
Transcriptome and HPLC Analysis Reveal the Regulatory Mechanisms of Aurantio-Obtusin in Space Environment-Induced Senna obtusifolia Lines

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. HPLC Analysis
2.3. RNA Extraction and Sequencing
2.4. Sequence Alignment and Bioinformation Analysis
2.5. Identification of Differentially Expressed Genes (DEGs)
2.6. Expression Analysis of Key Enzyme Genes
2.7. Protein Association Network Relationship Prediction
2.8. Validation of DEGs by qRT–PCR
3. Results
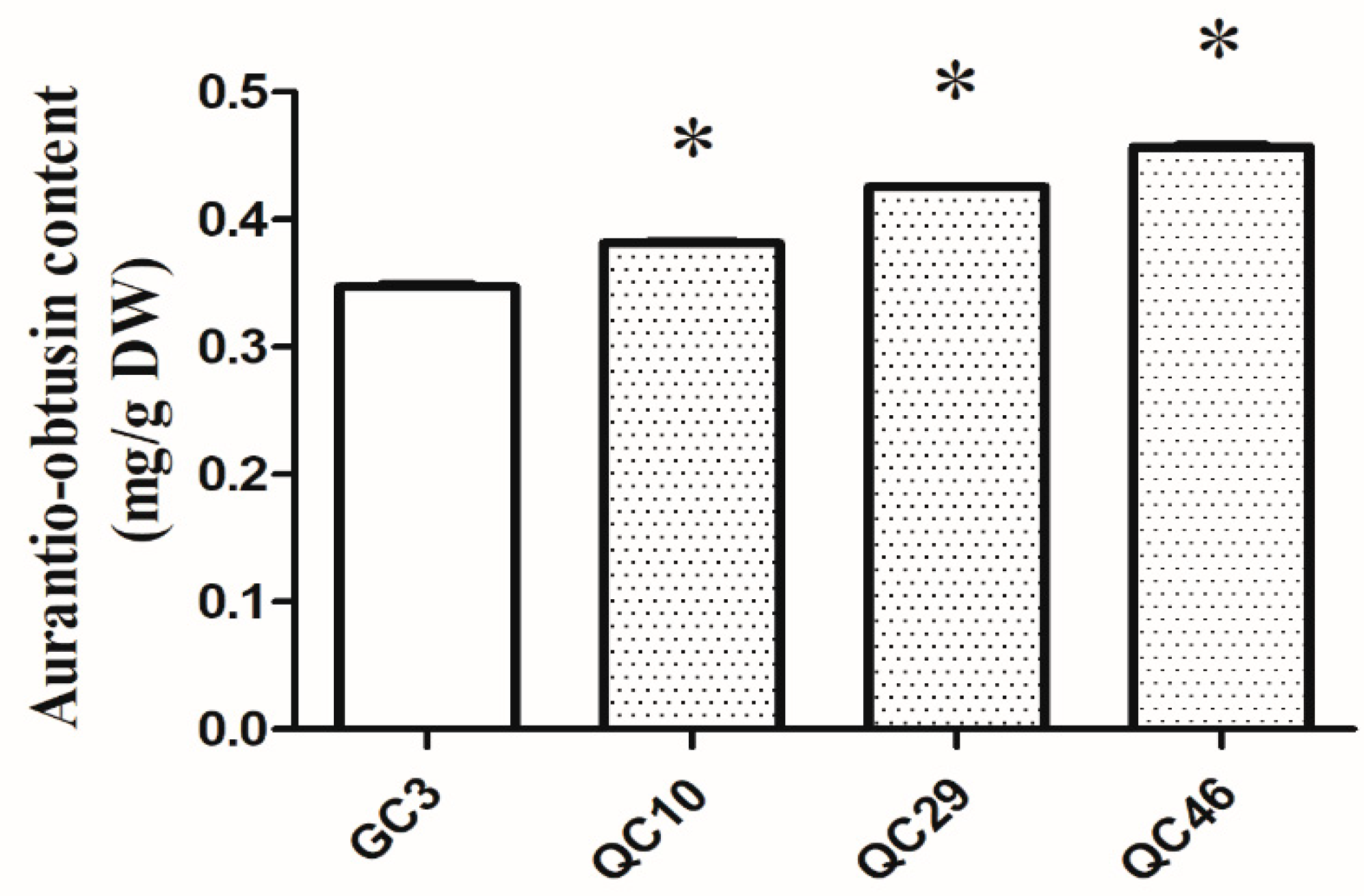
3.1. AO Content in SP-Lines and GC3
3.2. Transcriptome Features
3.3. Functional Annotation and Classification
3.4. DEGs Identification between SP-Lines and GC3
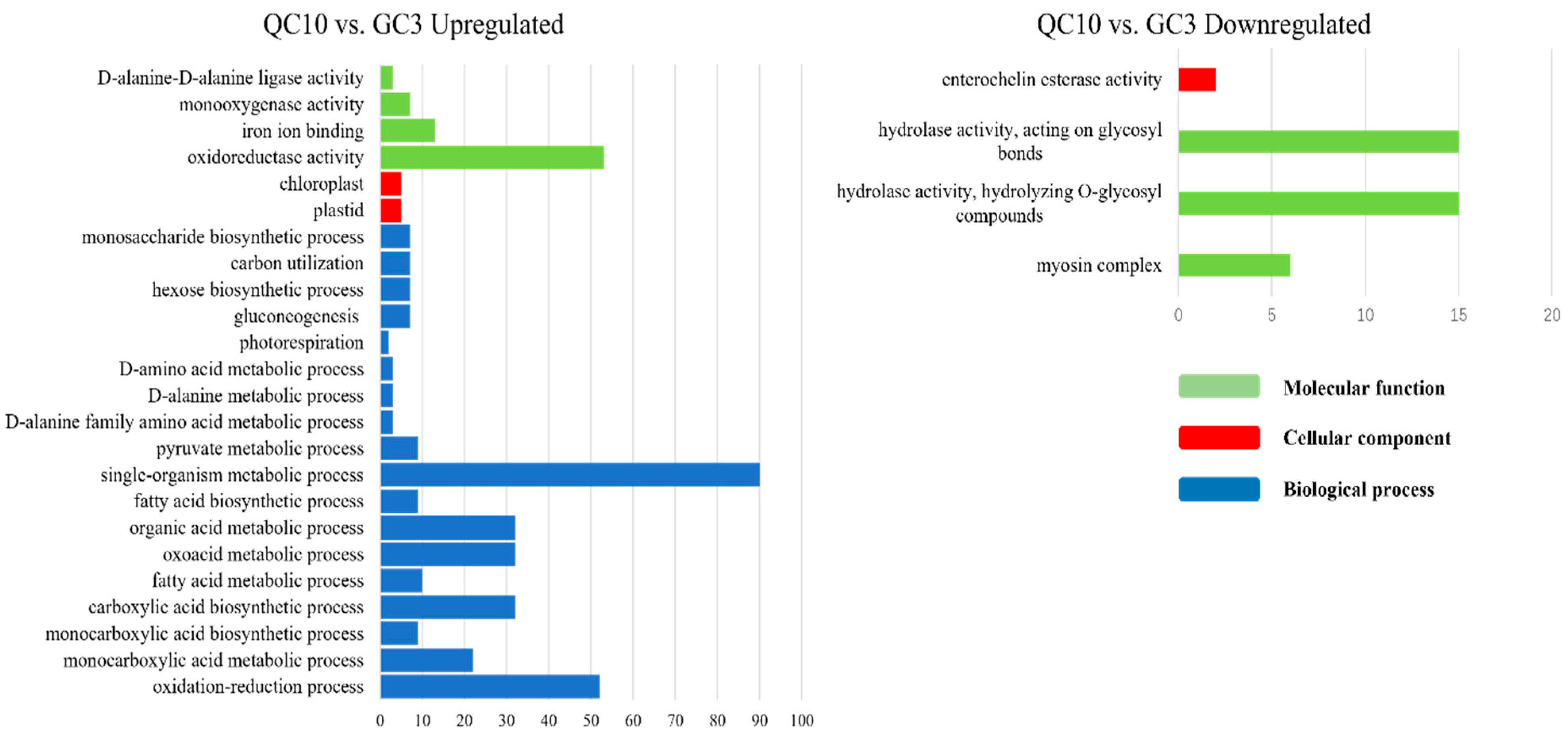
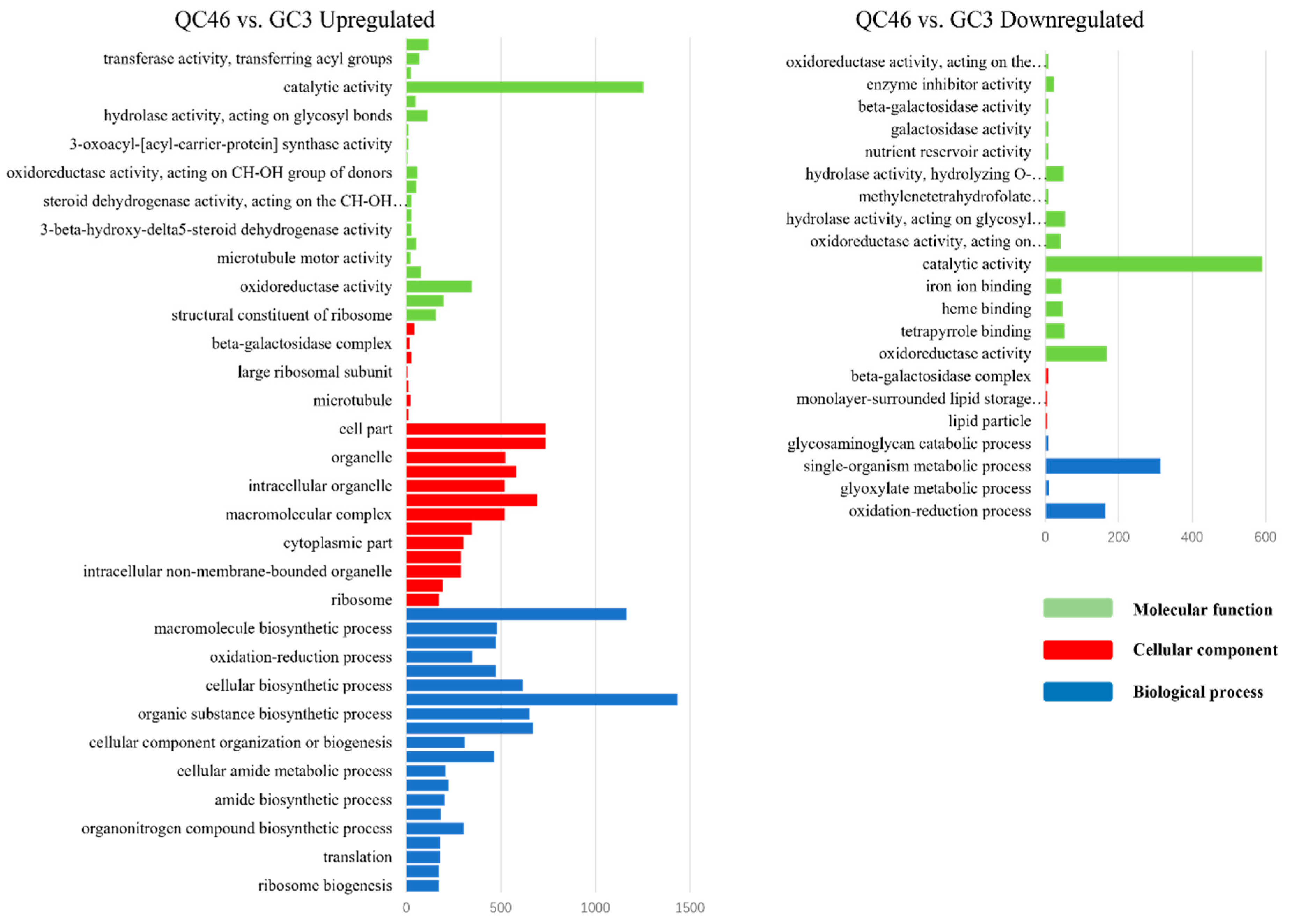
3.5. GO Enrichment of DEGs
3.6. KEGG Pathway Analysis of DEGs
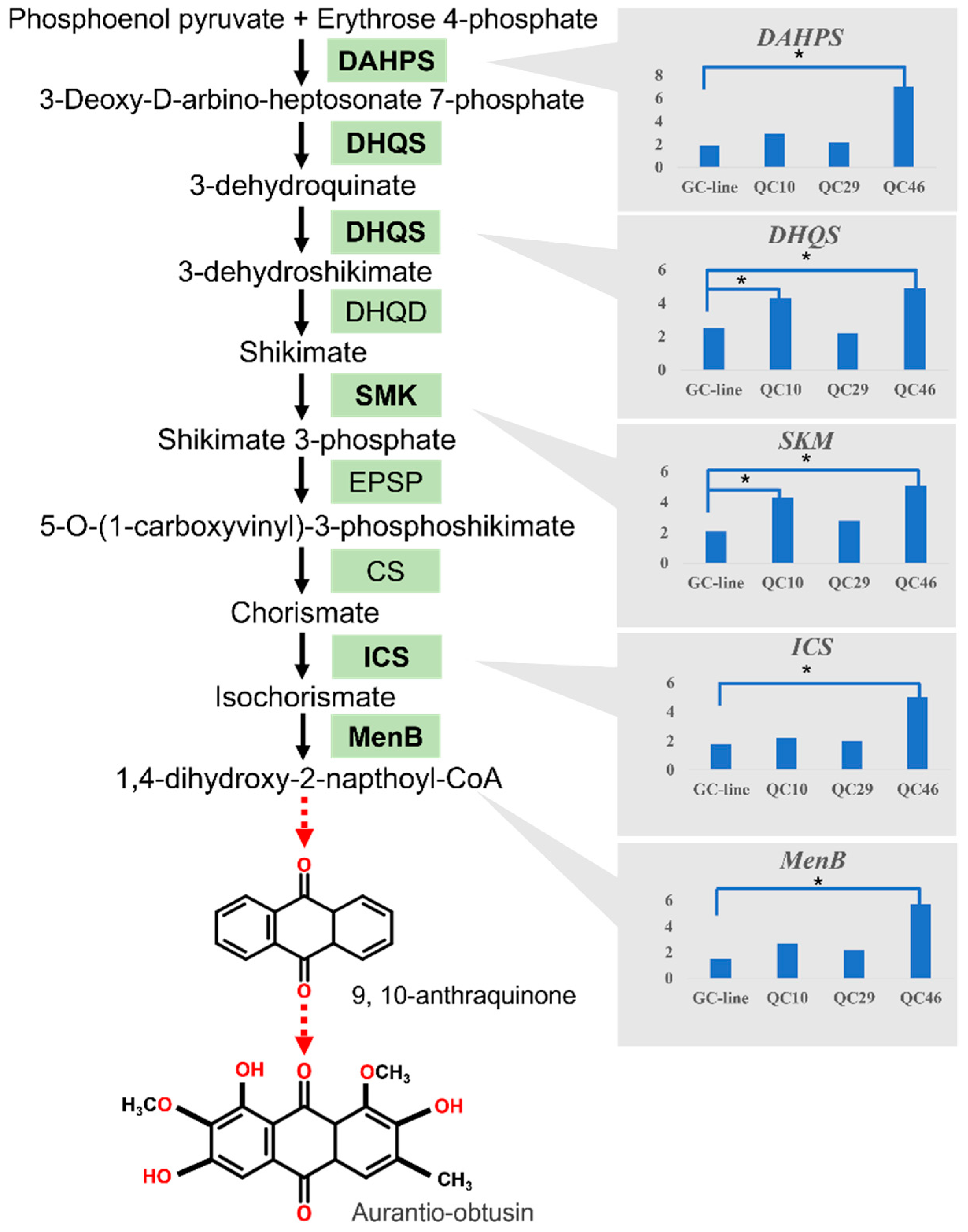
3.7. DEGs Involved in Anthraquinone Biosynthesis Pathway
3.8. Expression Analysis of Key Enzyme Genes in the Shikimate Acid Pathway
3.9. Protein Interaction Network Analysis
3.10. Transcriptome Validation
4. Discussion
4.1. Secondary Metabolites in Space Environment-Induced Medicinal Plants Were Altered Significantly
4.2. De Novo Assembly of S. obtusifolia Seeds Transcriptome
4.3. SP-Lines Have Different AO Accumulation Mechanisms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, F.; Cheng, L.; Cao, Y.X.; Wei, Q.S.; Tong, C.Y.; Shi, S.Y. Online extraction and enrichment coupling with high-speed counter-current chromatography for effective and target isolation of antitumor anthraquinones from seeds of Cassia obtusifolia. J. Sep. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Shih, Y.H.; Chen, F.A.; Wang, L.F.; Hsu, J.L. Discovery and study of novel antihypertensive peptides derived from Cassia obtusifolia seeds. J. Agric. Food Chem. 2019, 67, 7810–7820. [Google Scholar] [CrossRef]
