
Effect of Copper Sulphate and Cadmium Chloride on Non-Human Primate Sperm Function In Vitro


Abstract
:1. Introduction
2. Materials and Methods
2.1. Species Studied
2.2. Sample Collection and Evaluation
2.3. Selection of Motile Spermatozoa
2.4. Exposure of Spermatozoa to Heavy Metals
2.5. Sperm Structural and Functional Testing
2.5.1. Motility Analysis
2.5.2. Viability and Vitality Analysis
Eosin-Nigrosin (E-N)
Hoechst and Propidium Iodide (H&PI)
WST-1 Cell Proliferation Reagent
Evaluation of Acrosome Intactness
Evaluation of Hyperactivation
Statistical Analysis
3. Results
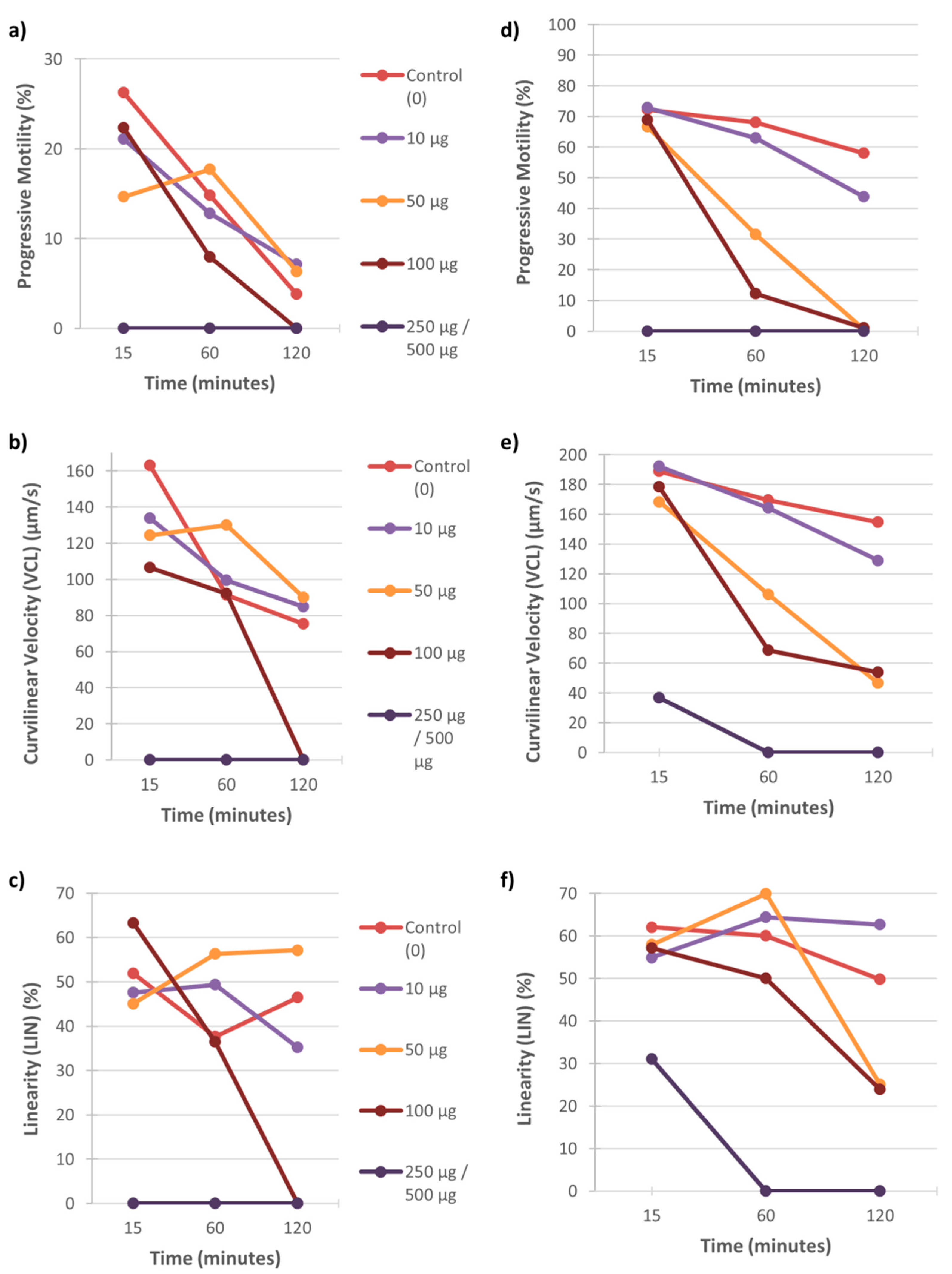
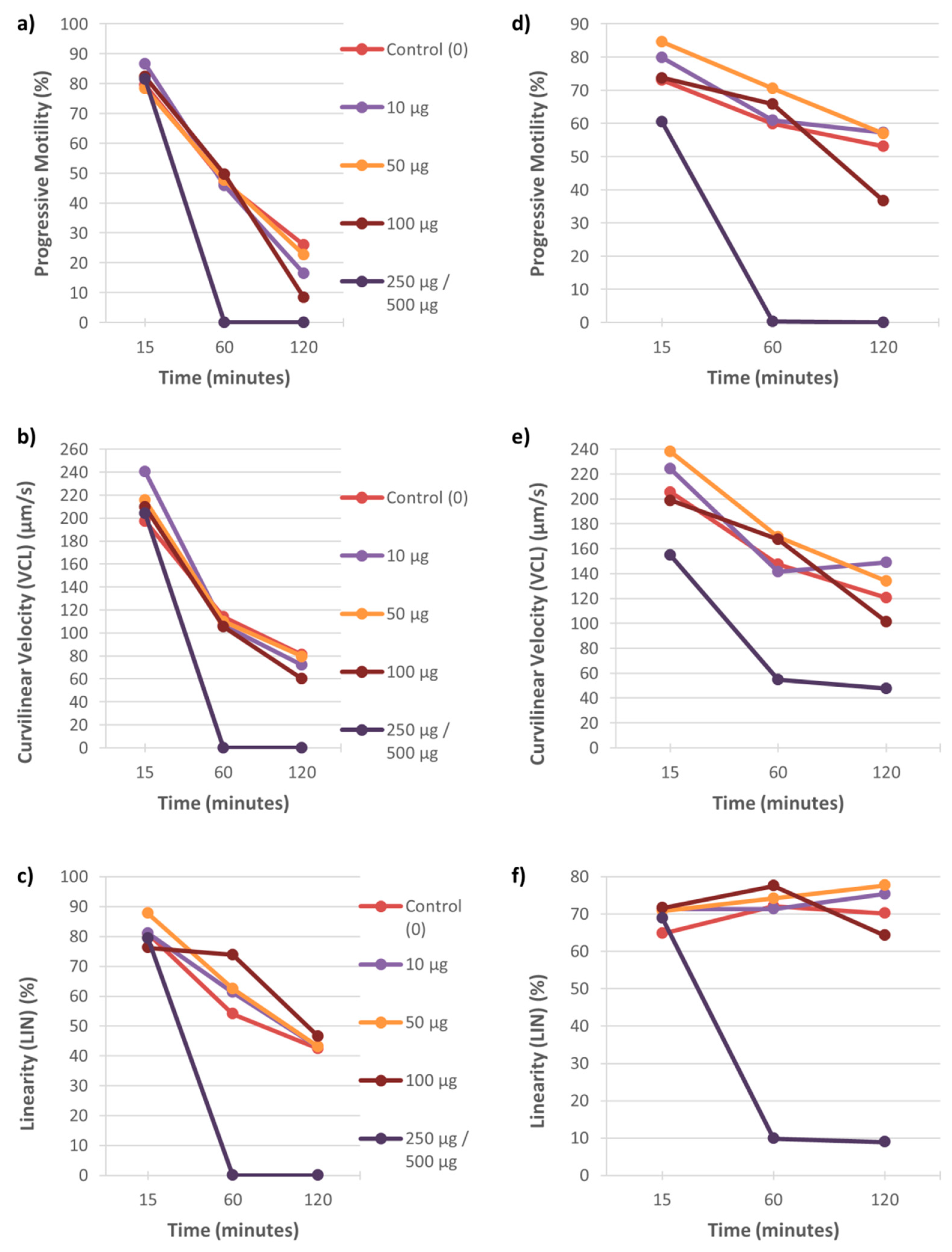
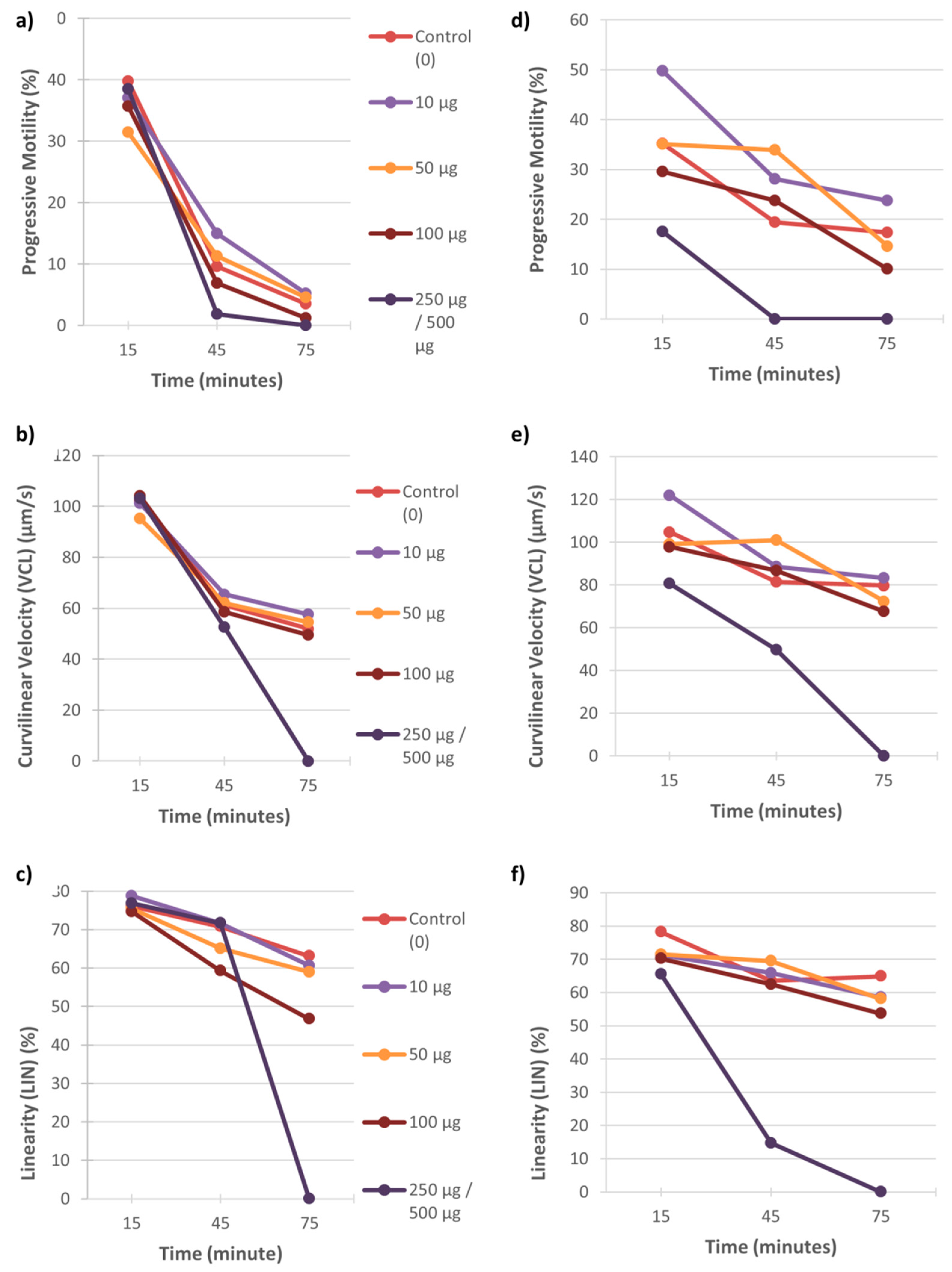
3.1. Motility
3.2. Viability and Vitality
3.2.1. Eosin-Nigrosin (E-N) and Hoechst and Propidium Iodide (H&PI) Staining
3.2.2. Vervet WST-1 Cytotoxicity Assay
3.3. Acrosome Intactness
3.4. Hyperactivation
4. Discussion
4.1. The Effect of Heavy Metals on Sperm Motility
4.2. The Effect of Heavy Metals on Sperm Viability and Vitality
4.3. The Effect of Heavy Metals on Acrosome Intactness
4.4. The Effect of Heavy Metals on Sperm Hyperactivation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mathur, P.P.; D’Cruz, S.C. The effect of environmental contaminants on testicular function. Asian J. Androl. 2011, 13, 585–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurewicz, J.; Radwan, M.; Sobala, W.; Radwan, P.; Bochenek, M.; Hanke, W. Effects of occupational exposure—Is there a link between exposure based on an occupational questionnaire and semen quality. Syst. Biol. Reprod. Med. 2014, 60, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D. Lifestyle causes of male infertility. Arab J. Urol. 2018, 16, 10–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puluputturi, S.R.; Dayapulae, J.R. Metals: Male reproductive function. Int. J. Pharm. Sci. Rev. Res. 2012, 16, 56–60. [Google Scholar]
- Maartens, P.J.; Aboua, Y.G.; DuPlessis, S.S. Role of environmental factors and gonadotoxin exposure in unexplained male infertility. In Unexplained Infertility: Pathophysiology, Evaluation and Treatment; Springer: New York, NY, USA, 2015. [Google Scholar] [CrossRef]
- Mumford, S.L.; Kim, S.; Chen, Z.; Gore-Langton, R.E.; Barr, D.B.; Louis, G.M.B. Persistent organic pollutants and semen quality: The life study. Chemosphere 2015, 135, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Alharbi, O.M.L.; Arsh, A.; Khattab, R.A.; Ali, I. Health and environmental effects of persistent organic pollutants. J. Mol. Liq. 2018, 263, 442–453. [Google Scholar] [CrossRef]
- Ji, H.; Miao, M.; Liang, H.; Shi, H.; Ruan, D.; Li, Y.; Wang, J.; Yuan, W. Exposure of environmental bisphenol A in relation to routine sperm parameters and sperm movement characteristics among fertile men. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Anifandis, G.; Amiridis, G.; Dafopoulos, K.; Daponte, A.; Dovolou, E.; Gavriil, E.; Gorgogietas, V.; Kachpani, E.; Mamuris, Z.; Messini, C.I.; et al. The in vitro impact of the herbicide roundup on human sperm motility and sperm mitochondria. Toxics 2017, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Anifandis, G.; Katsanaki, K.; Lagodonti, G.; Messini, C.; Simopoulou, M.; Dafopoulos, K.; Daponte, A. The effect of glyphosate on human sperm motility and sperm DNA fragmentation. Int. J. Environ. Res. Public Health 2018, 15, 1117. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.; Gomes, W.; van Denmark, N. The Testis; Academic Press: Cambridge, MA, USA, 1970; Volume 3. [Google Scholar]
