
Salicylic Acid Manipulates Ion Accumulation and Distribution in Favor of Salinity Tolerance in Chenopodium quinoa


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Procedure
2.2. Treatments
2.3. Measurements
2.4. Data Analysis
3. Results
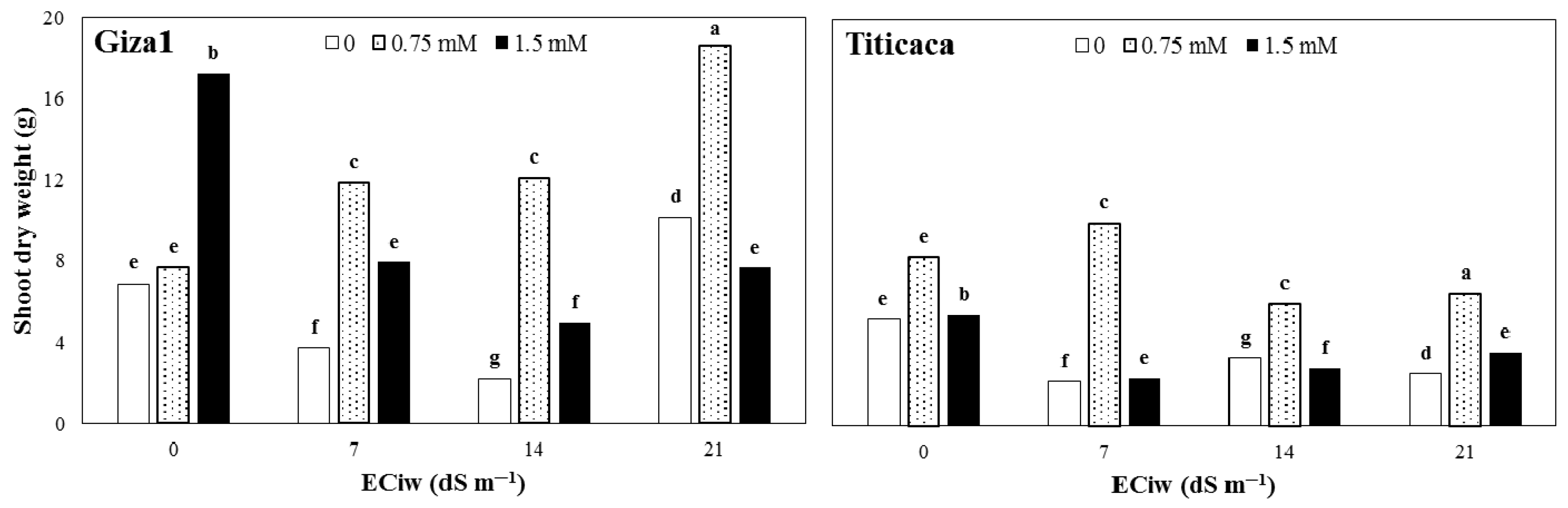
3.1. Shoot and Root Dry Weight
3.2. Ion Concentration in Shoot
3.3. Ion Concentration in Root
3.4. Ion Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Azooz, M.M.; Youssef, A.M.; Ahmad, P. Evaluation of salicylic acid (SA) application on growth, osmotic solutes and antioxidant enzyme activities on broad bean seedlings grown under diluted seawater. Int. J. Plant Physiol. Biochem. 2011, 3, 253–264. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Parnian, A.; Spasiano, D.; Race, M.; Ashraf, M. Haloculture: A system to mitigate the negative impacts of pandemics on the environment, society and economy, emphasizing COVID-19. Environ. Res. 2021, 198, 111228. [Google Scholar] [CrossRef] [PubMed]
- Parisi, A.; Monno, V.; Fidelibus, M.D. Cascading vulnerability scenarios in the management of groundwater depletion and salinization in semi-arid areas. Int. J. Disaster Risk Reduct. 2018, 30, 292–305. [Google Scholar] [CrossRef]
- Suarez, D.L.; Wood, J.D.; Lesch, S.M. Effect of SAR on water infiltration under a sequential rain–irrigation management system. Agric. Water Manag. 2006, 86, 150–164. [Google Scholar] [CrossRef]
- Khan, M.A.; Gul, B. Halophyte seed germination. In Ecophysiology of High Salinity Tolerant Plants; Khan, M.A., Weber, D.J., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 11–30. [Google Scholar] [CrossRef]
- Khan, M.A.; Ozturk, M.; Gul, B.; Ahmed, M.Z. Halophytes for Food Security in Dry Lands; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Manousaki, E.; Kalogerakis, N. Halophytes—An emerging trend in phytoremediation. Int. J. Phytoremediat. 2011, 13, 959–969. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Emam, Y.; Pessarakli, M. Grain filling pattern of Hordeum vulgare as affected by salicylic acid and salt stress. J. Plant Nutr. 2019, 42, 278–286. [Google Scholar] [CrossRef]
- Panda, A.; Rangani, J.; Kumari, A.; Parida, A.K. Efficient regulation of arsenic translocation to shoot tissue and modulation of phytochelatin levels and antioxidative defense system confers salinity and arsenic tolerance in the halophyte Suaeda maritima. Environ. Exp. Bot. 2017, 143, 149–171. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [Green Version]
