
Quorum Sensing Bacteria in the Phycosphere of HAB Microalgae and Their Ecological Functions Related to Cross-Kingdom Interactions

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Source and Pretreatment of Microalgae
2.2. Isolation and Purification of Microalgae Associated Bacteria
2.3. Screening of Quorum Sensing Bacteria
2.4. Strain Identification and Phylogenetic Analysis
2.5. Extraction and Identification of AHLs
2.6. The Biological Functions of QS Bacteria
2.6.1. Biofilm Formation
Crystal Violet Staining Method
Confocal Laser Scanning Microscope Detection Method (CLSM)
2.6.2. DMSP Consumption
2.6.3. Detection of Microalgae Cell Density
3. Results
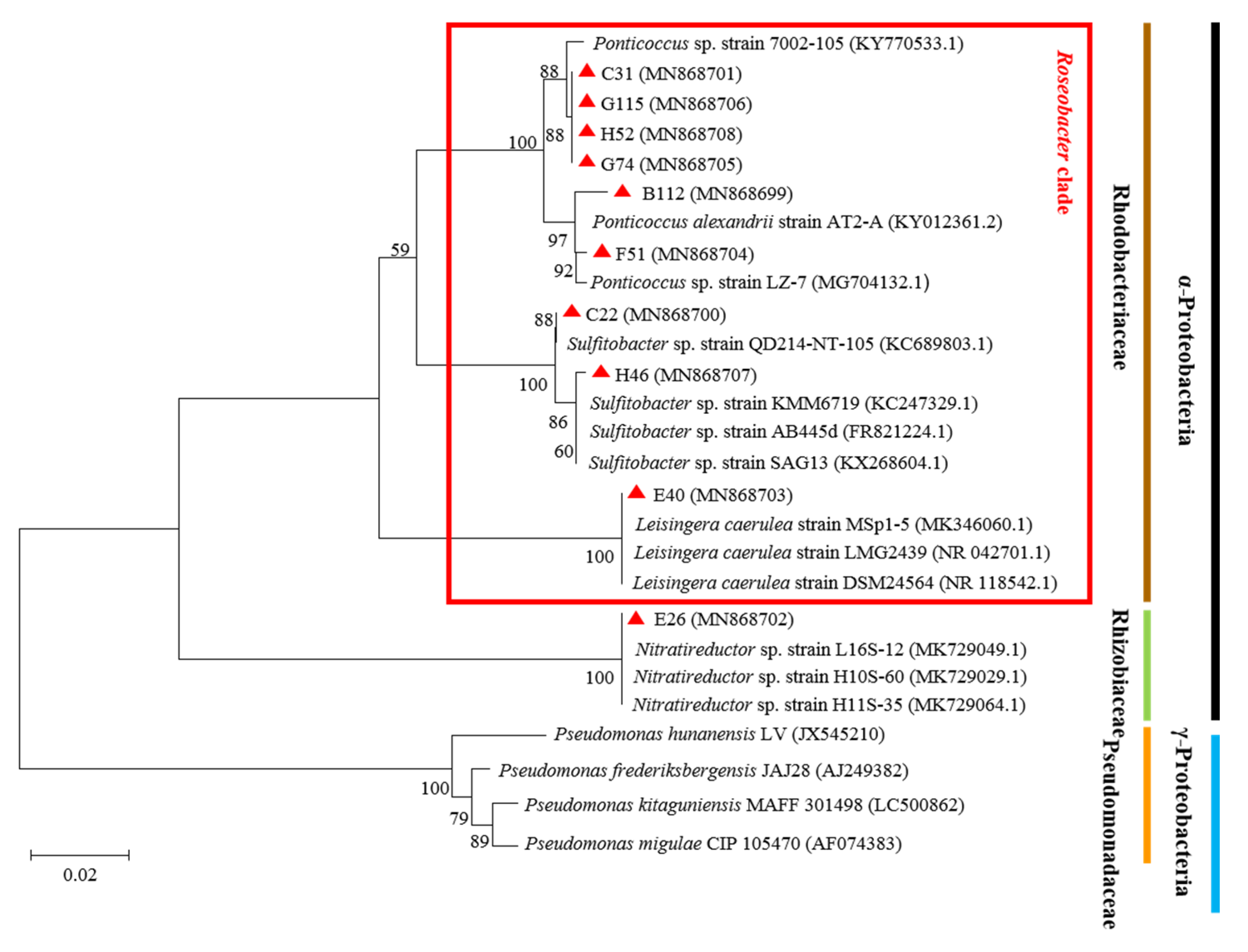
3.1. Molecular Identification and Phylogenetic Tree Analysis of QS Bacteria
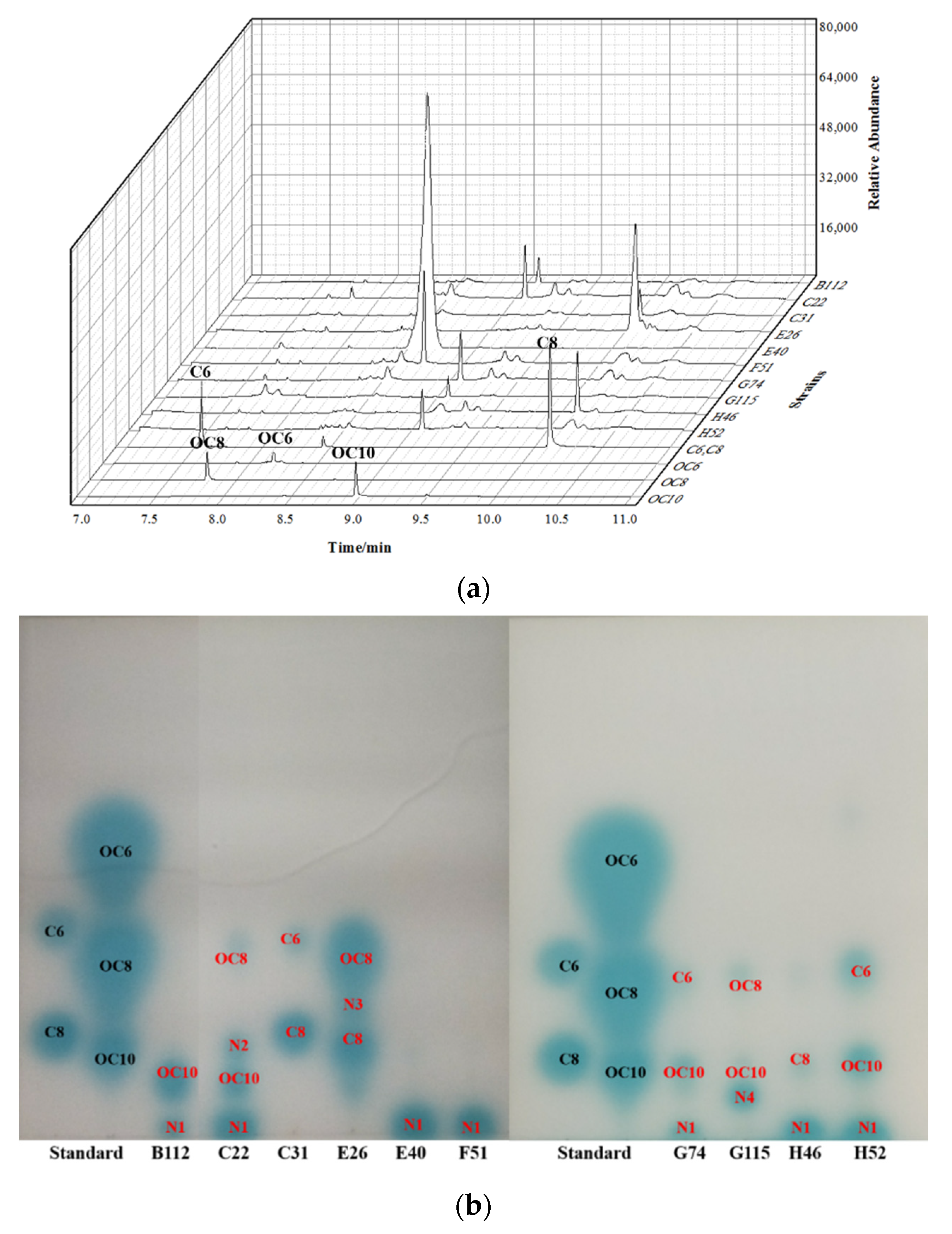
3.2. Profile of the AHL Signalling Molecules
3.3. Ecological Functions of the QS Bacteria
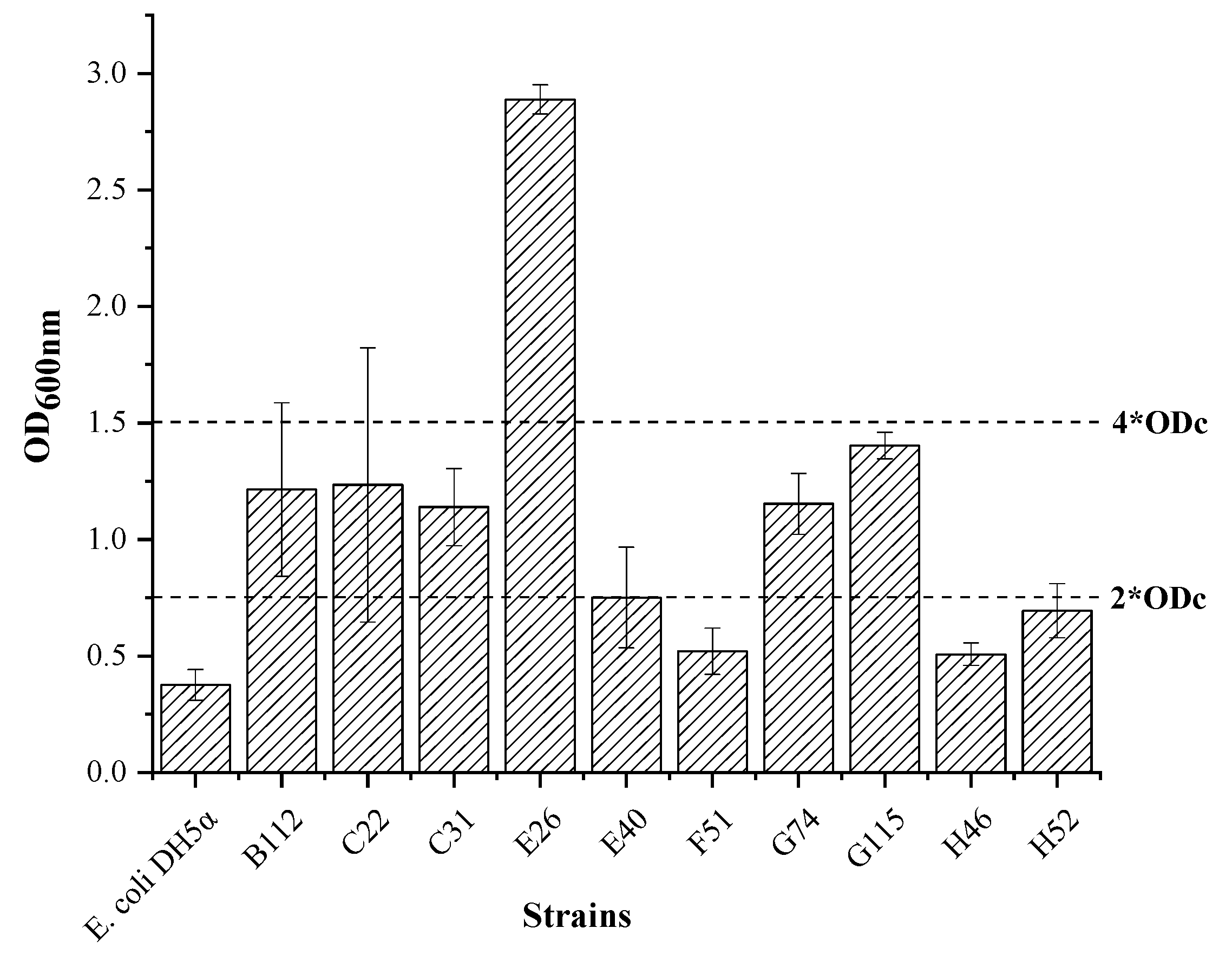
3.3.1. Film-Forming Ability of the QS Bacteria
3.3.2. Utilization of DMSP by QS Bacteria
3.3.3. The Influence of QS Bacteria on the Growth of Microalgae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Hill, R.T.; Zheng, T.; Hu, X.; Wang, B. Effects of bacterial communities on biofuel-producing microalgae: Stimulation, inhibition and harvesting. Crit. Rev. Biotechnol. 2016, 36, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.H.; Tang, L.L.; Zhang, Y.Y. Algae-bacteria interactions and their ecological functions in the ocean. Microbiology 2018, 45, 2043–2053. [Google Scholar]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between diatoms and bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie-Oleza, J.A.; Sousoni, D.; Lloyd, M.; Armengaud, J.; Scanlan, D.J. Nutrient recycling facilitates long-term stability of marine microbial phototroph-heterotroph interactions. Nat. Microbiol. 2017, 2, 17100. [Google Scholar] [CrossRef] [Green Version]
- Seymour, J.R.; Amin, S.A.; Raina, J.B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef] [PubMed]