- Monseny, J.M.P.; Arrondo, J.L.A. Practica Andrologia, Scientific; Masson-Salvat: Barcelona, Spain, 1993. [Google Scholar]
- Bustos-Obregon, E. Adverse effects of exposure to agro pesticides on male reproduction. APMIS 2001, 109, 233–242. [Google Scholar] [CrossRef]
- Pizent, A.; Tariba, B.; Zivkovic, T. Reproductive toxicity of metals in men. Arch. Ind. Hyg. Toxicol. 2012, 63, 35. [Google Scholar] [CrossRef]
- Rana, S.V.S. Perspectives in endocrine toxicity of heavy metals—A review. Biol. Trace Elem. Res. 2014, 160, 1–14. [Google Scholar] [CrossRef]
- Apostoli, P.; Kiss, P.; Porru, S.; Bonde, J.P.; Vanhoorne, M. Male reproductive toxicity of lead in animals and humans. Occup. Environ. Med. 1998, 55, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Benoff, S.; Jacob, A.; Hurley, I.R. Male infertility and environmental exposure to lead and cadmium. Hum. Reprod. Update 2000, 6, 107–121. [Google Scholar] [CrossRef]
- Apostoli, P.; Telišman, S.; Sager, P.R. Reproductive and developmental toxicity of metals. In Handbook on the Toxicology of Metals; Elsevier: Amsterdam, The Netherlands, 2007; pp. 213–249. [Google Scholar] [CrossRef]
- Wirth, J.J.; Mijal, R.S. Adverse effects of low level heavy metal exposure on male reproductive function. Syst. Biol. Reprod. Med. 2010, 56, 147–167. [Google Scholar] [CrossRef] [Green Version]
- Apostoli, P.; Catalani, S. Metal ions affecting reproduction and development. Metal Ions Life Sci. 2011, 8, 263–303. [Google Scholar] [CrossRef]
- Carette, D.; Perrard, M.-H.; Prisant, N.; Gilleron, J.; Pointis, G.; Segretain, D.; Durand, P. Hexavalent chromium at low concentration alters Sertoli cell barrier and connexin 43 gap junction but not claudin-11 and N-cadherin in the rat seminiferous tubule culture model. Toxicol. Appl. Pharmacol. 2013, 268, 27–36. [Google Scholar] [CrossRef]
- Mima, M.; Greenwald, D.; Ohlander, S. Environmental toxins and male fertility. Curr. Urol. Rep. 2018, 19, 393–400. [Google Scholar] [CrossRef]
- Adejuwon, C.A.; Ilesanmi, A.O.; Ode, E.O.; Akinlade, K.S. Biophysical and biochemical analysis of semen in infertile Nigerian males. Afr. J. Med. Med. Sci. 1996, 25, 217–219. [Google Scholar]
- Yuyan, L.; Junqing, W.; Wei, Y.; Weijin, Z.; Ersheng, G. Are serum zinc and copper levels related to semen quality? Fertil. Steril. 2008, 89, 1008–1011. [Google Scholar] [CrossRef]
- Kumar, N.; Singh, A. Trends of male factor infertility, an important cause of infertility: A review of literature. J. Hum. Reprod. Sci. 2015, 8, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Barratt, C.L.R.; Björndahl, L.; de Jonge, C.J.; Lamb, D.J.; Martini, F.O.; McLachlan, R.; Oates, R.D.; van der Poel, S.; John, B.S.; Sigman, M.; et al. The diagnosis of male infertility: An analysis of the evidence to support the development of global WHO guidance-challenges and future research opportunities. Hum. Reprod. Update 2017, 23, 660–680. [Google Scholar] [CrossRef]