- Kwon, K.S.; Lee, J.H.; So, K.S.; Park, B.K.; Lim, H.; Choi, J.S.; Kim, H.P. Aurantio-obtusin, an anthraquinone from cassiae semen, ameliorates lung inflammatory responses. Phytother. Res. 2018, 32, 1537–1545. [Google Scholar] [CrossRef]
- Meng, Y.Y.; Liu, Y.; Fang, N.N.; Guo, Y.M. Hepatoprotective effects of cassia semen ethanol extract on non-alcoholic fatty liver disease in experimental rat. Pharm. Biol. 2019, 57, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Yuen, H.; Hung, A.; Yang, A.W.H.; Lenon, G.B. Mechanisms of action of cassia semen for weight management: A computational molecular docking study of serotonin receptor 5-HT2C. Int. J. Mol. Sci. 2020, 21, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, S.; Rimpy; Ahuja, M. Carboxymethyl modification of Cassia obtusifolia galactomannan and its evaluation as sustained release carrier. Int. J. Biol. Macromol. 2020, 164, 3823–3834. [Google Scholar] [CrossRef]
- Napierała, M.; Nawrot, J.; Gornowicz-Porowska, J.; Florek, E.; Moroch, A.; Adamski, Z.; Kroma, A.; Miechowicz, I.; Nowak, G. Separation and HPLC characterization of active natural steroids in a standardized extract from the Serratula coronata herb with antiseborrheic dermatitis activity. Int. J. Environ. Res. Public Health 2020, 17, 6453. [Google Scholar] [CrossRef]
- Ibrahim, N.I.; Wong, S.K.; Mohamed, I.N.; Mohamed, N.; Chin, K.Y.; Ima-Nirwana, S.; Shuid, A.N. Wound healing properties of selected natural products. Int. J. Environ. Res. Public Health 2018, 15, 2360. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Liu, Z.; Li, D.; Yao, R.; Hou, L.; Li, X.; Feng, H. Physiological characterization and comparative transcriptome analysis of a slow-growing reduced-thylakoid mutant of Chinese cabbage (Brassica campestris ssp. pekinensis). Front. Plant Sci. 2016, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.X.; Li, A.L. Transcriptome analysis of differentially expressed genes involved in proanthocyanidin accumulation in the rhizomes of Fagopyrum dibotrys and an irradiation-induced mutant. Front. Physiol. 2016, 18, 100. [Google Scholar] [CrossRef] [Green Version]