- De-Bashan, L.E.; Antoun, H.; Bashan, Y. Involvement of indole-3-acetic acid produced by the growth-promoting bacterium Azospirillum spp. in promoting growth of Chlorella vulgaris. J. Phycol. 2008, 44, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.A.; Green, D.H.; Hart, M.C.; Kupper, F.C.; Sunda, W.G.; Carrano, C.J. Photolysis of iron-siderophore chelates promotes bacterial-algal mutualism. Proc. Natl. Acad. Sci. USA 2009, 106, 17071–17076. [Google Scholar] [CrossRef] [Green Version]
- Cnuz-López, R.; Maske, H. The vitamin B1 and B12 required bythe marine dinoflagellate Lingulodinium polyedrum can be provided by its associated bacterial community in culture. Front. Microbiol. 2016, 7, 560. [Google Scholar]
- Sonnenschein, E.C.; Syit, D.A.; Grossart, H.P.; Ullrich, M.S. Chemotaxis of Marinobacter adhaerens and its impact on attachment to the diatom Thalassiosira weissflogi. Appl. Environ. Microbiol. 2012, 78, 6900–6907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twigg, M.S.; Tait, K.; Williams, P.; Atkinson, S.; Cámara, M. Interference with the germination and growth of Ulva zoospores by quorum-sensing molecules from UIva-associated epiphytic bacteria. Environ. Microbiol. 2014, 16, 445–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klindworth, A.; Mann, A.J.; Huang, S.X.; Wichels, A.; Quast, C.; Waldmann, J.; Teeling, H.; Glöckner, F.O. Diversity and activity of marine bacterioplankton during a diatom bloom in the North Sea assessed by total RNA and pyrotag sequencing. Mar. Genom. 2014, 18, 185–192. [Google Scholar] [CrossRef]
- Needham, D.M.; Fuhrman, J.A. Pronounced daily succession of phytoplankton, archaea and bacteria following a spring bloom. Nat. Microbiol. 2016, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Daft, M.J.; McCord, S.B.; Stewart, W.D.P. Ecological studies on algal-lysing bacteria in fresh waters. Freshw. Biol. 1975, 5, 577–596. [Google Scholar] [CrossRef]
- Mitsutani, A.; Yamasaki, I.; Kitaguchi, H. Analysis of algicidal proteins of a diatom-lytic marine bacterium Pseudoalteromonas sp. strain A25 by two-dimensional electrophoresis. Phycologia 2001, 40, 286–291. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Findlay, J.A.; Patil, A.D. Antibacterial constituents of the diatom Navicula delognei. Indian J. Nat. Prod. 1984, 47, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Vidoudez, C.; Nejstgaard, J.C.; Jakobsen, H.H.; Pohnert, G. Dynamics of dissolved and particulate polyunsaturated aldehydes in mesocosms inoculated with different densities of the diatom Skeletonema marinoi. Mar. Drugs 2011, 9, 345–358. [Google Scholar] [CrossRef] [Green Version]