- Oehninger, S.; Franken, D.R.; Ombelet, W. Sperm functional tests. Fertil. Steril. 2014, 102, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Kızılay, F.; Altay, B. Klinik uygulamadaki sperm fonksiyon testleri. Turk. J. Urol. 2017, 43, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Niżański, W.; Partyka, A.; Prochowska, S. Evaluation of spermatozoal function—Useful tools or just science. Reprod. Domest. Anim. 2016, 51, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Abu, D.A.H.; Franken, D.R.; Hoffman, B.; Henkel, R. Sequential analysis of sperm functional aspects involved in fertilisation: A pilot study. Andrologia 2012, 44, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Van der Horst, G.; du Plessis, S.S. Not just the marriage of figaro: But the marriage of WHO/ESHRE semen analysis criteria with sperm functionality. Postępy Andrologii Online 2017, 4, 6–21. [Google Scholar]
- Husak, V. Copper and copper-containing pesticides: Metabolism, toxicity and oxidative stress. J. Vasyl Stefanyk Precarpathian Natl. Univ. 2015, 2, 38–50. [Google Scholar] [CrossRef]
- Hardneck, F.; Israel, G.; Pool, E.; Maree, L. Quantitative assessment of heavy metal effects on sperm function using computer-aided sperm analysis and cytotoxicity assays. Andrologia 2018, 50, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Shibahara, H.; Obara, H.; Ayustawati; Hirano, Y.; Suzuki, T.; Ohno, A.; Takamizawa, S.; Suzuki, M. Prediction of pregnancy by intrauterine insemination using CASA estimates and strict criteria in patients with male factor infertility. Int. J. Androl. 2004, 27, 63–68. [Google Scholar] [CrossRef]
- Nagy, Á.; Polichronopoulos, T.; Gáspárdy, A.; Solti, L.; Cseh, S. Correlation between bull fertility and sperm cell velocity parameters generated by computer-assisted semen analysis. Acta Vet. Hung. 2015, 63, 370–381. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, H.; Andrabi, S.M.H.; Anwar, M.; Jahan, S. Use of post-thaw semen quality parameters to predict fertility of water buffalo (Bubalus bubalis) bull during peak breeding season. Andrologia 2017, 49. [Google Scholar] [CrossRef]
- Marchiani, S.; Tamburrino, L.; Farnetani, G.; Muratori, M.; Vignozzi, L.; Baldi, E. Acute effects on human sperm exposed in vitro to cadmium chloride and diisobutyl phthalate. Reproduction 2019, 158, 281–290. [Google Scholar] [CrossRef]
- Dancet, E.A.F.; Spiessens, C.; Vangenechten, R.; Billiet, J.; de Tavernier, J.; Welkenhuysen, M.; D’Hooghe, T.M. Acceptability of preclinical research on nonhuman primates in reproductive medicine: The patient perspective. Reprod. Sci. 2011, 18, 70–78. [Google Scholar] [CrossRef]
- Van der Horst, G. Reproduction: Male. In The Laboratory Primate; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar] [CrossRef]
- Chellman, G.J.; Bussiere, J.L.; Makori, N.; Martin, P.L.; Ooshima, Y.; Weinbauer, G.F. Developmental and reproductive toxicology studies in nonhuman primates. Birth Defects Res. Part B 2009, 86, 446–462. [Google Scholar] [CrossRef]
- Faqi, A.S. A critical evaluation of developmental and reproductive toxicology in nonhuman primates. Syst. Biol. Reprod. Med. 2012, 58, 23–32. [Google Scholar] [CrossRef]