- Volkov, V. Salinity tolerance in plants. Quantitative approach to ion transport starting from halophytes and stepping to genetic and protein engineering for manipulating ion fluxes. Front. Plant Sci. 2015, 6, 873. [Google Scholar] [CrossRef] [PubMed]
- Razzaghi, F.; Jacobsen, S.E.; Jensen, C.R.; Andersen, M.N. Ionic and photosynthetic homeostasis in quinoa challenged by salinity and drought–mechanisms of tolerance. Funct. Plant Biol. 2015, 42, 136–148. [Google Scholar] [CrossRef]
- Shabala, S.; Bose, J.; Hedrich, R. Salt bladders: Do they matter? Trends Plant Sci. 2014, 19, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Adolf, V.I.; Jacobsen, S.E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Emam, Y.; Hashemi, S.E.; Gaur, A. Role of chlormequat chloride and salicylic acid in improving cereal crops production under saline conditions. In Improving Cereal Productivity through Climate Smart Practices; Sareen, S., Sharma, P., Singh, C., Jasrotia, P., Singh, G.P., Sarial, A.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 145–158. [Google Scholar] [CrossRef]
- Abideen, Z.; Koyro, H.-W.; Huchzermeyer, B.; Ahmed, M.Z.; Gul, B.; Khan, M.A. Moderate salinity stimulates growth and photosynthesis of Phragmites karka by water relations and tissue specific ion regulation. Environ. Exp. Bot. 2014, 105, 70–76. [Google Scholar] [CrossRef]
- Pirasteh-Anosheh, H.; Emam, Y.; Rousta, M.J.; Ashraf, M. Salicylic acid induced salinity tolerance through manipulation of ion distribution rather than ion accumulation. J. Plant Growth Regul. 2017, 36, 227–239. [Google Scholar] [CrossRef]
- Cai, Z.Q.; Gao, Q. Comparative physiological and biochemical mechanisms of salt tolerance in five contrasting highland quinoa cultivars. BMC Plant Biol. 2020, 20, 70. [Google Scholar] [CrossRef] [Green Version]
- Hariadi, Y.; Marandon, K.; Tian, Y.; Jacobsen, S.E.; Shabala, S. Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 2011, 62, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, H.; Yaning, C.; Waqas, M.; Shareef, M.; Raza, S.T. Differential response of quinoa genotypes to drought and foliage-applied H2O2 in relation to oxidative damage, osmotic adjustment and antioxidant capacity. Ecotoxicol. Environ. Saf. 2018, 164, 344–354. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Aloisi, I.; Del Duca, S.; Canelo, V.; Torrigiani, P.; Silva, H.; Biondi, S. Salares versus coastal ecotypes of quinoa: Salinity responses in Chilean landraces from contrasting habitats. Plant Physiol. Biochem. 2016, 101, 1–13. [Google Scholar] [CrossRef]
- Shabala, S.; Hariadi, Y.; Jacobsen, S.E. Genotypic difference in salinity tolerance in quinoa is determined by differential control of xylem Na+ loading and stomatal density. J. Plant Physiol. 2013, 170, 906–914. [Google Scholar] [CrossRef] [PubMed]
