, H.H.; Ruller, R.; Squina, F.M. Unraveling the cellulolytic and hemicellulolytic potential of two novel Streptomyces strains. Ann. Microbiol. 2018, 68, 677–688. [Google Scholar] [CrossRef]
- Lam, M.Q.; Oates, N.C.; Thevarajoo, S.; Tokiman, L.; Goh, K.M.; McQueen-Mason, S.J.; Bruce, N.C.; Chong, C.S. Genomic analysis of a lignocellulose degrading strain from the underexplored genus Meridianimaribacter. Genomics 2020, 112, 952–960. [Google Scholar] [CrossRef]
- Do, T.H.; Dao, T.K.; Nguyen, K.H.V.; Le, N.G.; Nguyen, T.M.P.; Le, T.L.; Phung, T.N.; van Straalen, N.M.; Roelofs, D.; Truong, N.H. Metagenomic analysis of bacterial community structure and diversity of lignocellulolytic bacteria in Vietnamese native goat rumen. Asian Australas. J. Anim. Sci. 2018, 31, 738–747. [Google Scholar] [CrossRef]
- Ahmed, S.; Rahman, M.S.; Hasan, M.M.; Paul, N.; Sajib, A.A. Microbial degradation of lignocellulosic biomass: Discovery of novel natural lignocellulolytic bacteria. Biotechnologia 2018, 99, 137–146. [Google Scholar] [CrossRef]
- Houfani, A.A.; Větrovský, T.; Navarrete, O.U.; Štursová, M.; Tláskal, V.; Beiko, R.G.; Boucherba, N.; Baldrian, P.; Benallaoua, S.; Jorquera, M.A. Cellulase−Hemicellulase Activities and Bacterial Community Composition of Different Soils from Algerian Ecosystems. Microb. Ecol. 2019, 77, 713–725. [Google Scholar] [CrossRef]
- Cragg, S.M.; Beckham, G.T.; Bruce, N.C.; Bugg, T.D.H.; Distel, D.L.; Dupree, P.; Etxabe, A.G.; Goodell, B.S.; Jellison, J.; McGeehan, J.E.; et al. Lignocellulose degradation mechanisms across the Tree of Life. Curr. Opin. Chem. Biol. 2015, 29, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Al-Masaudi, S.; El Kaoutari, A.; Drula, E.; Al-Mehdar, H.; Redwan, E.M.; Lombard, V.; Henrissat, B. A metagenomics investigation of carbohydrate-active enzymes along the gastrointestinal tract of Saudi sheep. Front. Microbiol. 2017, 8, 666. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Zhang, K.; Liao, H.; Hector, S.B.; Shi, X.; Li, J.; Liu, B.; Xu, T.; Tong, C.; Liu, X.; et al. Genomic and secretomic insight into lignocellulolytic system of an endophytic bacterium Pantoea ananatis Sd-1. Biotechnol. Biofuels 2016, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Dong, D.; Wang, H.; Müller, K.; Qin, Y.; Wang, H.; Wu, W. Metagenomic analysis of microbial consortia enriched from compost: New insights into the role of Actinobacteria in lignocellulose decomposition. Biotechnol. Biofuels 2016, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, G.; Zhou, S.; Luo, R.; Gesang, Z.; Suolang, S. Metagenomic insights into the diversity of carbohydrate-degrading enzymes in the yak fecal microbial community. BMC Microbiol. 2020, 20, 302. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, D.J.; Maruthamuthu, M.; Van Elsas, J.D. Metasecretome analysis of a lignocellulolytic microbial consortium grown on wheat straw, xylan and xylose. Biotechnol. Biofuels 2015, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Cline, L.C.; Zak, D.R. Soil microbial communities are shaped by plant-driven changes in resource availability during secondary succession. Ecology 2015, 96, 3374–3385. [Google Scholar] [CrossRef]
