3.1. Analysis of Variables at Baseline and Genetic Correlates
Baseline variables of postmenopausal women having endothelial dysfunction categorized according to osteoporosis status are summarized in
Table 1. No evidence of statistically significant differences between the two groups for age, YSM, DBP, LDL, HDL, and TC (
p > 0.05) was observed. Values of BMI, SBP and TG were considerable higher in women having osteoporosis than women without osteoporosis and these differences were statistically significant (
p < 0.001) between both the groups. Albeit, all the postmenopausal women participated in the present study had confirmed endothelial dysfunction (RHI < 1.67), nonetheless, its effect was considerably enlarged in women having osteoporosis and differences between the groups were statistically dissimilar (
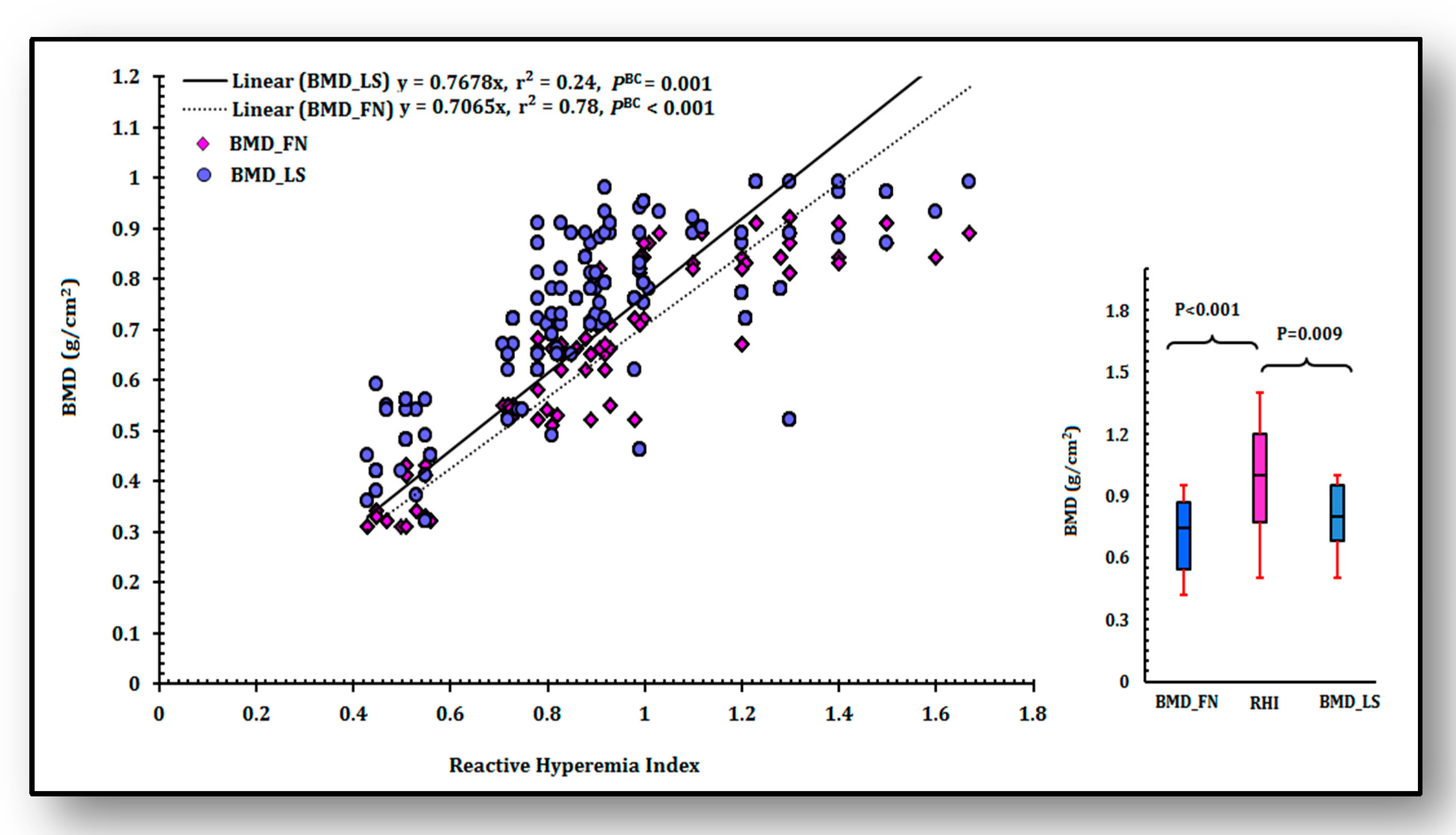
p < 0.001). Similarly, it is evident that adjusted values of BMD at femoral neck and lumbar spine were markedly low in women having osteoporosis than women without it (
p < 0.00).