- Li, W.R.; Liu, C.F.; Liu, J.L.; Bai, Z.Q.; Liang, Z.S. Transcriptomic analysis reveals the GRAS family genes respond to gibberellin in Salvia miltiorrhiza hairy roots. BMC Genom. 2020, 27, 727. [Google Scholar] [CrossRef] [PubMed]
- Pei, T.L.; Ma, P.D.; Ding, K.; Liu, S.J.; Jia, Y.Y.; Ru, M.; Dong, J.E.; Liang, Z.S. SmJAZ8 acts as a core repressor regulating JA-induced biosynthesis of salvianolic acids and tanshinones in Salvia miltiorrhiza hairy roots. J. Exp. Bot. 2018, 69, 1663–1678. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Wang, X.X.; Cao, X.X.; Zhang, K.Q.; Sun, J.; Li, Y.H.; Sui, J.L.; Liu, Y.J. Integration of transcriptomics and metabolomics confirmed hepatoprotective effects of steamed shoot extracts of ginseng (Panax ginseng C.A. Meyer) on toxicity caused by overdosed acetaminophen. Biomed. Pharmacother. 2021, 143, 112177. [Google Scholar] [CrossRef]
- Bian, X.B.; Yan Zhao, Y.; Xiao, S.Y.; Yang, H.; Han, Y.Z.; Zhang, L.X. Metabolome and transcriptome analysis reveals the molecular profiles underlying the ginseng response to rusty root symptoms. BMC Plant Biol. 2021, 21, 215. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Liu, H.J.; Pan, J.Y.; Zhang, S.Y.; Guo, Y.X.; Xian, Y.S.; Sun, Z.R.; Zhang, Z.L. Transcriptomics and metabolomics changes triggered by inflorescence removal in Panax notoginseng (Burk.). Front. Plant Sci. 2021, 12, 761821. [Google Scholar] [CrossRef]
- Li, J.; Ma, L.; Zhang, S.T.; Zuo, C.L.; Song, N.; Zhu, S.S.; Wu, J.S. Transcriptome analysis of 1 and 3 year old Panax notoginseng roots and functional characterization of saponin biosynthetic genes DS and CYP716A47 like. Planta 2019, 249, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Song, J.; Luo, H.; Zhang, Y.; Li, Q.; Zhu, Y.; Xu, J.; Li, Y.; Song, C.; Wang, B.; et al. Analysis of the genome sequence of the medicinal plant Salvia miltiorrhiza. Mol. Plant 2016, 9, 949–952. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Cui, G.H.; Chen, T.; Ma, X.H.; Wang, R.S.; Jin, B.L.; Yang, J.; Kang, L.P.; Tang, J.F.; Lai, C.J.S.; et al. Expansion within the CYP71D subfamily drives the heterocyclization of tanshinones synthesis in Salvia miltiorrhiza. Nat. Commun. 2021, 12, 685. [Google Scholar] [CrossRef]
- Kang, S.H.; Pandey, R.P.; Lee, C.M.; Sim, J.S.; Jeong, J.T.; Choi, B.S.; Jung, M.H.; Ginzburg, D.; Zhao, K.M.; Won, S.Y.; et al. Genome-enabled discovery of anthraquinone biosynthesis in Senna tora. Nat. Commun. 2020, 11, 5875. [Google Scholar] [CrossRef]
- Mao, R.J.; Li, W.R.; He, Z.G.; Bai, Z.Q.; Xia, P.G.; Liang, Z.S.; Liu, Y. Physiological, transcriptional, and metabolic alterations in spaceflight-subjected Senna obtusifolia. Plant Physiol. Biochem. 2019, 139, 33–43. [Google Scholar] [CrossRef]
- Mao, R.J.; Qi, Z.H.; Han, R.L.; Liu, F.H.; Liu, Y.; Liang, Z.S. Spaceflight-induced variation on biological traits and effective components of Cassia obtusifolia. China J. Chin. Mater. Med. 2015, 40, 2571–2575. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of People’s Republic of China, Part 1; Chemical Industry Press: Beijing, China, 2015; Volume 145. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, S.; García-Gómez, J.M.; Terol, J.; Williams, T.D.; Nagaraj, S.H.; Nueda, M.J.; Robles, M.; Talón, M.; Dopazo, J.; Conesa, A. High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 2008, 36, 3420–3435. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids. Res. 2011, 39, 316–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, K.; Dong, Q.; Li, C.; Liu, C.; Ma, F. Genome wide identification and characterization of apple bHLH transcription factors and expression analysis in response to drought and salt stress. Front. Plant Sci. 2017, 8, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Huang, Q.; Wu, X.; Zhou, Z.W.; Ding, M.Q.; Shi, M.; Huang, F.F.; Li, S.; Wang, Y.; Kai, G.Y. Comprehensive transcriptome profling of Salvia miltiorrhiza for discovery of genes associated with the biosynthesis of tanshinones and phenolic acids. Sci. Rep. 2017, 7, 10554. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.P.; Wang, Y.J.; Silva, J.A.; Fan, Y.M.; Yu, X.N. Transcriptome and chemical analysis reveal putative genes involved in flower color change in Paeonia ‘Coral Sunset’. Plant Physiol. Biochem. 2019, 138, 130–139. [Google Scholar] [CrossRef]
- Biswal, B.; Jena, B.; Giri, A.K.; Acharya, L. De novo transcriptome and tissue specific expression analysis of genes associated with biosynthesis of secondary metabolites in Operculina turpethum (L.). Sci. Rep. 2021, 11, 22539. [Google Scholar] [CrossRef]
- Wu, Z.C.; Wang, W.; Sun, L.W.; Wei, A.Z.; Wang, D.M. Accumulation and biosynthesis of hydroxyl-α-sanshool in varieties of Zanthoxylum bungeanum Maxim. by HPLC-fingerprint and transcriptome analyses. Ind. Crops Prod. 2020, 145, 111998. [Google Scholar] [CrossRef]
- Jo, Y.D.; Ryu, J.H.; Kim, Y.S.; Kang, K.Y.; Hong, M.J.; Choi, H.I.; Lim, G.H.; Kim, J.B.; Kim, S.H. Dramatic increase in content of diverse flavonoids accompanied with down-regulation of F-Box genes in a Chrysanthemum (Chrysanthemum × morifolium (Ramat.) Hemsl.) mutant cultivar producing Dark-Purple ray florets. Genes 2020, 11, 865. [Google Scholar] [CrossRef]
- Lei, H.Y.; Niu, T.Z.; Song, H.F.; Bai, B.X.; Han, P.; Wang, Z.J.; Liu, A.K. Comparative transcriptome profiling reveals differentially expressed genes involved in flavonoid biosynthesis between biennial and triennial Sophora flavescens. Ind. Crop Prod. 2021, 161, 113217. [Google Scholar] [CrossRef]