- Pirasteh-Anosheh, H.; Emam, Y. Induced salinity tolerance and altered ion storage factor in Hordeum vulgare plants upon salicylic-acid priming. Iran Agric. Res. 2017, 36, 41–48. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops–what is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.A.; Zarebanadkouki, M.; Meunier, F.; Javaux, M.; Kaestner, A.; Carminati, A. Root type matters: Measurement of water uptake by seminal, crown, and lateral roots in maize. J. Exp. Bot. 2018, 69, 1199–1206. [Google Scholar] [CrossRef] [Green Version]
- Noreen, S.; Siddiq, A.; Hussain, K.; Ahmad, S.; Hasanuzzaman, M. Foliar application of salicylic acid with salinity stress on physiological and biochemical attributes of sunflower (Helianthus annuus L.) crop. Acta Sci. Pol.-Hortorum Cultus 2017, 16, 57–74. [Google Scholar] [CrossRef]
- Szepesi, Á. Influence of exogenous salicylic acid on antioxidant enzyme activities in the roots of salt stressed tomato plants. Acta Biol. Szeged. 2008, 52, 199–200. [Google Scholar]
- Szepesi, Á.; Csiszár, J.; Gémes, K.; Horváth, E.; Horváth, F.; Simon, M.L.; Tari, I. Salicylic acid improves acclimation to salt stress by stimulating abscisic aldehyde oxidase activity and abscisic acid accumulation, and increases Na+ content in leaves without toxicity symptoms in Solanum lycopersicum L. J. Plant Physiol. 2009, 166, 914–925. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Per, T.S.; Khan, N.A. Salicylic acid alleviates adverse effects of heat stress on photosynthesis through changes in proline production and ethylene formation. Plant Signal. Behav. 2013, 8, e26374. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, E.; Turan, M.; Guvenc, I. Effect of foliar salicylic acid applications on growth, chlorophyll, and mineral content of cucumber grown under salt stress. J. Plant Nutr. 2008, 31, 593–612. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is not all about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Bonales-Alatorre, E.; Shabala, S.; Chen, Z.H.; Pottosin, I. Reduced tonoplast fast-activating and slow-activating channel activity is essential for conferring salinity tolerance in a facultative halophyte, quinoa. Plant Physiol. 2013, 162, 940–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davenport, R.; James, R.A.; Zakrisson-Plogander, A.; Tester, M.; Munns, R. Control of sodium transport in durum wheat. Plant Physiol. 2005, 137, 807–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Cultivar | Salt Stress (dS m−1) | SA Level (mM) | Na+ | K+ | Cl─ | Ca2+ | Mg2+ | K/Na | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Giza1 | 0 | 0 | 0.72 | ±0.064 | 0.86 | ±0.070 | 0.63 | ±0.023 | 1.33 | ±0.062 | 0.74 | ±0.117 | 1.20 | ±0.094 |