- Paul, C.; Pohnert, G. Production and role of volatile halogenated compounds from marine algae. Nat. Prod. Rep. 2011, 28, 186–195. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnard, A.M.L.; Salmond, G.P.C. Quorum sensing in Erwinia species. Anal. Bioanal. Chem. 2007, 387, 415–423. [Google Scholar] [CrossRef]
- Montaner, B.; Pérez-Tomás, R. Prodigiosin-induced apoptosis in human colon cancer cells. Life Sci. 2001, 68, 2025–2036. [Google Scholar] [CrossRef]
- Steindler, L.; Venturi, V. Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol. Lett. 2007, 266, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Shaw, P.D.; Ping, G.; Daly, S.L.; Cha, C.; Cronan, J.E.; Rinehart, K.L.; Farrand, S.K. Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layer chromatography. Proc. Natl. Acad. Sci. USA 1997, 94, 6036–6041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortori, C.A.; Dubern, J.F.; Chhabra, S.R.; Cámara, M.; Hardie, K.; Williams, P.; Barrett, D.A. Simultaneous quantitative profiling of N-acyl-l-homoserine lactone and 2-alkyl-4(1H)-quinolone families of quorum-sensing signaling molecules using LC-MS/MS. Anal. Bioanal. Chem. 2011, 399, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Wagner-Döbler, I.; Ballhausen, B.; Berger, M.; Brinkhoff, T.; Buchholz, I.; Bunk, B.; Cvpionka, H.; Daniel, R.; Drepper, T.; Gerdts, G.; et al. The complete genome sequence of the algal symbiont Dinoroseobacter shibae: A hitchhiker’s guide to life in the sea. ISME J. 2010, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Hube, A.E.; Hevduck-soller, B.; Fischer, U. Phvlogenetic classification of heterotrophic bacteria associated with filamentous marine cyanobacteria in culture. Syst. Appl. Microbiol. 