- Zhang, J.Z.; Gao, W.Y.; Gao, Y.; Liu, D.L.; Huang, L.Q. Analysis of influences of spaceflight on chemical constituents in licorice by HPLC-ESI-MS/MS. Acta. Physiol. Plant. 2011, 33, 2511–2520. [Google Scholar] [CrossRef]
- Peng, L.; Ru, M.; Wang, Y.; Wang, K.R.; Li, B.; Xia, P.G.; Liang, Z.S. Spaceflight environment-induced variation in root yield and active constituents of Salvia miltiorrhiza. Planta Med. 2014, 80, 1029–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunsan, E.A.; Tukur, H.M.; Maigandi, S.A.; Makinde, A.A.; Malami, B.S.; Ogo, N.I. Utilization of Senna obtusifolia as a potential non-conventional feedstuff in sheep. Niger. Vet. J. 2011, 32, 199–203. [Google Scholar]
- Deng, Y.; Zheng, H.; Yan, Z.; Liao, D.; Li, C.; Zhou, J.Y.; Liao, H. Full-length transcriptome survey and expression analysis of Cassia obtusifolia to discover putative genes related to aurantio-obtusin biosynthesis, seed formation and development, and stress response. Int. J. Mol. Sci. 2018, 19, 2476. [Google Scholar] [CrossRef] [Green Version]
- Nam, J.; Seol, D.W.; Lee, C.G.; Wee, G.; Yang, S.; Pan, C.H. Obtusifolin, an anthraquinone extracted from Senna obtusifolia (L.) H.S.Irwin & Barneby, reduces inflammation in a mouse osteoarthritis model. Pharmaceuticals 2021, 14, 249. [Google Scholar] [CrossRef]
- Hou, J.Y.; Gu, Y.; Zhao, S.; Huo, M.Q.; Wang, S.F.; Zhang, Y.L.; Qiao, Y.J.; Li, X. Anti-inflammatory effects of aurantio-obtusin from seed of Cassia obtusifolia L. through modulation of the NF-κB pathway. Molecules 2018, 23, 3093. [Google Scholar] [CrossRef] [Green Version]
- Li, H.Y.; Liu, J.L.; Pei, T.L.; Bai, Z.Q.; Han, R.L.; Liang, Z.S. Overexpression of SmANS enhances anthocyanin accumulation and alters phenolic acids content in Salvia miltiorrhiza and Salvia miltiorrhiza Bge f. alba plantlets. Int. J. Mol. Sci. 2019, 20, 2225. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chen, J.F.; Zhou, X.; Chen, X.F.; Li, Q.; Tan, H.X.; Dong, X.; Xiao, Y.; Chen, L.D.; Chen, W.S. Dynamic metabolic and transcriptomic profiling of methyl jasmonate-treated hairy roots reveals synthetic characters and regulators of lignan biosynthesis in Isatis indigotica Fort. Plant Biotechnol. J. 2016, 14, 2217–2227. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Yuan, L.; Wu, B.; Li, X.; Chen, S.; Lu, S. Genome-wide identification and characterization of novel genes involved in terpenoid biosynthesis in Salvia miltiorrhiza. J. Exp. Bot. 2012, 63, 2809–2823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Wang, S.Q.; Wang, Z.Z. Genome-Wide Comprehensive Analysis the Molecular Phylogenetic Evaluation and Tissue-Specific Expression of SABATH Gene Family in Salvia miltiorrhiza. Genes 2017, 8, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.Z.; Jiang, M.D.; Xing, B.C.; Liang, L.J.; Zhang, B.X.; Liang, Z.S. Systematic Analysis of Kelch Repeat F-box (KFB) Protein Family and Identification of Phenolic Acid Regulation Members in Salvia miltiorrhiza Bunge. Genes 2020, 11, 557. [Google Scholar] [CrossRef]
- Guo, J.; Ma, X.; Cai, Y.; Ma, Y.; Zhan, Z.; Zhou, Y.J.; Liu, W.; Guan, M.; Yang, J.; Cui, G.; et al. Cytochrome p450 promiscuity leads to a bifurcating biosynthetic pathway for tanshinones. New Phytol. 2016, 210, 525–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgakov, V.P.; Tchernoded, G.K.; Mischenko, N.P.; Shkryl, Y.N.; Glazunov, V.P.; Fedoreyev, S.A.; Zhuravlev, Y.N. Effects of Ca2+ channel blockers and protein kinase/phosphatase inhibitors on growth and anthraquinone production in Rubia cordifolia callus cultures transformed by the rolB and rolC genes. Planta 2003, 217, 349–355. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/23387816 (accessed on 21 June 2020). [CrossRef]