Genotype frequencies of all the SNPs of three genes (eNOS, ACE and VEGFA) were within range (p > 0.05) of Hardy-Weinberg equilibrium. Minor allele frequencies (MAF) of rs1800780, rs3918181 and rs1808593 of eNOS gene were similar between both groups (p > 0.05), however, for rs2070744, rs1799983, and rs891512, MAF differed significantly (p < 0.05) between osteoporotic and non-osteoporotic women. MAF of three SNPs of ACE gene i.e., rs1800764, rs1799752 and rs4343 were observed to be significantly dissimilar (p < 0.05), however MAF of rs4459609 was similar (p = 7.86) between women having osteoporosis and women without it. MAFs of SNP rs833061 within VEGFA gene were detected to be non-significant (p = 0.793) between both the groups of women, whereas MAFs of other three SNPs; rs2010963, rs699947 and rs1570360 were observed to be considerably different (p < 0.05) between them.
3.3. Genotype Specific Implications of Genes through Different Genetic Models
Role and relevance of individual SNPs of
eNOS gene were analysed by comparing carriage of risky allele between women with osteoporosis and women without osteoporosis by taking major allele as referent (
Table 3). Genotype specific codominant, dominant, recessive and multiplicative models after adjusting the effect of variables revealed that minor allele C of rs2070744 were associated in codominant in heterozygous (OR 1.55, 95%CI: 1.18–2.19,
p = 0.007), homozygous (OR 1.73, 95%CI: 1.09–3.82,
p = 0.043), dominant (OR 1.65, 95%CI: 1.10–3.19,
p = 0.037) and multiplicative modes (OR 1.33, 95%CI: 1.14–2.00,
p = 0.005). Similarly, minor alleles T and A of rs1799983 and rs891512 respectively showed association in codominant in heterozygous (ORs 2.53 & 1.55, 95%CIs: 1.81–3.91, 1.15–2.28,
p = 0.006 &0.029), homozygous (ORs 3.00 & 2.20, 95%CIs: 1.14–8.19, 1.10–4.92,
p = 0.042 & 0.040), dominant (ORs 2.84 & 1.67, 95%CI: 1.66–5.29, 1.02–2.77,
p = 0.009 & 0.050) and multiplicative modes (ORs 2.13 & 1.51, 95%CI: 1.27–2.92, 1.13–2.09,
p = 0.002 & 0.034). SNPs rs1800780, rs3918181 and rs1808593 did not associate with osteoporosis risk.
SNP rs4459609 polymorphism of
ACE gene failed to show any association with osteoporosis risk (
Table 4) whereas, minor allele C of rs1800764 showed association in codominant homozygous (OR 1.71, 95%CI: 1.08–3.17,
p = 0.038), and multiplicative modes (OR 1.22, 95%CI: 1.01–1.62,
p = 0.043). This allele could not retain its significance in dominant mode after adjustment with the variables (
p > 0.05). Another in/del polymorphism (rs1799752) of
ACE gene showed strong association of D allele in all the possible modes i.e., codominant in heterozygous (OR 1.37, 95%CI: 1.05–2.27,
p = 0.044), homozygous (OR 2.19, 95%CI: 1.33–3.41,
p = 0.007), dominant (OR 1.68, 95%CI: 1.20–2.38,
p = 0.013), recessive (OR 1.42, 95%CI 1.03–2.13,
p = 0.028) and multiplicative (OR 1.22, 95%CI: 1.11–1.69,
p = 0.008). Another SNP rs4343 showed that the minor allele G was associated in codominant in heterozygous (OR 1.51, 95%CI: 1.14–2.17,
p = 0.009), dominant (OR 1.43, 95%CI: 1.17–2.00,
p = 0.028) and multiplicative modes (OR 1.29, 95%CI: 1.09–1.72,
p = 0.015).