2009, 32, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Cude, W.N.; Mooney, J.; Tavanaei, A.A.; Hadden, M.K.; Frank, A.M.; Gulvik, C.A.; May, A.L.; Buchan, A. Production of the antimicrobial secondary metabolite indigoidine contributes to competitive surface colonization by the marine roseobacter Phaeobacter sp. strain Y4I. Appl. Environ. Microbiol. 2012, 78, 4771–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, H.; Belas, R. Molecular mechanisms underlying Roseobacter–phytoplankton symbioses. Curr. Opin. Biotechnol. 2010, 21, 332–338. [Google Scholar] [CrossRef]
- Lv, H.; Zhou, J.; Cai, Z.H. Dynamic variation of quorum sensing bacteria in a Scrppsiella trochoidea bloom. Ecol. Sci. 2016, 35, 23–30. [Google Scholar]
- Huang, X.Q.; Cai, Z.H.; Lin, G.H.; Zhou, J. Quorum sensing modulating algae–bacteria interactions. Chin. J. Appl. Environ. Biol. 2016, 22, 0708–0717. [Google Scholar]
- Xu, L.Y.; Zheng, L.; Han, X.T.; Cui, Z.S.; Guo, X.C.; Li, X.Z. Screening of microalgae associated bacteria with quorum sensing system and there algicidal activity. Oceanolgia Et Limnol. Sin. 2012, 43, 1149–1155. [Google Scholar]
- Chi, W.D.; Zheng, L.; He, C.F.; Han, B.; Zheng, M.G.; Gao, W.; Sun, C.J.; Zhou, G.F.; Gao, X.X. Quorum sensing of microalgae associated marine Ponticoccus sp. PD-2 and its algicidal function regulation. AMB Express 2017, 7, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. Nucleic Acid Tech. Bact. Syst. 1991, 115–175. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.T.; Cowles, K.; Bermejo, M.A.; Curson, A.R.J.; Zheng, Y.Y.; Liu, J.L.; Newton-Payne, S.; Hind, A.J.; Li, C.Y.; Rivera, P.P.L.; et al. Bacteria are important dimethylsulfoniopropionate producers in coastal sediments. Nat. Microbiol. 2019, 4, 1815–1825. [Google Scholar] [CrossRef] [Green Version]
- Reisch, C.R.; Moran, M.A.; Whitman, W.B. Bacterial catabolism of dimethylsulfoniopropionate (DMSP). Front. Microbiol. 2011, 2, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyenal, H.; Lewandowski, Z.; Harkin, G. Quantifying biofilm structure: Facts and fiction. Biofouling 2004, 20, 1–23. [Google Scholar] [CrossRef]
- Chen, L.; Wang, B.; Yao, M.M.; CaI, Z.H. Construction and Significance of Quantitative Analysis of Microbial Biofilm Based on COMSTAT Program. Prog. Mod. Biomed. 2014, 1201–1206. (In Chinese) [Google Scholar] [CrossRef]