| 0.75 | 0.39 | ±0.099 | 1.00 | ±0.062 | 0.42 | ±0.008 | 1.61 | ±0.096 | 0.98 | ±0.234 | 2.93 | ±0.766 | ||
| 1.5 | 0.35 | ±0.091 | 1.01 | ±0.064 | 0.40 | ±0.008 | 1.71 | ±0.099 | 0.92 | ±0.224 | 3.37 | ±0.911 | ||
| 7 | 0 | 1.38 | ±0.142 | 0.64 | ±0.073 | 0.90 | ±0.057 | 1.49 | ±0.095 | 1.13 | ±0.233 | 0.47 | ±0.054 | |
| 0.75 | 0.92 | ±0.092 | 0.73 | ±0.048 | 0.69 | ±0.038 | 1.70 | ±0.117 | 1.23 | ±0.309 | 0.81 | ±0.105 | ||
| 1.5 | 0.84 | ±0.094 | 0.73 | ±0.050 | 0.71 | ±0.037 | 1.73 | ±0.104 | 1.19 | ±0.293 | 0.90 | ±0.113 | ||
| 14 | 0 | 2.51 | ±0.108 | 0.42 | ±0.066 | 1.08 | ±0.077 | 1.16 | ±0.129 | 1.09 | ±0.376 | 0.17 | ±0.022 | |
| 0.75 | 1.92 | ±0.208 | 0.57 | ±0.079 | 0.90 | ±0.076 | 1.47 | ±0.157 | 1.01 | ±0.235 | 0.31 | ±0.057 | ||
| 1.5 | 1.89 | ±0.180 | 0.56 | ±0.082 | 0.89 | ±0.060 | 1.43 | ±0.162 | 0.97 | ±0.222 | 0.31 | ±0.052 | ||
| 21 | 0 | 2.66 | ±0.318 | 0.39 | ±0.059 | 1.29 | ±0.103 | 1.01 | ±0.082 | 0.68 | ±0.028 | 0.15 | ±0.021 | |
| 0.75 | 2.01 | ±0.156 | 0.50 | ±0.067 | 1.01 | ±0.082 | 1.35 | ±0.110 | 0.95 | ±0.185 | 0.26 | ±0.044 | ||
| 1.5 | 2.09 | ±0.163 | 0.50 | ±0.067 | 0.99 | ±0.076 | 1.21 | ±0.061 | 0.81 | ±0.071 | 0.24 | ±0.044 | ||
| Titicaca | 0 | 0 | 0.77 | ±0.038 | 0.83 | ±0.098 | 0.64 | ±0.014 | 1.35 | ±0.078 | 0.90 | ±0.161 | 1.09 | ±0.153 |
| 0.75 | 0.43 | ±0.069 | 0.96 | ±0.102 | 0.43 | ±0.011 | 1.62 | ±0.088 | 1.21 | ±0.250 | 2.31 | ±0.266 | ||
| 1.5 | 0.38 | ±0.065 | 0.96 | ±0.106 | 0.41 | ±0.011 | 1.76 | ±0.109 | 1.20 | ±0.291 | 2.62 | ±0.335 | ||
| 7 | 0 | 1.48 | ±0.090 | 0.61 | ±0.101 | 0.92 | ±0.038 | 1.53 | ±0.102 | 1.39 | ±0.271 | 0.42 | ±0.083 | |
| 0.75 | 1.01 | ±0.077 | 0.68 | ±0.082 | 0.71 | ±0.022 | 1.67 | ±0.107 | 1.50 | ±0.305 | 0.70 | ±0.137 | ||
| 1.5 | 0.92 | ±0.069 | 0.69 | ±0.084 | 0.72 | ±0.022 | 1.68 | ±0.104 | 1.44 | ±0.300 | 0.77 | ±0.148 | ||
| 14 | 0 | 2.66 | ±0.174 | 0.40 | ±0.090 | 1.10 | ±0.054 | 1.32 | ±0.280 | 1.38 | ±0.348 | 0.15 | ±0.035 | |
| 0.75 | 2.08 | ±0.194 | 0.53 | ±0.114 | 0.92 | ±0.056 | 1.49 | ±0.150 | 1.22 | ±0.233 | 0.26 | ±0.065 | ||
| 1.5 | 2.07 | ±0.179 | 0.52 | ±0.114 | 0.92 | ±0.037 | 1.65 | ±0.244 | 1.31 | ±0.324 | 0.26 | ±0.071 | ||
| 21 | 0 | 2.84 | ±0.193 | 0.36 | ±0.083 | 1.33 | ±0.066 | 1.12 | ±0.184 | 0.85 | ±0.187 | 0.13 | ±0.033 | |
| 0.75 | 2.21 | ±0.180 | 0.46 | ±0.097 | 1.05 | ±0.050 | 1.48 | ±0.173 | 1.22 | ±0.257 | 0.22 | ±0.061 | ||