Functional SNP rs2010963 of
VEGFA exhibited significant protective association in codominant in heterozygous (OR 0.60, 95%CI: 0.42–0.83,
p = 0.027), dominant (OR 0.57, 95%CI: 0.41–0.79,
p = 0.036) and multiplicative (OR 0.65, 95%CI: 0.50–0.82,
p = 0.010) modes (
Table 5). Minor allele C of SNP rs699947 was observed to be associated in codominant in homozygous (OR 1.72, 95%CI: 1.11–2.72,
p = 0.027), dominant (OR 1.30, 95%CI: 1.00–1.82,
p = 0.048), recessive (OR 1.51, 95%CI: 1.01–2.24,
p = 0.039) and multiplicative modes (OR 1.22, 95%CI: 1.05–1.61,
p = 0.027). Minor allele A of another SNP rs1570360 was observed to be associated with osteoporosis risk in codominance in heterozygous (OR 1.52, 95%CI: 1.11–2.19,
p = 0.011), homozygous (OR 2.13, 95%CI: 1.17–3.29,
p = 0.026), dominant (OR 1.69, 95%CI: 1.20–2.31,
p = 0.017) and multiplicative modes (OR 1.53, 95%CI: 1.18–1.95,
p = 0.025). None of the models showed significant association of SNP rs833061 with osteoporosis risk (
p > 0.05).
3.5. SNP-SNP Cross Talks, Risky Traits and Their Modes of Association
Several genotype specific single marker effects within SNPs of
eNOS, ACE and
VEGFA genes on risk covariates were deduced. All significant single marker effects (
p < 0.05, r > 0.04) were further analysed for pair wise epistatic effects with Bonferroni corrections (
Table 6). It was revealed that 10 SNP pair associations with risky traits were evident of osteoporosis risk. Functional SNP of eNOS gene; rs2070744 showed epistatic relationship with SNPs rs4343, rs1799983, rs1800764 and rs891512 influencing RHI (
p = 0.001), LDL (
p = 0.005), TG (0.001) and SBP (
p = 0.003) through interactive (I), additive x additive (AA), dominant × additive (DA) and additive × dominant (AD) modes respectively. Interestingly, SNP pairs rs2070744-rs1800764 impacted TG through DA mode in control subjects (
p = 0.041) also. Another SNP pairs which showed impact on the risk of osteoporosis through AA, dominant × dominant (DD) and I mode were rs4343-rs2010963 (
p = 0.009), rs1800764-rs1799752 (
p = 0.029) and rs1799983-rs891512 (
p = 0.032) influencing RHI, TC and RHI respectively. Similarly, SNP rs1799983 coupled with rs699947 (
p = 0.001), rs699947 with rs1799752 (
p = 0.003) and rs891512 with rs1799752 (
p = 0.022) to influence BMI, RHI and LDL through DD, I and DA modes, respectively. SNP pair rs1799983-rs699947 was observed to afflict DD influence in women without osteoporosis also (
p = 0.041). Two locus SNP-SNP epistatic links without risk variables have been shown in figure embedded in the
Table 6, to have quicker glance without some perplex interactions.
3.6. Haplotype Analysis, Their Contribution and Best Mode of Impact for Osteoporosis Risk
SNPs within
eNOS gene (in the order of rs2070744, rs1799983, rs1800780, rs391881, rs891512 and rs1808593) developed into 64 possible haplotypes and out of them 29 haplotypes were visible. Of these, 21 haplotypes had frequencies less than 5 percent, therefore, excluded from the analysis. The remaining eight haplotypes captured 85percent of genetic variance of women having normal bone mass and 88 percent of women having osteoporosis (
Table 7).The major alleles at position 1, 2, 5 and 6 and minor alleles at 3 and 4 of studied
eNOS SNPs appeared in the form of haplotype TGAAGT was having highest frequency, hence served as referent for the analysis. Minor alleles of all the SNPs except at position 6in the form of CTAAA
T appeared to be the risky haplotype for osteoporosis risk (OR 2.80, 95%CI: 1.53–5.13,
p = 0.001). Inter group comparisons of this haplotype after Bonferroni corrections exhibited significant differences approaching GWAS
p values (
p = 1 × 10
−8). It was observed to be susceptibility haplotype for the risk of osteoporosis (OR 2.43, 95%CI: 1.22–4.71,
p = 0.007), when its influence was examined after adjusting the effects of confounders (BMI, YSM, SBP and TG).