- Buchan, A.; Gonzalez, J.M.; Moran, M.A. Overview of the Marine Roseobacter Lineage. Appl. Environ. Microbiol. 2005, 71, 5665–5677. [Google Scholar] [CrossRef] [Green Version]
- Alavi, M.; Miller, T.; Erlandson, K.; Schneider, R.; Belas, R. Bacterial community associated with Pfiesteria-like dinoflagellate cultures. Environ. Microbiol. 2001, 3, 380–396. [Google Scholar] [CrossRef] [PubMed]
- Zan, J.; Liu, Y.; Fuqua, C.; Hill, R. Acyl-homoserine lactone quorum sensing in the Roseobacter clade. Int. J. Mol. Sci. 2014, 15, 654–669. [Google Scholar] [CrossRef]
- Wagner-Döbler, I.; Thiel, V.; Eberl, L.; Allgaier, M.; Bodor, A.; Meyer, S.; Ebner, S.; Hennig, A.; Pukall, R.; Schulz, S. Discovery of complex mixtures of novel long-chain quorum sensing signals in free-living and host-associated marine alphaproteobacteria. Chembiochem 2005, 6, 2195–2206. [Google Scholar] [CrossRef]
- Decho, A.W.; Visscher, P.T.; Ferry, J.; Kawaguchi, T.; He, L.; Przekop, K.M.; Norman, R.S.; Reid, R.P. Autoinducers extracted from microbial mats reveal a surprising diversity of N-acyl homoserine lactones (AHLs) and abundance changes that may relate to diel pH. Environ. Microbiol. 2009, 11, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Rivas, M.O.; Vargas, P.; Riquelme, C.E. Interactions of Botryococcus braunii cultures with bacterial biofilms. Microb. Ecol. 2010, 60, 628–635. [Google Scholar] [CrossRef]
- Bi, X.D.; Zhou, W.L.; Xing, K.Z.; Tang, X.X. Effects of AHLs (N-hexanoyl-DL-homoserine lactone, C6-HSL) on growth and antioxidant defense system of Chlorella vulgaris. Mar. Environ. Sci. 2012, 31, 897–900. [Google Scholar]
- Zhou, J.; Lyu, Y.; Richlen, M.L.; Anderson, D.M.; Cai, Z. Quorum sensing is a language of chemical signals and plays an ecological role in algal-bacterial interactions. Crit. Rev. Plant Sci. 2016, 35, 81–105. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, M.; Yang, L.; Su¨nje, J.P.; Tolker-Nielsen, T. An update on Pseudomonas aeruginosa biofilm formation, tolerance, and dispersal. FEMS Immunol. Med. Microbiol. 