- Subramanyam, M.; Rinaldi, N.; Mertsching, E.; Hutto, D. Selection of relevant species. In Preclinical Safety Evaluation of Biopharmaceuticals: A Science-Based Approach to Facilitating Clinical Trials; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Prag, F.C. Evaluation of Standard and Development of New Sperm Function Tests in Selected Primate Species. Master’s Thesis, University of the Western Cape, Cape Town, South Africa, 2017. [Google Scholar]
- Mdhluli, M.C.; Seier, J.V.; van der Horst, G. The male vervet monkey: Sperm characteristics and use in reproductive research. Gynecol. Obstet. Investig. 2004, 57, 17–18. [Google Scholar]
- De Villiers, C. A comparison between the semen and sperm parameters from the captive-bred vervet monkey (Chlorocebus aethiops) and rhesus monkey (Macaca mulatta). J. Med. Primatol. 2018, 47, 211–216. [Google Scholar] [CrossRef]
- Bornman, M.S.; van Vuuren, M.; Meltzer, D.G.; van der Merwe, C.A.; Rensburg, S.J.v. Quality of semen obtained by electroejaculation from chacma baboons (Papio ursinus). J. Med. Primatol. 1988, 17, 57–61. [Google Scholar] [CrossRef]
- Alberts, S.C.; Altmann, J. Reproductive behavior in wild baboons. Gynecol. Obstet. Investig. 2004, 57, 10–13. [Google Scholar]
- Sparman, M.L.; Ramsey, C.M.; Thomas, C.M.; Mitalipov, S.M.; Fanton, J.W.; Maginnis, G.M.; Stouffer, R.L.; Wolf, D.P. Evaluation of the vervet (Clorocebus aethiops) as a model for the assisted reproductive technologies. Am. J. Primatol. 2007, 69, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Seier, J.V.; Fincham, J.E.; Menkveld, R.; Venter, F.S. Semen characteristics of vervet monkeys. Lab. Anim. 1989, 23, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Cseh, S.; Chan, P.J.; Corselli, J.; Bailey, L.L. Electroejaculated baboon (Papio anubis) sperm requires a higher dosage of pentoxifylline to enhance motility. J. Assist. Reprod. Genet. 2000, 17, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, D. Practical Laboratory Andrology; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Mortimer, S.T. A critical review of the physiological importance and analysis of sperm movement in mammals. Hum. Reprod. Update 1997, 3, 403–439. [Google Scholar] [CrossRef] [Green Version]
- Maree, L.; van der Horst, G. Quantification and identification of sperm subpopulations using computer-aided sperm analysis and species-specific cut-off values for swimming speed. Biotech. Histochem. 2013, 88, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Roche Diagnostics. Cell Proliferation Reagent WST-1; Roche Diagnostics: Mannheim, Germany, 2018. [Google Scholar]
- Berridge, M.V.; Tan, A.S. Trans-plasma membrane electron transport: A cellular assay for NADH- and NADPH-oxidase based on extracellular, superoxide-mediated reduction of the sulfonated tetrazolium salt WST-1. Protoplasma 1998, 205, 74–82. [Google Scholar] [CrossRef]
- Ntanjana, N. Hyperactivation in Human Semen and Sperm Subpopulations by Selected Calcium Modulators. Master’s Thesis, University of the Western Cape, Cape Town, South Africa, 2015. [Google Scholar]
- Boshoff, N.H.; Lambrechts, H.; Maree, L.; Cloete, S.W.P.; van der Horst, G. A novel flush technique to simulate natural dispersal of spermatozoa in the female reproductive tract and expedite motility assessment of fresh ejaculated merino (Ovis aries) sperm. S. Afr. J. Anim. Sci. 2018, 48, 469. [Google Scholar] [CrossRef]