Haplotype analysis of SNPs in the order of rs4459609, rs1800764, rs1799752 and rs4343 of ACE gene exhibited 11 visible haplotypes but six haplotypes were excluded as these had lower frequencies (<5 percent) and were not amenable to be used for authentic results. The remaining five haplotypes showed 88–90 of genetic variability in both the groups of women. All the major alleles representing ATIA haplotype emerged to be the most common haplotype in both the groups, so it was taken as referent. All minor alleles of ACE gene SNPs, except at position 1 in the form of haplotype ACDG appeared to be risky (OR 3.03, 95%CI: 1.86–4.88, p < 0.001) and it was confirmed that it conferred 2.5 fold higher risk of developing osteoporosis in postmenopausal women having endothelial dysfunction after correcting the effect of risk predictors (OR 2.50, 95%CI: 1.28–3.96, p = 0.002).
SNPs of VEGFA gene (in the order of rs2010963, rs833061, rs699947 and rs1570360) demonstrated 11 haplotypes with five haplotypes having the least frequencies (<5 percent) hence excluded, leaving six haplotypes that were included for further analysis. Haplotype analysis after taking haplotype GCTG as reference, revealed that GATA is a risky (OR 2.83, 95%CI: 1.65–4.86, p < 0.001) whereas CACG is a protective haplotype (OR 0.51, 95%CI: 0.28–0.92, p = 0.035) for endothelial dysfunction associated osteoporosis risk. However, CACG could not retain its significance (OR 0.78, 95%CI: 0.45–1.33, p = 0.43) whereas, haplotype GATA maintained its significance after adjusting the effects of risk variables (OR 2.10, 95%CI: 1.31–3.29, p = 0.009).
Although the results implied that those postmenopausal women who possessed these susceptibility haplotypes were at higher risk of developing osteoporosis than those women who did not have it, in which best possible way these haplotypes inflicted their maximum effects needed to be identified (
Table 8). Their functional effects on BMD were modeled and tested with Wald statistics under dominant, recessive, multiplicative and general modes of inheritance and selection of the best fit model was identified with AIC and