| 1.5 | 2.30 | ±0.181 | 0.46 | ±0.097 | 1.03 | ±0.047 | 1.25 | ±0.098 | 0.98 | ±0.171 | 0.21 | ±0.058 | ||
| Cultivar | Salt Stress (dS m−1) | SA Level (mM) | Na+ | K+ | Cl− | Ca2+ | Mg2+ | K/Na | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Giza1 | 0 | 0 | 0.17 | ±0.009 | 1.01 | ±0.097 | 0.69 | ±0.057 | 1.29 | ±0.08 | 0.62 | ±0.03 | 5.87 | ±0.38 |
| 0.75 | 0.18 | ±0.010 | 1.06 | ±0.098 | 0.65 | ±0.041 | 1.26 | ±0.07 | 0.68 | ±0.03 | 5.71 | ±0.35 | ||
| 1.5 | 0.20 | ±0.011 | 1.10 | ±0.099 | 0.62 | ±0.032 | 1.19 | ±0.05 | 1.30 | ±0.03 | 5.48 | ±0.31 | ||
| 7 | 0 | 1.18 | ±0.145 | 2.21 | ±0.149 | 0.84 | ±0.060 | 1.61 | ±0.11 | 1.44 | ±0.06 | 1.90 | ±0.15 | |
| 0.75 | 1.58 | ±0.150 | 2.62 | ±0.176 | 0.96 | ±0.050 | 1.62 | ±0.08 | 2.07 | ±0.06 | 1.67 | ±0.09 | ||
| 1.5 | 2.14 | ±0.112 | 2.79 | ±0.187 | 1.35 | ±0.036 | 1.45 | ±0.09 | 1.75 | ±0.06 | 1.30 | ±0.03 | ||
| 14 | 0 | 1.71 | ±0.088 | 2.56 | ±0.172 | 1.31 | ±0.141 | 1.37 | ±0.06 | 1.33 | ±0.06 | 1.50 | ±0.05 | |
| 0.75 | 2.97 | ±0.206 | 3.01 | ±0.202 | 1.43 | ±0.075 | 1.36 | ±0.06 | 1.84 | ±0.06 | 1.01 | ±0.02 | ||
| 1.5 | 3.37 | ±0.191 | 2.90 | ±0.188 | 1.90 | ±0.090 | 1.21 | ±0.02 | 1.16 | ±0.05 | 0.86 | ±0.01 | ||
| 21 | 0 | 2.35 | ±0.218 | 3.08 | ±0.207 | 1.71 | ±0.270 | 0.88 | ±0.01 | 0.58 | ±0.04 | 1.32 | ±0.06 | |
| 0.75 | 3.58 | ±0.107 | 3.31 | ±0.223 | 2.03 | ±0.106 | 0.97 | ±0.03 | 1.31 | ±0.05 | 0.92 | ±0.04 | ||
| 1.5 | 4.16 | ±0.125 | 3.71 | ±0.149 | 2.35 | ±0.158 | 0.82 | ±0.01 | 0.94 | ±0.05 | 0.89 | ±0.04 | ||
| Titicaca | 0 | 0 | 0.34 | ±0.023 | 0.48 | ±0.033 | 0.74 | ±0.023 | 1.18 | ±0.072 | 0.424 | ±0.041 | 1.44 | ±0.003 |
| 0.75 | 0.33 | ±0.022 | 0.45 | ±0.030 | 0.77 | ±0.040 | 1.25 | ±0.067 | 0.532 | ±0.043 | 1.35 | ±0.005 | ||
| 1.5 | 0.35 | ±0.027 | 0.42 | ±0.028 | 0.83 | ±0.036 | 1.05 | ±0.067 | 0.639 | ±0.045 | 1.18 | ±0.014 | ||
| 7 | 0 | 1.31 | ±0.050 | 1.28 | ±0.086 | 0.91 | ±0.038 | 1.32 | ±0.055 | 0.456 | ±0.037 | 0.99 | ±0.097 | |
| 0.75 | 1.45 | ±0.030 | 0.78 | ±0.053 | 1.12 | ±0.059 | 1.41 | ±0.073 | 0.434 | ±0.032 | 0.54 | ±0.026 | ||
| 1.5 | 2.51 | ±0.101 | 0.54 | ±0.036 | 1.40 | ±0.111 | 1.19 | ±0.015 | 0.431 | ±0.036 | 0.22 | ±0.017 | ||
| 14 | 0 | 1.64 | ±0.042 | 2.15 | ±0.145 | 1.12 | ±0.067 | 1.12 | ±0.070 | 0.150 | ±0.037 | 1.32 | ±0.105 | |
| 0.75 | 2.11 | ±0.039 | 1.82 | ±0.122 | 1.71 | ±0.010 | 1.17 | ±0.077 | 0.825 | ±0.043 | 0.86 | ±0.043 | ||
| 1.5 | 3.57 | ±0.025 | 1.29 | ±0.087 | 2.07 | ±0.031 | 0.98 | ±0.021 | 0.635 | ±0.037 | 0.36 | ±0.024 | ||
| 21 | 0 | 2.09 | ±0.104 | 1.77 | ±0.119 | 1.53 | ±0.053 | 0.81 | ±0.020 | 0.107 | ±0.037 | 0.85 | ±0.071 | |