2010, 59, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, M.; Bangera, M.G.; Bumgarner, R.E.; Parsek, M.R.; Greenberg, E.P. Gene expression in Pseudomonas aeruginosa biofilms. Nature 2001, 413, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Küpper, F.C.; Müller, D.G.; Peters, A.F.; Potin, P. Oligoalginate recognition and oxidative burst play a key role in natural and induced resistance of sporophytes of Laminariales. J. Chem. Ecol. 2002, 28, 2057–2081. [Google Scholar] [CrossRef]
- Natrah, F.M.I.; Defoirdt, T.; Sorgeloos, P.; Bossier, P. Disruption of bacterial cell-to-cell communication by marine organisms and its relevance to aquaculture. Mar. Biotechnol. 2011, 13, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Geng, H.; Belas, R. Expression of tropodithietic acid biosynthesis is controlled by a novel autoinducer. J. Bacteriol. 2010, 192, 4377–4387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, J.P. Bioactive compound synthetic capacity and ecological significance of marine bacterial genus Pseudoalteromonas. Mar. Drugs 2007, 5, 220–241. [Google Scholar] [CrossRef] [PubMed]
- Kiene, R.P.; Linn, L.J.; Bruton, J.A. New and important roles for DMSP in marine microbial communities. J. Sea Res. 2000, 43, 209–224. [Google Scholar] [CrossRef]
- Curson, A.; Rogers, R.; Todd, J.D.; Brearley, C.A.; Johnston, A. Molecular genetic analysis of a dimethylsulfoniopropionate lyase that liberates the climate-changing gas dimethylsulfide in several marine α-proteobacteria and Rhodobacter sphaeroides. Environ. Microbiol. 2008, 10, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Raina, J.B.; Tapiolas, D.; Willis, B.L.; Bourne, D.G. Coral-associated bacteria and their role in the biogeochemical cycling of sulfur. Appl. Environ. Microbiol. 2009, 75, 3492–3501. [Google Scholar] [CrossRef] [Green Version]
- Johnson, W.M.; Soule, M.C.K.; Kujawinski, E.B. Evidence for quorum sensing and differential metabolite production by a marine bacterium in response to DMSP. ISME J. 2016, 10, 2304–2316. [Google Scholar] [CrossRef] [Green Version]
- Reisch, C.R.; Crabb, W.M.; Gifford, S.M.; Teng, Q.; Whitman, W.B. Metabolism of dimethylsulphoniopropionate by Ruegeria pomeroyi DSS-3. Mol. Microbiol. 2013, 89, 774–791. [Google Scholar] [CrossRef] [PubMed]