- World Health Organization. Laboratory Manual for the Examination and Processing of Human Semen; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar] [CrossRef] [Green Version]
- Baumber, J.; Meyers, S.A. Changes in membrane lipid order with capacitation in rhesus macaque (Macaca mulatta) spermatozoa. J. Androl. 2006, 27, 578–587. [Google Scholar] [CrossRef] [Green Version]
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef]
- Mínguez-Alarcón, L.; Williams, P.L.; Chiu, Y.H.; Gaskins, A.J.; Nassan, F.L.; Dadd, R.; Petrozza, J.; Hauser, R.; Chavarro, J.E. Secular trends in semen parameters among men attending a fertility center between 2000 and 2017: Identifying potential predictors. Environ. Int. 2018, 121, 1297–1303. [Google Scholar] [CrossRef]
- Van der Horst, G.; Maree, L.; du Plessis, S.S. Current perspectives of CASA applications in diverse mammalian spermatozoa. Reprod. Fertil. Dev. 2018, 30, 875–888. [Google Scholar] [CrossRef] [Green Version]
- Slanina, T.; Miškeje, M.; Petrovičová, I.; Lukáč, N.; Massányi, P. Changes in turkey spermatozoza motility parameters after addition of copper sulphate in vitro. J. Microbiol. Biotechnol. Food Sci. 2015, 4, 98–100. [Google Scholar] [CrossRef] [Green Version]
- Hess, R.A. Effects of environmental toxicants on the efferent ducts, epididymis and fertility. J. Reprod. Fertil. Suppl. 1998, 53, 247–259. [Google Scholar]
- Waliszewski, S.M.; Aguirre, A.A.; Infanzón, R.M.; Siliceo, J. Carry-over of persistent organochlorine pesticides through placenta to fetus. Salud Pública de México 2000, 42, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Ojemaye, C.Y.; Petrik, L. Occurrences, levels and risk assessment studies of emerging pollutants (pharmaceuticals, perfluoroalkyl and endocrine disrupting compounds) in fish samples from Kalk bay harbour, South Africa. Environ. Pollut. 2019, 252, 562–572. [Google Scholar] [CrossRef]
- Olivari, F.A.; Hernández, P.P.; Allende, M.L. Acute copper exposure induces oxidative stress and cell death in lateral line hair cells of zebrafish larvae. Brain Res. 2008, 1244, 1–12. [Google Scholar] [CrossRef]
- Tvrdá, E.; Lukáč, N.; Schneidgenová, M.; Lukáčová, J.; Szabó, C.; Goc, Z.; Greń, A.; Massányi, P. Impact of seminal chemical elements on the oxidative balance in bovine seminal plasma and spermatozoa. J. Vet. Med. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Viarengo, A.; Pertica, M.; Mancinelli, G.; Zanicchi, G.; Orunesu, M. Rapid induction of copper-binding proteins in the gills of metal exposed mussels. Comp. Biochem. Physiol. Part C 1980, 67, 215–218. [Google Scholar] [CrossRef]
- Wimalasena, D.S.; Wiese, T.J.; Wimalasena, K. Copper ions disrupt dopamine metabolism via inhibition of V-H+-ATPase: A possible contributing factor to neurotoxicity. J. Neurochem. 2007, 101, 313–326. [Google Scholar] [CrossRef]
- Earnshaw, M.J.; Wilson, S.; Akberali, H.B.; Butler, R.D.; Marriott, K.R.M. The action of heavy metals on the gametes of the marine mussel, Mytilus edulis (L.)-III. The effect of applied copper and zinc on sperm motilityin relation to ultrastructural damage and intracellular metal localisation. Mar. Environ. Res. 1986, 20, 261–278. [Google Scholar] [CrossRef]
- Krumschnabel, G.; Manzl, C.; Berger, C.; Hofer, B. Oxidative stress, mitochondrial permeability transition, and cell death in cu-exposed trout hepatocytes. Toxicol. Appl. Pharmacol. 2005, 209, 62–73. [Google Scholar] [CrossRef]