| 0.75 | 2.54 | ±0.051 | 1.89 | ±0.127 | 2.20 | ±0.054 | 0.87 | ±0.012 | 0.889 | ±0.043 | 0.75 | ±0.057 | ||
| 1.5 | 3.86 | ±0.078 | 1.28 | ±0.086 | 2.40 | ±0.062 | 0.73 | ±0.006 | 0.899 | ±0.045 | 0.33 | ±0.028 | ||
| Cultivar | Salt Stress (dS m−1) | SA Level (mM) | Na+ | K+ | Cl− | Ca2+ | Mg2+ | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Giza1 | 0 | 0 | 0.192 | ±0.006 | 0.540 | ±0.033 | 0.525 | ±0.012 | 0.491 | ±0.023 | 0.455 | ±0.045 |
| 0.75 | 0.319 | ±0.054 | 0.513 | ±0.032 | 0.609 | ±0.012 | 0.438 | ±0.015 | 0.410 | ±0.062 | ||
| 1.5 | 0.365 | ±0.059 | 0.521 | ±0.032 | 0.607 | ±0.010 | 0.410 | ±0.015 | 0.586 | ±0.062 | ||
| 7 | 0 | 0.461 | ±0.033 | 0.774 | ±0.011 | 0.481 | ±0.021 | 0.519 | ±0.020 | 0.561 | ±0.047 | |
| 0.75 | 0.633 | ±0.028 | 0.783 | ±0.011 | 0.580 | ±0.002 | 0.488 | ±0.016 | 0.626 | ±0.054 | ||
| 1.5 | 0.719 | ±0.013 | 0.792 | ±0.011 | 0.657 | ±0.006 | 0.456 | ±0.018 | 0.596 | ±0.056 | ||
| 14 | 0 | 0.405 | ±0.021 | 0.859 | ±0.013 | 0.548 | ±0.010 | 0.542 | ±0.036 | 0.549 | ±0.070 | |
| 0.75 | 0.608 | ±0.029 | 0.841 | ±0.013 | 0.614 | ±0.010 | 0.480 | ±0.024 | 0.644 | ±0.048 | ||
| 1.5 | 0.641 | ±0.024 | 0.838 | ±0.016 | 0.681 | ±0.008 | 0.459 | ±0.024 | 0.543 | ±0.052 | ||
| 21 | 0 | 0.469 | ±0.025 | 0.888 | ±0.011 | 0.571 | ±0.025 | 0.466 | ±0.022 | 0.461 | ±0.013 | |
| 0.75 | 0.640 | ±0.021 | 0.869 | ±0.012 | 0.667 | ±0.007 | 0.418 | ±0.024 | 0.579 | ±0.042 | ||
| 1.5 | 0.666 | ±0.022 | 0.882 | ±0.010 | 0.703 | ±0.021 | 0.403 | ±0.010 | 0.537 | ±0.019 | ||
| Titicaca | 0 | 0 | 0.305 | ±0.024 | 0.372 | ±0.037 | 0.538 | ±0.009 | 0.466 | ±0.027 | 0.331 | ±0.057 |
| 0.75 | 0.443 | ±0.054 | 0.321 | ±0.030 | 0.641 | ±0.016 | 0.436 | ±0.027 | 0.321 | ±0.061 | ||
| 1.5 | 0.488 | ±0.060 | 0.306 | ±0.029 | 0.669 | ±0.014 | 0.374 | ±0.030 | 0.368 | ±0.074 | ||
| 7 | 0 | 0.469 | ±0.007 | 0.679 | ±0.042 | 0.496 | ±0.021 | 0.463 | ±0.026 | 0.261 | ±0.052 | |
| 0.75 | 0.593 | ±0.023 | 0.537 | ±0.035 | 0.610 | ±0.018 | 0.457 | ±0.029 | 0.239 | ±0.049 | ||
| 1.5 | 0.733 | ±0.013 | 0.443 | ±0.036 | 0.656 | ±0.025 | 0.416 | ±0.016 | 0.246 | ±0.053 | ||
| 14 | 0 | 0.382 | ±0.016 | 0.846 | ±0.031 | 0.504 | ±0.013 | 0.469 | ±0.056 | 0.116 | ±0.048 | |
| 0.75 | 0.505 | ±0.027 | 0.777 | ±0.039 | 0.651 | ±0.014 | 0.442 | ±0.041 | 0.416 | ±0.057 | ||
| 1.5 | 0.635 | ±0.020 | 0.716 | ±0.047 | 0.694 | ±0.007 | 0.381 | ±0.041 | 0.348 | ±0.070 | ||
| 21 | 0 | 0.425 | ±0.021 | 0.832 | ±0.033 | 0.536 | ±0.004 | 0.428 | ±0.033 | 0.128 | ±0.051 | |
| 0.75 | 0.537 | ±0.017 | 0.806 | ±0.033 | 0.677 | ±0.016 | 0.375 | ±0.026 | 0.435 | ±0.062 | ||