R2h (
Table 8). Analysis clarified that susceptibility haplotype CTAAAT of eNOS gene influenced osteoporosis risk in multiplicative mode (β ± SE: 2.19 ± 0.86,
p < 0.001), haplotypes ACDG of ACE gene (β ± SE: 1.73 ± 0.54,
p = 0.001) and haplotype GATA of VEGFA gene (β ± SE: 3.07 ± 0.81,
p < 0.001) influenced bone mass in postmenopausal women in dominant modes.
3.7. Predictive Ability of Haplotypes and Traditional Risk Factors for the Diagnosis of Osteoporosis
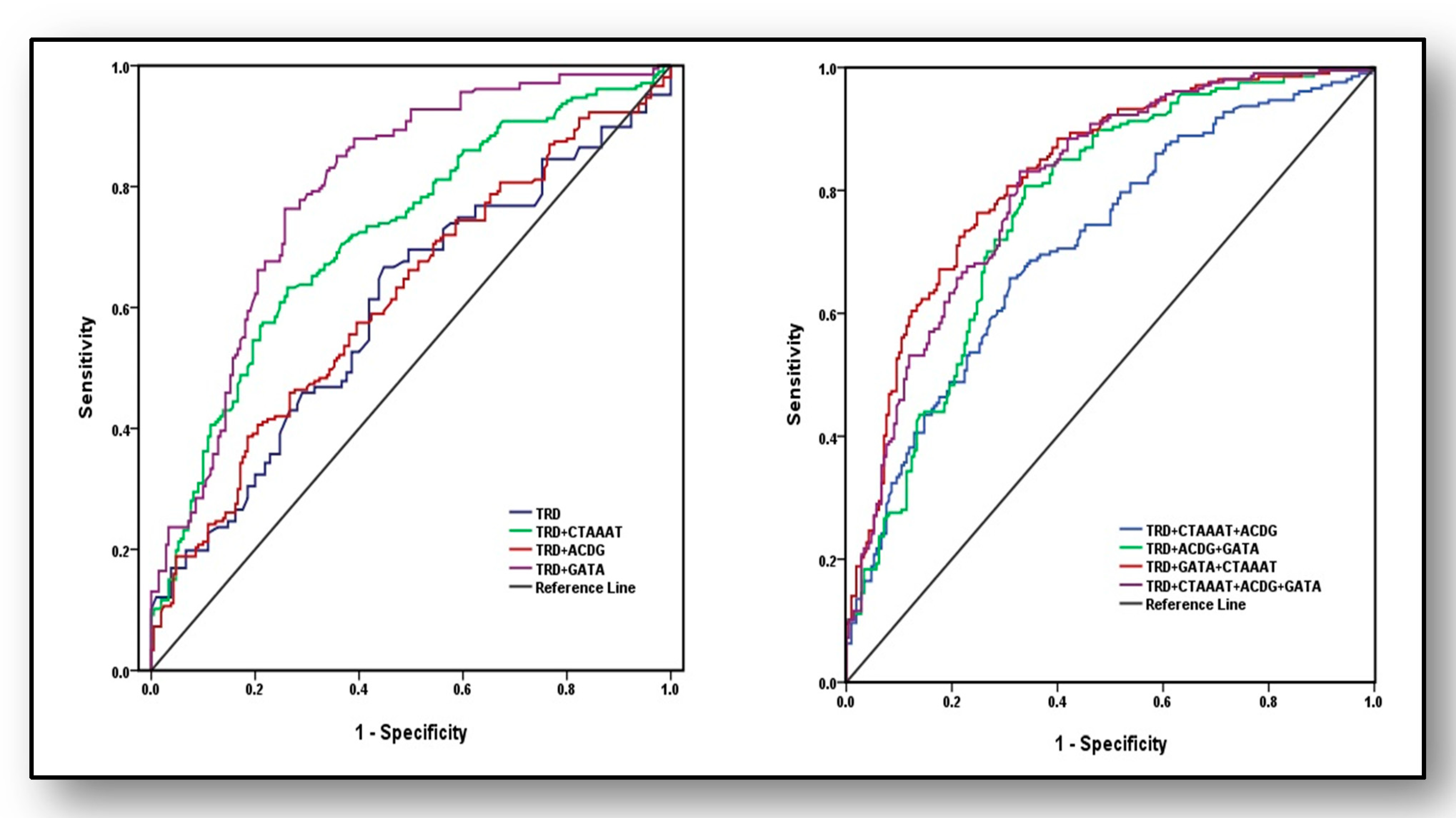
Predictive strengths of susceptibility haplotypes and/or traditional risk factors (TRD) for the diagnosis of osteoporosis were analysed (
Figure 3) using AUROC curve by including weighted risk scores (effect estimates). In the first set, four models were used; only TRD, TRD plus haplotype CTAAAT, TRD plus haplotype ACDG and TRD plus haplotype GATA. Area under curve (AUC) revealed marginal and weak analytical power for TRD (AUC = 0.60 ± 0.028, 95%CI: 0.545–0.654,
p < 0.001), which increased to acceptable limits (AUC = 0.72 ± 0.025, 95%CI: 0.670–0.767,
p < 0.001) after adding risk scores of haplotype CTAAAT of eNOS gene. Its predictive power marginally increased when TRD were modeled with haplotype ACDG of ACE gene (AUC = 0.614 ± 0.027, 95%CI: 0.560–0.668,
p < 0.001) whereas, its predictive ability showed maximum strength with haplotype GATA (AUC = 0.8 ± 0.022, 95%CI: 0.751–0.838,
p < 0.001). In the second set, AUROC was analysed with four different models: (i) TRD + CTAAAT + ACDG, (ii) TRD + ACDG + GATA, (iii) TRD + GATA + CTAAAT and (iv) TRD + CTAAAT + ACDG + GATA, which exposed that third model (TRD + GATA+ CTAAAT) had the highest ability to predict osteoporosis (AUC = 0.824 ± 0.020, 95%CI: 0.784–0.864,
p < 0.001) followed by fourth model including TRD + CTAAAT + ACDG + GATA (AUC = 0.806 ± 0.021, 95%CI: 0.765–0.847,
p < 0.001)and TRD + ACDG + GATA (AUC = 0.769 ± 0.023, 95%CI: 0.724–0.814,
p < 0.001) and with least predictive power of the first model (AUC = 0.713 ± 0.025, 95%CI: 0.665–0.762,
p < 0.001).

 ,
, 



{kind=link}
{kind=link}
{kind=link}