- Güllert, S.; Fischer, M.A.; Turaev, D.; Noebauer, B.; Ilmberger, N.; Wemheuer, B.; Alawi, M.; Rattei, T.; Daniel, R.; Schmitz, R.A.; et al. Deep metagenome and metatranscriptome analyses of microbial communities affiliated with an industrial biogas fermenter, a cow rumen, and elephant feces reveal major differences in carbohydrate hydrolysis strategies. Biotechnol. Biofuels 2016, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Korenblum, E.; Jiménez, D.J.; van Elsas, J.D. Succession of lignocellulolytic bacterial consortia bred anaerobically from lake sediment. Microb. Biotechnol. 2016, 9, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Marynowska, M.; Goux, X.; Sillam-Dussès, D.; Rouland-Lefèvre, C.; Roisin, Y.; Delfosse, P.; Calusinska, M. Optimization of a metatranscriptomic approach to study the lignocellulolytic potential of the higher termite gut microbiome. BMC Genom. 2017, 18, 681. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, D.J.; De Mares, M.C.; Salles, J.F. Temporal expression dynamics of plant biomass-degrading enzymes by a synthetic bacterial consortium growing on sugarcane bagasse. Front. Microbiol. 2018, 9, 299. [Google Scholar] [CrossRef]
- Jiménez, D.J.; Chaves-Moreno, D.; Van Elsas, J.D. Unveiling the metabolic potential of two soil-derived microbial consortia selected on wheat straw. Sci. Rep. 2015, 5, 13845. [Google Scholar] [CrossRef] [Green Version]
- Lemos, L.N.; Pereira, R.V.; Quaggio, R.B.; Martins, L.F.; Moura, L.M.S.; da Silva, A.R.; Antunes, L.P.; da Silva, A.M.; Setubal, J.C. Genome-centric analysis of a thermophilic and cellulolytic bacterial consortium derived from composting. Front. Microbiol. 2017, 8, 644. [Google Scholar] [CrossRef]
- Wilhelm, R.C.; Singh, R.; Eltis, L.D.; Mohn, W.W. Bacterial contributions to delignification and lignocellulose degradation in forest soils with metagenomic and quantitative stable isotope probing. ISME J. 2019, 13, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Raza, M.F.; Zheng, Z.; Zhang, X.; Dong, X.; Zhang, H. Complete genome sequence of Bacillus velezensis ZY-1-1 reveals the genetic basis for its hemicellulosic/cellulosic substrate-inducible xylanase and cellulase activities. 3 Biotech 2018, 8, 465. [Google Scholar] [CrossRef]
- Chen, L.; Gu, W.; Xu, H.Y.; Yang, G.L.; Shan, X.F.; Chen, G.; Kang, Y.H.; Wang, C.F.; Qian, A.D. Comparative genome analysis of Bacillus velezensis reveals a potential for degrading lignocellulosic biomass. 3 Biotech 2018, 8, 253. [Google Scholar] [CrossRef]
- Fabryová, A.; Kostovčík, M.; Díez-Méndez, A.; Jiménez-Gómez, A.; Celador-Lera, L.; Saati-Santamaría, Z.; Sechovcová, H.; Menéndez, E.; Kolařik, M.; García-Fraile, P. On the bright side of a forest pest-the metabolic potential of bark beetles’ bacterial associates. Sci. Total Environ. 2018, 619–620, 9–17. [Google Scholar] [CrossRef]
- Chen, C.; Yang, H.; Xuan, J.; Cui, Q.; Feng, Y. Resonance assignments of a cellulosomal double-dockerin from Clostridium thermocellum. Biomol. NMR Assign. 2019, 13, 97–101. [Google Scholar] [CrossRef]
- Ren, N.; Wang, A.; Cao, G.; Xu, J.; Gao, L. Bioconversion of lignocellulosic biomass to hydrogen: Potential and challenges. Biotechnol. Adv. 2009, 27, 1051–1060. [Google Scholar] [CrossRef]
- Ravindran, R.; Jaiswal, A.K. A comprehensive review on pre-treatment strategy for lignocellulosic food industry waste: Challenges and opportunities. Bioresour. Technol. 2016, 199, 92–102. [Google Scholar] [CrossRef] [Green Version]