- Oliveira, H.; Spanò, M.; Santos, C.; Pereira, M.D.L. Adverse effects of cadmium exposure on mouse sperm. Reprod. Toxicol. 2009, 28, 550–555. [Google Scholar] [CrossRef]
- Kanous, K.S.; Casey, C.; Lindemann, C.B. Inhibition of microtubule sliding by Ni2+ and Cd2+: Evidence for a differential response of certain microtubule pairs within the bovine sperm axoneme. Cell Motil. Cytoskelet. 1993, 26, 66–76. [Google Scholar] [CrossRef]
- Lindemann, C.B.; Gardner, T.K.; Westbrook, E.; Kanous, K.S. The calcium-induced curvature reversal of rat sperm is potentiated by CAMP and inhibited by anti-calmodulin. Cell Motil. Cytoskelet. 1991, 20, 316–324. [Google Scholar] [CrossRef]
- Schlingmann, K.; Michaut, M.A.; Mcelwee, J.L.; Wolff, C.A.; Travis, A.J.; Turner, R.M. Calmodulin and CaMKII in the sperm principal piece: Evidence for a motility-related calcium/calmodulin pathway. J. Androl. 2007, 28, 706–716. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, H.; Saravia, F.; Wallgren, M.; Roca, J.; Peña, F.J. Influence of seminal plasma on the kinematics of boar spermatozoa during freezing. Theriogenology 2008, 70, 1242–1250. [Google Scholar] [CrossRef]
- Tsakmakidis, I.A.; Lymberopoulos, A.G.; Khalifa, T.A.A. Relationship between sperm quality traits and field-fertility of porcine semen. J. Vet. Sci. 2010, 11, 151–154. [Google Scholar] [CrossRef] [Green Version]
- Amann, R.P.; Waberski, D. Computer-assisted sperm analysis (CASA): Capabilities and potential developments. Theriogenology 2014, 81, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Kwolek-Mirek, M.; Zadrag-Tecza, R. Comparison of methods used for assessing the viability and vitality of yeast cells. FEMS Yeast Res. 2014, 14, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Tabassomi, M.; Alavi-Shoushtari, S.M. Effects of in vitro copper sulphate supplementation on the ejaculated sperm characteristics in water buffaloes (Bubalus bubalis). Vet. Res. Forum 2013, 4, 31–36. [Google Scholar] [PubMed]
- Zhao, X.; Cheng, Z.; Zhu, Y.; Li, S.; Zhang, L.; Luo, Y. Effects of paternal cadmium exposure on the sperm quality of male rats and the neurobehavioral system of their offspring. Exp. Ther. Med. 2015, 10, 2356–2360. [Google Scholar] [CrossRef] [Green Version]
- Maya-Soriano, M.J.; Abello, M.T.; Fernandez-Bellon, H.; Martin, M.; Vidal, J.; Salvador, C.; Lopez-Bejar, M. Reproductive assessment and preliminary evaluation of assisted reproductive technologies in drills (Mandrillus leucophaeus). J. Zoo Aquar. Res. 2015, 3, 116–122. [Google Scholar]
- Talwar, G.P.; Naz, R.K.; Das, C.; Das, R.P. A practicable immunological approach to block spermatogenesis without loss of androgens. Proc. Natl. Acad. Sci. USA 1979, 76, 5882–5885. [Google Scholar] [CrossRef] [Green Version]
- Cardaci, P.J. Apical Testis Structure and the Effects of Cadmium Treatment on Spermatogenesis in Drosophila. Ph.D. Thesis, Seton Hall University, South Orange, NJ, USA, 2016. [Google Scholar]
- Department of Water Affairs and Forestry. South African Water Quality Guidelines, 2nd ed.; Domestic Water Use; Copper; Department of Water Affairs and Forestry: Pretoria, South Africa, 1996; Volume 1, pp. 57–59.