- Reddy, R.R.N.; Harishbhai, M.R.; Harendrabhai, S.P.; Makasana, J.; Athamaram, N.G.; Ponnuchamy, M.; Kumar, J. Next generation sequencing and transcriptome analysis predicts biosynthetic pathway of sennosides from Senna (Cassia angustifolia Vahl.), a non-model plant with potent laxative properties. PLoS ONE 2015, 10, e0129422. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Clean Bases (GB) | Error Rate (%) | Q20 (%) | Q30 (%) | GC Content (%) |
|---|---|---|---|---|---|---|---|
| GC1 | 62,851,716 | 60,127,312 | 9.02 | 0.02 | 97.10 | 92.69 | 43.75 |
| GC2 | 61,381,444 | 58,745,742 | 8.81 | 0.02 | 97.29 | 93.06 | 43.64 |
| GC3 | 60,908,612 | 58,019,204 | 8.70 | 0.02 | 97.13 | 92.75 | 44.22 |
| QC10 | 73,498,576 | 71,952,222 | 10.79 | 0.02 | 96.58 | 91.40 | 43.82 |
| QC29 | 56,322,732 | 55,073,530 | 8.26 | 0.02 | 96.79 | 91.86 | 43.58 |
| QC46 | 61,461,010 | 59,680,544 | 8.95 | 0.02 | 95.87 | 89.94 | 44.18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, R.; Bai, Z.; Wu, J.; Han, R.; Zhang, X.; Chai, W.; Liang, Z. Transcriptome and HPLC Analysis Reveal the Regulatory Mechanisms of Aurantio-Obtusin in Space Environment-Induced Senna obtusifolia Lines. Int. J. Environ. Res. Public Health 2022, 19, 898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19020898
Mao R, Bai Z, Wu J, Han R, Zhang X, Chai W, Liang Z. Transcriptome and HPLC Analysis Reveal the Regulatory Mechanisms of Aurantio-Obtusin in Space Environment-Induced Senna obtusifolia Lines. International Journal of Environmental Research and Public Health. 2022; 19(2):898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19020898
Chicago/Turabian StyleMao, Renjun, Zhenqing Bai, Jiawen Wu, Ruilian Han, Xuemin Zhang, Weiguo Chai, and Zongsuo Liang. 2022. "Transcriptome and HPLC Analysis Reveal the Regulatory Mechanisms of Aurantio-Obtusin in Space Environment-Induced Senna obtusifolia Lines" International Journal of Environmental Research and Public Health 19, no. 2: 898. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19020898