- Kiene, R.P.; Linn, L.J.; González, J.; Moran, A.; Bruton, J.A. Dimethylsulfoniopropionate and methanethiol are important precursors of methionine and protein-sulfur in marine bacterioplankton. Appl. Environ. Microbiol. 1999, 65, 4549–4558. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, T.; Miyazaki, Y.; Matsuyama, Y.; Muraoka, W.; Yamaguchi, K.; Oda, T. Producing mechanism of an algicidal compound against red tide phytoplankton in a marine bacterium γ-proteobacterium. Appl. Microbiol. Biotechnol. 2006, 73, 684–690. [Google Scholar] [CrossRef]
- Guo, X.; Liu, X.; Wu, L.; Pan, J.; Yang, H. The algicidal activity of Aeromonas sp. strain GLY-2107 against bloom-forming Microcystis aeruginosa is regulated by N-acyl homoserine lactone-mediated quorum sensing. Environ. Microbiol. 2016, 18, 3867–3883. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, G.F.; Zhu, X.S.; Chen, L.; Cai, Z.H. A review of the relationship between algae and bacteria in harmful algal blooms. Acta Ecol. Sin. 2014, 34, 269–281. [Google Scholar]
- Hong, Y.; Hu, H.Y.; Xie, X.; Li, F.M. Responses of enzymatic antioxidants and non-enzymatic antioxidants in the cyanobacterium Microcystis aeruginosa to the allelochemical ethyl 2-methyl acetoacetate (EMA) isolated from reed (Phragmites communis). J. Plant Physiol. 2008, 165, 1264–1273. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain (Accession Number) | Closest Match in GenBank (Accession Number) | Similarity (%) | Alignment Fragment Size (bp) |
|---|---|---|---|
| B112 (MN868699) | Ponticoccus alexandrii strain AT2-A (KY012361.2) | 99 | 1335 |
| C22 (MN868700) | Sulfitobacter sp. strain QD214-NT-105 (JQ675546.1) | 99 | 1319 |
| C31 (MN868701) | Ponticoccus sp. strain 7002-105 (KY770533.1) | 99 | 1321 |
| E26 (MN868702) | Nitratireductor sp. strain L16S-12 (KY770526.1) | 99 | 1365 |
| E40 (MN868703) | Leisingera caerulea strain DSM 24564 (NR_118542.1) | 99 | 1309 |
| F51 (M868704) | Ponticoccus sp. LZ-7 (MG704132.1) | 99 | 1318 |
| G74 (MN868705) | Ponticoccus sp. strain 7002-105 (KY770533.1) | 99 | 1305 |
| G115 (MN868706) | Ponticoccus sp. strain 7002-105 (KY770533.1) | 99 | 1319 |