- Aitken, R.J.; Ryan, A.L.; Curry, B.J.; Baker, M.A. Multiple forms of redox activity in populations of human spermatozoa. Mol. Hum. Reprod. 2003, 9, 645–661. [Google Scholar] [CrossRef] [Green Version]
- Knazicka, Z.; Tvrda, E.; Bardos, L.; Lukac, N. Dose- and time-dependent effect of copper ions on the viability of bull spermatozoa in different media. J. Environ. Sci. Health Part A 2012, 47, 1294–1300. [Google Scholar] [CrossRef] [PubMed]
- Dawson, E.B.; Ritter, S.; Harris, W.A.; Evans, D.R.; Powell, L.C. Comparison of sperm viability with seminal plasma metal levels. Biol. Trace Elem. Res. 1998, 64, 215–219. [Google Scholar] [CrossRef]
- Chia, S.E.; Xu, B.; Ong, C.N.; Tsakok, F.M.H.; Lee, S.T. Effect of cadmium and cigarette smoking on human semen quality. Int. J. Fertil. Menopausal Stud. 1994, 39, 292–298. [Google Scholar]
- Liu, R.; Gao, J.C.; Zhang, H.G.; Wang, R.X.; Zhang, Z.H.; Liu, X.Y. Seminal plasma zinc level may be associated with the effect of cigarette smoking on sperm parameters. J. Int. Med. Res. 2010, 38, 923–928. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Sharma, S. Evaluation of toxic effect of cadmium on sperm count, sperm motility and sperm abnormality in albino mice. Int. J. Adv. Res. 2015, 3, 335–343. [Google Scholar]
- University of Utah. Male Fertility Tests and Procedure. Available online: https://healthcare.utah.edu/fertility/treatments/diagnostic-testing/all-tests.php#long (accessed on 29 June 2020).
- Guraya, S.S. Cellular and molecular biology of capacitation and acrosome reaction in spermatozoa. Int. Rev. Cytol. 2000, 199, 1–64. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Bulla, J.; Massanyi, P.; Choudhury, M.D. Copper affects spermatozoa motility, morphology and cell membrane integrity in rabbits in vitro. J. Biotechnol. 2010, 150, 451. [Google Scholar] [CrossRef]
- Misro, M.M.; Chaki, S.P.; Chandra, M.; Maheshwari, A.; Nandan, D. Release of copper from CuT 380a co-incubated with human semen and its effect on sperm function in vitro. Indian J. Physiol. Pharmacol. 2008, 52, 267–273. [Google Scholar] [PubMed]
- Mortimer, S.T.; Swan, M.A.; Mortimer, D. Effect of seminal plasma on capacitation and hyperactivation in human spermatozoa. Hum. Reprod. 1998, 13, 2139–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, D.; Nandi, P.; Varghese, A.C.; Gutgutia, R.; Banerjee, S.; Bhattacharyya, A.K. The in vitro effect of benzo[a]pyrene on human sperm hyperactivation and acrosome reaction. Fertil. Steril. 2010, 94, 595–598. [Google Scholar] [CrossRef]
- Hung, P.H.; Baumber, J.; Meyers, S.A.; VandeVoort, C.A. Effects of environmental tobacco smoke in vitro on rhesus monkey sperm function. Reprod. Toxicol. 2007, 23, 499–506. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Time | CuSO4 | Live Spermatozoa | CdCl2 | Live Spermatozoa | ||
|---|---|---|---|---|---|---|
| (min) | (µg/mL) | (%) | (µg/mL) | (%) | ||
| Vervet # | Baboon § | Vervet # | Baboon § | |||
| 15 | 0 | 54.0 ± 37.1 | 62.9 ± 11.4 | 0 | 53.2 ± 30.7 | 45.0 ± 39.5 a |
| 10 | 54.5 ± 33.9 | 46.8 ± 13.5 | 10 | 49.3 ± 29.8 | 43.6 ± 10.3 a | |
| 50 | 48.6 ± 37.5 | 57.9 ± 8.8 | 50 | 57.5 ± 24.5 | 39.6 ± 11.3 a | |
| 100 | 33.2 ± 33.3 | 34.3 ± 14.8 | 100 | 41.4 ± 29.4 | 36.1 ± 15.3 a | |