- den Haan, R.; van Rensburg, E.; Rose, S.H.; Görgens, J.F.; van Zyl, W.H. Progress and challenges in the engineering of non-cellulolytic microorganisms for consolidated bioprocessing. Curr. Opin. Biotechnol. 2015, 33, 32–38. [Google Scholar] [CrossRef]
- Liu, Z.H.; Le, R.K.; Kosa, M.; Yang, B.; Yuan, J.; Ragauskas, A.J. Identifying and creating pathways to improve biological lignin valorization. Renew. Sustain. Energy Rev. 2019, 105, 349–362. [Google Scholar] [CrossRef]
- Gronenberg, L.S.; Marcheschi, R.J.; Liao, J.C. Next generation biofuel engineering in prokaryotes. Curr. Opin. Chem. Biol. 2013, 17, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wei, Z.; Gao, X.; Wu, J.; Chen, X.; Zhao, Y.; Jia, L.; Wen, D. Reconstruction of core microbes based on producing lignocellulolytic enzymes causing by bacterial inoculation during rice straw composting. Bioresour. Technol. 2020, 315, 123849. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.H.; Kim, D.S. Extracellular cellulolytic enzymes of Bacillus circulans are present as two multipleprotein complexes. Appl. Biochem. Biotechnol. 1993, 42, 83–94. [Google Scholar] [CrossRef]
- Beukes, N.; Pletschke, B.I. Effect of sulfur-containing compounds on Bacillus cellulosome-associated “CMCase” and “Avicelase” activities. FEMS Microbiol. Lett. 2006, 264, 226–231. [Google Scholar] [CrossRef]
- Soundar, S.; Chandra, T.S. Cellulose degradation by a mixed bacterial culture. J. Ind. Microbiol. 1987, 2, 257–265. [Google Scholar] [CrossRef]
- Wushke, S.; Levin, D.B.; Cicek, N.; Sparling, R. Characterization of enriched aerotolerant cellulose-degrading communities for biofuels production using differing selection pressures and inoculum sources. Can. J. Microbiol. 2013, 59, 679–683. [Google Scholar] [CrossRef]
- Kato, S.; Haruta, S.; Cui, Z.J.; Ishii, M.; Igarashi, Y. Effective cellulose degradation by a mixed-culture system composed of a cellulolytic Clostridium and aerobic non-cellulolytic bacteria. FEMS Microbiol. Ecol. 2004, 51, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Komilis, D.P.; Ham, R.K. The effect of lignin and sugars to the aerobic decomposition of solid wastes. In Waste Management; Elsevier Ltd.: Amsterdam, The Netherlands, 2003; Volume 23, pp. 419–423. [Google Scholar]
- Song, N.; Cai, H.Y.; Yan, Z.S.; Jiang, H.L. Cellulose degradation by one mesophilic strain Caulobacter sp. FMC1 under both aerobic and anaerobic conditions. Bioresour. Technol. 2013, 131, 281–287. [Google Scholar] [CrossRef]
- Sawatdeenarunat, C.; Surendra, K.C.; Takara, D.; Oechsner, H.; Khanal, S.K. Anaerobic digestion of lignocellulosic biomass: Challenges and opportunities. Bioresour. Technol. 2015, 178, 178–186. [Google Scholar] [CrossRef]
- Beckham, G.T.; Johnson, C.W.; Karp, E.M.; Salvachúa, D.; Vardon, D.R. Opportunities and challenges in biological lignin valorization. Curr. Opin. Biotechnol. 2016, 42, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Mupondwa, E.; Li, X.; Tabil, L.; Sokhansanj, S.; Adapa, P. Status of Canada’s lignocellulosic ethanol: Part II: Hydrolysis and fermentation technologies. Renew. Sustain. Energy Rev. 2017, 79, 1535–1555. [Google Scholar] [CrossRef]