| H46 (MN868707) | Sulfitobacter sp. strain KMM6719 (KC2247329.1) | 100 | 1319 |
| H52 (MN868708) | Ponticoccus sp. strain 7002-105 (KY770533.1) | 99 | 1345 |
| Biomass (mg/cm3) | The Average Thickness (μm) | Average Diffusion Distance | Surface Area to Volume Ratio (μm2/μm3) | |
|---|---|---|---|---|
| B112 | 7.311 ± 0.195 | 18.687 ± 0.407 | 0.064 ± 0.001 | 2.722 ± 0.02 |
| C22 | 5.918 ± 0.344 | 16.210 ± 0.920 | 0.068 ± 0.003 | 2.430 ± 0.020 |
| C31 | 7.877 ± 0.090 | 20.785 ± 1.454 | 0.053 ± 0.003 | 2.369 ± 0.092 |
| E26 | 7.067 ± 0.219 | 22.132 ± 0.277 | 0.046 ± 0.002 | 2.110 ± 0.093 |
| E40 | 5.274 ± 0.168 | 14.134 ± 0.904 | 0.048 ± 0.004 | 2.285 ± 0.076 |
| F51 | 5.367 ± 0.715 | 15.874 ± 2.107 | 0.059 ± 0.006 | 2.622 ± 0.044 |
| G115 | 8.686 ± 0.390 | 23.928 ± 1.255 | 0.102 ± 0.008 | 1.712 ± 0.110 |
| G74 | 6.634 ± 1.570 | 18.707 ± 4.686 | 0.061 ± 0.018 | 2.622 ± 0.026 |
| H46 | 5.142 ± 0.713 | 13.157 ± 1.710 | 0.110 ± 0.013 | 2.268 ± 0.043 |
| H52 | 7.844 ± 0.720 | 21.793 ± 2.626 | 0.048 ± 0.007 | 2.323 ± 0.014 |
| E. coli DH5α | 0.429 ± 0.547 | 0.2594 ± 0.636 | 0.012 ± 0.824 | 4.386 ± 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zheng, L.; Wang, S.; Zhao, Y.; Xu, X.; Han, B.; Hu, T. Quorum Sensing Bacteria in the Phycosphere of HAB Microalgae and Their Ecological Functions Related to Cross-Kingdom Interactions. Int. J. Environ. Res. Public Health 2022, 19, 163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19010163
Zhang Y, Zheng L, Wang S, Zhao Y, Xu X, Han B, Hu T. Quorum Sensing Bacteria in the Phycosphere of HAB Microalgae and Their Ecological Functions Related to Cross-Kingdom Interactions. International Journal of Environmental Research and Public Health. 2022; 19(1):163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19010163
Chicago/Turabian StyleZhang, Yanchao, Li Zheng, Shuai Wang, Yangguo Zhao, Xiyuan Xu, Bin Han, and Tianyi Hu. 2022. "Quorum Sensing Bacteria in the Phycosphere of HAB Microalgae and Their Ecological Functions Related to Cross-Kingdom Interactions" International Journal of Environmental Research and Public Health 19, no. 1: 163. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19010163