| 250 | 19.3 ± 29.8 | 51.5 ± 18.4 | 500 | 46.6 ± 38.3 | 12.5 ± 12.0 b | |
| 90/120 * | 0 | 26.1 ± 28.3 a | 51.5 ± 20.6 | 0 | 50.9 ± 36.5 a | 33.8 ± 31.3 |
| 10 | 30.0 ± 40.4 a | 55.7 ± 15.1 | 10 | 40.9 ± 28.8 a | 27.1 ± 13.8 | |
| 50 | 22.9 ± 39.6 a | 40.4 ± 7.5 | 50 | 38.4 ± 38.8 a | 28.7 ± 25.1 | |
| 100 | 0.0 ± 0.0 b | 31.9 ± 9.7 | 100 | 37.6 ± 36.1 a | 19.9 ± 24.1 | |
| 250 | 0.0 ± 0.0 b | 55.7 ± 17.1 | 500 | 0.2 ± 0.5 b | 9.8 ± 9.1 | |
| Time | CuSO4 | Intact Acrosome | CdCl2 | Intact Acrosome | ||
|---|---|---|---|---|---|---|
| (min) | (µg/mL) | (%) | (µg/mL) | (%) | ||
| Vervet | Baboon | Vervet | Baboon | |||
| 15 | 0 | 26.2 ± 22.9 | 74.3 ± 18.1 a | 0 | 43.5 ± 35.0 | 94.1 ± 7.2 |
| 10 | 35.6 ± 24.8 | 77.9 ± 11.2 a | 10 | 54.1 ± 40.8 | 91.1 ± 10.4 | |
| 50 | 21.1 ± 26.9 | 60.2 ± 31.9 a | 50 | 52.1 ± 39.4 | 90.6 ± 12.5 | |
| 100 | 28.2 ± 33.3 | 55.6 ± 31.2 a | 100 | 47.2 ± 36.5 | 92.6 ± 8.8 | |
| 250 | 24.4 ± 28.3 | 19.2 ± 29.8 b | 500 | 48.8 ± 37.6 | 84.4 ± 7.5 | |
| 90/120 * | 0 | 34.4 ± 20.4 | 71.7 ± 13.2 a | 0 | 37.8 ± 36.1 | 91.5 ± 4.7 |
| 10 | 34.2 ± 26.6 | 75.0 ± 12.8 a | 10 | 47.5 ± 35.5 | 90.9 ± 8.9 | |
| 50 | 27.8 ± 30.8 | 69.7 ± 17.5 b | 50 | 41.1 ± 33.1 | 90.2 ± 5.6 | |
| 100 | 28.2 ± 24.7 | 45.4 ± 23.0 c | 100 | 30.9 ± 35.8 | 91.6 ± 7.4 | |
| 250 | 25.0 ± 35.0 | 17.3 ± 27.0 d | 500 | 28.9 ± 23.2 | 85.2 ± 5.6 | |
| Time | CuSO4 | Caffeine | Hyperactivation | CdCl2 | Caffeine | Hyperactivation | ||
|---|---|---|---|---|---|---|---|---|
| (min) | (µg/mL) | (mM) | (%) | (µg/mL) | (mM) | (%) | ||
| Vervet | Rhesus | Vervet | Rhesus | |||||
| 0 | 0 | 10.1 ± 10.6 | 63.5 | 0 | 0 | 6.0 ± 1.2 a | 41.1 | |
| 15 (V) | 0 | 5 | 12.0 ± 3.2 | 70.4 | 0 | 5 | 14.1 ± 12.8 b | 71.2 |
| 10 (R) | 100 | 0 | 4.2 ± 3.6 | 71.9 | 100 | 0 | 0.4 ± 0.8 c | 37.8 |
| 100 | 5 | 16.8 ± 1.9 | 71.1 | 100 | 5 | 12.8 ± 18.5 d | 61.5 | |
| 0 | 0 | 7.6 ± 7.7 | 59.1 | 0 | 0 | 7.7 ± 6.6 a | 17.7 | |
| 20 (V) | 0 | 5 | 7.0 ± 7.1 | 68.8 | 0 | 5 | 11.2 ± 16.3 a | 82.3 |
| 35 (R) | 100 | 0 | 3.4 ± 5.1 | 35.6 | 100 | 0 | 0.8 ± 1.7 b | 26.4 |
| 100 | 5 | 13.2 ± 12.4 | 64.1 | 100 | 5 | 9.2 ± 15.7 c | 66.1 | |
| 0 | 0 | 5.6 ± 6.3 a | 65.6 | 0 | 0 | 3.9 ± 5.0 a | 16.3 | |
| 40 (V) | 0 | 5 | 6.7 ± 8.5 a | 78.8 | 0 | 5 | 6.8 ± 13.5 a | 66.5 |
| 50 (R) | 100 | 0 | 1.1 ± 1.4 b | 41.1 | 100 | 0 | 0.4 ± 0.9 b | 15.0 |
| 100 | 5 | 5.6 ± 5.1 c | 71.7 | 100 | 5 | 7.2 ± 10.7 c | 54.0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardneck, F.; de Villiers, C.; Maree, L. Effect of Copper Sulphate and Cadmium Chloride on Non-Human Primate Sperm Function In Vitro. Int. J. Environ. Res. Public Health 2021, 18, 6200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18126200
Hardneck F, de Villiers C, Maree L. Effect of Copper Sulphate and Cadmium Chloride on Non-Human Primate Sperm Function In Vitro. International Journal of Environmental Research and Public Health. 2021; 18(12):6200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18126200
Chicago/Turabian StyleHardneck, Farren, Charon de Villiers, and Liana Maree. 2021. "Effect of Copper Sulphate and Cadmium Chloride on Non-Human Primate Sperm Function In Vitro" International Journal of Environmental Research and Public Health 18, no. 12: 6200. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18126200