- Guo, M.; Song, W.; Buhain, J. Bioenergy and biofuels: History, status, and perspective. Renew. Sustain. Energy Rev. 2015, 42, 712–725. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| S. N. | Bacteria Strain | Substrate (Biomass) | Degradation % (Yield) | Degradation Time | Method of Analysis | Product | References |
|---|---|---|---|---|---|---|---|
| 1. | Pantoea ananatis Sd-1 | Pesticide carbaryl, rice straw | 45 | 24 h | Enzyme assays | Reducing sugars | [71] |
| 2. | Firmicutes, Proteobacteria | Wheat, rice, sugarcane, and pea ball-milled straws | - | 84 h | Biolog (MT2) microplate system | Cellulase and xylanase | [23] |
| 3. | Aeromonas hydrophila, Pseudomonas poae, Streptomyces thermoviolaceus, Klebsiella oxytoca, Bacillus amyloliquefaciens | Wheat, rice, sugarcane, and pea straw | 100 | 72 h | Enzyme assays & genomics | Strawase, cellulase | [72] |
| 4. | Streptomyces, Bacillus and Paenibacillus | Saw dust | 40–100 | 7–10 days | Enzyme assays & genomics | Cellulase | [73] |
| 5. | Opitutus terrae, Spirosoma linguale, Solibacter usitatus | Lignite, molasses | - | 5 days | Fermentation | Biogas and organic acid | [14] |
| 6. | phosphate solubilizing (PSB) and potassium solubilizing (KSB) bacteria | Bagasse | - | 7 days 28 days | Randomized blocks design and assays | Carbon Dioxide | [74] |
| 7. | Ochrobactrum sp. | Palm oil mill effluent | 71 | 6 days | Aerobic treatment | CMCase and xylanase | [75] |
| 8. | Pantoea ananatis Sd-1 | Rice straw | 46 | 3 days | Enzyme assays, fenton chemistry | Biofuels | [76] |
| 9. | Bacillus, Streptomyces, Burkholderia | Beech wood | - | 7 days | Enzyme assays. | Xylanase | [77] |
| 10. | Dickeya sp. WS52 | Sweet Pepper and Tomato Stalk | - | 4 days | Enzymatic hydrolysis and genomics | CMCase and pectinase | [78] |
| 11. | Clostridium sp., Petrimonas sp., Methanosarcina sp. and Methanospirillum sp. | Palm oil mill effluent (POME) | - | 18 h | PCR-DGGE and fermentation | Methane | [79] |
| 12. | Streptomyces lividans | Sunflower stalks and rape straw | - | 6 days | Fatty acid profiling | Triacylglycerol | [80] |
| 13. | Bacillus, Enterococcus, Lactococcus, Afipia, Alkaliphilus, Burkholderia, Erwinia, Geobacillus, Ralstonia, Rhodanobacter, Sediminibacterium andStreptococcus. | Rice straw | - | 4 days | Enzyme activity assay | Cellulase | [13] |
| 14. | Firmicutes, Actinobacteria, Proteobacteria and Bacteroidetes | Molasses | - | 124 days | Fermentation | Organic acids and other compounds | [81] |
| 15. | Proteobacteria, Firmicutes and Bacteroidetes | Corn stover | - | 10 days | Assays | Enzymes | [82] |
| 16. | Pseudomonas sp. GO2. | Corn stover | 99.8 | 131 h | Fermentation | Bioflocculant | [83] |
| 17. | Geobacillus sp. strain WSUCF1 | Prairie cordgrass | 100 | 120 h | Single pot bioconversion | Biohydrogen | [84] |
| 18. | Streptomyces sp. MDS | Rice waste biomass, wood straw, local grass powder, sugar cane barboja and sugar cane bagasse | 6 | 6 days | Solid state fermentation | Endoglucanase, exoglucanase, cellobiases, filter pa- perase, amylase, and xylanase | [85] |
| 19. | Bacillus sp. BS-5 | Corn cob | - | 72 h | Enzymes assays | Xylanase, endoglucanase | [55] |
| 20. | Paenibacillus, Streptomyces | Switch grass | - | 10 days | Solid-state and submerged-state cultivation | Biofuel | [86] |