| 1.5 | 0.628 | ±0.014 | 0.739 | ±0.043 | 0.701 | ±0.015 | 0.370 | ±0.016 | 0.486 | ±0.052 | ||
| Sodium | Potassium | K/Na Ratio | K/Na Selectivity | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | SF | Shoot | Root | SF | Shoot | Root | ||
| Shoot dry weight | −0.261 ns | −0.03 ns | 0.082 ns | 0.278 ns | 0.396 * | −0.531 ** | 0.433 * | 0.413 * | −0.159 ns |
| Root dry weight | −0.407 * | −0.217 ns | −0.138 ns | 0.372 ns | −0.097 ns | 0.038 ns | 0.426 * | 0.242 ns | 0.047 ns |
| Total dry weight | −0.405 * | −0.147 ns | 0.381 * | 0.335 ns | 0.410 * | −0.058 ns | 0.624 ** | 0.440 * | −0.095 ns |
| Chlorine | Calcium | Magnesium | |||||||
| Shoot | Root | SF | Shoot | Root | SF | Shoot | Root | SF | |
| Shoot dry weight | −0.265 ns | −0.067 ns | 0.403 * | 0.172 ns | 0.081 ns | −0.062 ns | −0.485 ** | 0.473 * | 0.526 ** |
| Root dry weight | −0.352 ns | −0.120 ns | 0.220 ns | 0.131 ns | −0.094 ns | −0.208 ns | −0.149 ns | −0.023 ns | 0.075 ns |
| Total dry weight | −0.225 ns | −0.113 ns | 0.452 * | 0.221 ns | 0.052 ns | −0.144 ns | −0.402 * | 0.409 * | 0.444 * |
| Variable Entered | Partial R2 | Model R2 | F Value | Pr > F | |
|---|---|---|---|---|---|
| Total dry weight | MgSF | 0.703 | 0.703 | 52.28 | 0.001 |
| Shoot Mg2+ | 0.190 | 0.893 | 32.17 | 0.003 | |
| Shoot K/Na | 0.050 | 0.943 | 16.50 | 0.022 | |
| Root K/Na | 0.030 | 0.973 | 11.90 | 0.012 | |
| ClSF | 0.019 | 0.992 | 9.25 | 0.043 | |
| Root dry weight | Root K/Na | 0.882 | 0.882 | 66.67 | 0.001 |
| Shoot Na+ | 0.092 | 0.974 | 20.91 | 0.011 | |
| NaSF | 0.021 | 0.995 | 10.75 | 0.022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohammadi, H.; Rahimpour, B.; Pirasteh-Anosheh, H.; Race, M. Salicylic Acid Manipulates Ion Accumulation and Distribution in Favor of Salinity Tolerance in Chenopodium quinoa. Int. J. Environ. Res. Public Health 2022, 19, 1576. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19031576
Mohammadi H, Rahimpour B, Pirasteh-Anosheh H, Race M. Salicylic Acid Manipulates Ion Accumulation and Distribution in Favor of Salinity Tolerance in Chenopodium quinoa. International Journal of Environmental Research and Public Health. 2022; 19(3):1576. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19031576
Chicago/Turabian StyleMohammadi, Hamid, Bahareh Rahimpour, Hadi Pirasteh-Anosheh, and Marco Race. 2022. "Salicylic Acid Manipulates Ion Accumulation and Distribution in Favor of Salinity Tolerance in Chenopodium quinoa" International Journal of Environmental Research and Public Health 19, no. 3: 1576. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19031576