| 21. | Actinobacteria | Olive pomace | - | 6 days | Submerged fermentation | Laccase, xylanase | [87] |
| 22. | Bacillus sp. K1 | Wheat Bran | 44 | 24 h | Lipid extraction and enzyme assays | Lipid, biodiesel | [88] |
| 23. | Bacillus sp. G0 | Miscanthus | 88 | 100 h | Pre-treatment | Xylanase | [89] |
| 24. | Cellulomonas, Pseudomonas, Bacillus, Clostridium, andFibrobacter | Peat | 86 | 42 days | Viability and decomposition tests | NA | [90] |
| 25. | Streptomyces sp., Pseudonocardiaceae, Micromonosporaceae, Saccharothrix | Stipa and Hilaria grass biomass | - | 35 days | Genomics and enzyme assays | Endo- and exo-cellulase, 27 glucosidase, endo- and exo-xylosidase and arabinofuranosidases. | [91] |
| 26. | Paenibacillus sp. | Rice straw, corn straw, soybean straw, and sugarcane bagasse | - | 3 days | Proximate analysis and bacteria growth comparisons | - | [92] |
| 27. | Bacillus cereus RSDa2, Stenotrophomonas maltophilia RSI6, Klebsiella pneumoniae RSI9 | Rice | - | 45 days | Enzyme assays and metagenomics | Compost | [93] |
| 28. | Anaerobic microbial consortium TC-5 | Untreated wheat straw | 45.7 | 9 days | Fermentation | Methane | [47] |
| 29. | Enterobacter xiangfangensis, Serratia rubidaea, Klebsiella pneumoniaeandCitrobacter sp. UWIBGS10. | Sugarcane bagasse | - | 168 h | Hydrolytic production | Glucose | [12] |
| 30. | Clostridium butyricum I5-42 | Starch free fibre from cassava pulp and xylan | - | 24 h | Enzyme assays, fermentation | 1,3-propanediol | [94] |
| 31. | Pantoea ananatis Sd-1 | Lignin & sugar substrates | - | 24 h | Enzyme assays and genomics | Bacterial pyranose 2-oxidase (P2Ox) | [95] |
| 32. | Paenibacillus polymyxa ND25 | CMC, avicel, corn starch, rice straw and sugarcane bagasse | - | 48 h | Enzyme assays | Endoglucanase, exoglucanase and β-glucosidase | [96] |
| 33. | Proteobacteria, Firmicutes, Bacteroidetes Dysgonomona, Sedimentibacter, Comamonas | Energy grass | - | 5–10 days | Shotgun sequencing and enzyme assays | Biogas | [44] |
| 34. | Pseudomonas, ArthrobacterandPseudoxanthomonas | Maize stover | 47 | 30 days | Enzyme assays and genomics | Carboxymethyl cellulase, avicelase, β-glucosidase, endo-β-1,4-xylanase, acetyl esterase, ferulic acid esterase, manganese peroxidase and laccase | [97] |
| 35. | Enhydrobacter | Rice husk | - | 65 days | Blackgram growth and nutrient status analysis | Compost | [98] |
| 36. | Bacillus velezensis 157 | Agro-industrial waste (soybean meal, wheat bran, sugarcane bagasse, wheat straw, rice husk, maize flour and maize straw) | - | 72 h | Solid-state fermentation | Cellulase, xylanase, α-amylase, and pectinase | [99] |
| 37. | Bacteroidetes, Firmicutes, Rikenellaceae, Clostridiaceae, Porphyromonadaceae, Bacteroidaceae, Ruminococcaceae, Firmicutes flavefaciens, Ruminococcus albus | Cow manure | - | 40 days | Biochemical methane potential tests | Biogas/methane | [100] |
| 38. | Ethanoligenens, Tepidimicrobium, Clostridium, Coprococcus, andRuminococcus | Swine Manure, Napier grass | 36.6 | 21 days | Enzyme activity assay | Biogas/Methane | [101] |
| 39. | Prevotella, Bacillus, Thermus, Truepera, and Caldicoprobacter, Thermopolyspora, andPseudoxanthomonas | Weed | - | - | Illumina sequencing | Compost | [102] |
| 40. | Fibrobacter succinogenes S85 | Cellulose | - | 16 h | HPLC, SEM, Activity assays | Cellulases | [103] |
| 41. | Bacteroidetes, Firmicutes, Proteobacteria, Actinobacteria, Fibrobacter, Bacteroides, Clostridium, Prevotella and Ruminococcus | Roughage | - | - | Metagenomics | Cazymes | [104] |
| 42. | Clostridium, Bacteroides, Ruminococcus andMethanosarcina | Rice straw | 83 | 30 days 126 days | Enzyme assays | Methane | [105] |
| 43. | Bacillus sp. TPF-1 | Algae, paper mill wastewater | 48 | 72 h | Enzyme assays | CMCase and xylanase | [106] |
| 44. | Caldicellulosiruptor bescii | Switch grass | 80 | 80 h | Fermentation | Acetate | [107] |
| 45. | Achromobacter sp. (PS1) | Rice-straw, wheat-straw and sugarcane-bagasse | 73.10 and 91.13 | 192 h | One-factor-at-a-time approach | Biosurfactant | [108] |
| 46. | Lactococcus, Serratia, DysgonomonasandEnterococcus | Bamboo shoot particles | - | 6 days | Enzyme assays | Endoglucanase, β-glucosidase, xylanase, exoglucanase, laccase and lignin peroxidase | [109] |
| 47. | Bacteroidetes, Lentisphaerae, Firmicutes and Fibrobacteres | Corn Stover | - | 6 days | Gas chromatography, sequencing | Lignocellulolytic enzyme | [110] |
| 48. | Sphingobacterium sp. ksn-11 | Corn husk, peanut husk, rice bran, sugarcane bagasse, paddy straw, and coffee cherry husk | 60 | 24 h | Sub merged fermentation and optimisation | Cellulase, xylanase, pectinase, mannanase, and laccase | [111] |
| 49. | Bacillus subtillis | Palm oil mill waste | 90 | 14 days | Batch digestion | Methane | [68] |
| 50. | Lactobacillus plantarum RI 11 | Molasses, rice straw, Palm Kernel Cake and soybean | - | 7 days | Enzyme assays | Endoglucanase, exoglucanase, β-glucosidase and mannanase | [112] |
| 51. | Clostridia sp. | Swine manure and corn stover | - | 75 days | Lignocellulolytic activity assays | Methane | [113] |
| 52. | Clostridiales | Wheat straw | 90 | 93 days | Kinetics and batch assays | Methane | [114] |
| 53. | Parabacteroides, Alcaligenes, Lysinibacillus, Sphingobacterium, andClostridium | Rice Straw | 71 | 20 days | Anaerobic digestion | Endo-glucanase | [69] |
| 54. | Alcaligenes faecalis subsp. phenolicus ZY-16 | Citrus peel wastes | 90 | 5 days | Enzyme activity assays | Bioflocculants | [70] |
| 55. | Clostridium thermocellum, C. stercorarium, andThermoanaerobacter thermohydrosulfuricus | Wheat and Cattail Biomass | 80 | 60 days | Consolidated bioprocessing | Ethanol | [115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chukwuma, O.B.; Rafatullah, M.; Tajarudin, H.A.; Ismail, N. A Review on Bacterial Contribution to Lignocellulose Breakdown into Useful Bio-Products. Int. J. Environ. Res. Public Health 2021, 18, 6001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18116001
Chukwuma OB, Rafatullah M, Tajarudin HA, Ismail N. A Review on Bacterial Contribution to Lignocellulose Breakdown into Useful Bio-Products. International Journal of Environmental Research and Public Health. 2021; 18(11):6001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18116001
Chicago/Turabian StyleChukwuma, Ogechukwu Bose, Mohd Rafatullah, Husnul Azan Tajarudin, and Norli Ismail. 2021. "A Review on Bacterial Contribution to Lignocellulose Breakdown into Useful Bio-Products" International Journal of Environmental Research and Public Health 18, no. 11: 6001. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18116001