We characterized white-tailed deer spatial distribution and visitation in relation to ‘4-Poster’ baited treatment stations to understand the influence of baited stations on deer movement and quantify deer use of tick treatment stations. Our study analyzed ‘4-Poster’ usage across a period of time and in ways that have not been studied in the past. Overall, we found evidence of small shifts in deer distributions over fine temporal scales in response to feeder stations being refilled with corn. We demonstrated small shifts towards feeding stations, alongside a complementary decrease in distance between collared deer. Presumably, those findings led to the small increase in the density of deer, or at least deer activity, we detected in the region around the ‘4-Poster’ feeders. In general, we found that collared deer visit feeders reasonably frequently throughout the year, with most individuals visiting more than one feeder within a park. Though both sexes primarily used feeders around crepuscular hours, we did document clear temporal differences in the probability of feeder visits. Interestingly, we detected changes in deer space use despite lacking sufficient samples from some age classes and using a 15 m proxy distance to feeders as a ‘visitation’. Overall, these findings give us a better understanding of both the effects of ‘4-Poster’ feeders on deer behavior and distribution, as well as on how deer use these feeders, pointing to clear management concerns and recommendations.
4.1. Impacts of Feeder Stations on Deer Distribution
Our findings on the impact of active ‘4-Poster’ feeders were generally consistent with past research. Though the literature reports that access to supplemental feed has had variable effects on white-tailed deer movements and spatial distribution, with studies generally reporting modest impacts, most have demonstrated that range size is likely to decrease in the presence of supplemental food [
21,
25,
27,
28,
49]. We saw evidence that the distribution of hourly locations and proximity to conspecifics changed with the availability of corn in ‘4-Poster’ feeders on a fine temporal scale. Deer hourly locations were slightly closer to ‘4-Poster’ feeders and other collared deer during the 3 days following feeder refilling. However, this finding did not extend to the 95% OD shrinking after feeders were serviced, possibly due to decreased statistical power from the smaller sample size. However, our 50% OD was found closer to feeder stations following the refill. This finding was supported by past research which has shown that the distance from core areas to supplemental feed areas can shift 4.1–115 m closer once food is available [
21,
22,
26], comparable to the average shift of ~35 m we documented. It is possible that corn was available at the feeders during a portion of the pre-servicing period, violating our assumption that feeders were empty. If corn was available during the pre-servicing period, this may cause deer to be closer to feeders, weakening our comparison between before and after periods. Similarly, the corn supply could have been completely depleted during the 3-day post-servicing period, but this is less likely as feeders were regularly stocked with 100–200 lbs of corn each service period.
Many studies have found that supplemental feed is unlikely to draw in more deer from outside locations, but if the feed is within an established range, then it can increase recruitment and compact ranges, leading to higher densities of deer [
26,
27]. Though our analysis did not look at recruitment, the documented shift in ranges towards feeders could lead to modestly elevated local densities. This is shown more directly in our analysis of deer clustering, where we documented collared deer being closer together after feeders were refilled, presumably in response to multiple deer shifting their ranges in the same direction.
Overall, our results support local deer populations shifting their ranges towards filled feeders, concentrating deer at point sources of food and closer to conspecifics. Although these shifts are of fairly small magnitudes, clustering and increasing shared space among deer can have negative consequences to managing the spread of diseases such as CWD or bovine tuberculosis, which have important implications for humans [
50,
51]. It is impossible to know about feeder use by uncollared deer without monitoring stations with camera traps. While individual deer may partition times of feeder visits and avoid direct contact with other deer, disease agents such as the prions of CWD are environmentally persistent in feces, urine, and saliva, allowing deer to become passively infected. Informed decisions on the appropriate use of ‘4-Poster’ feeders in specific areas will depend on current knowledge of deer densities, tick densities, disease prevalence, and community needs and concerns. Regardless, managers must critically balance the conflicting outcomes of ‘4-Poster’ use, which include local tick control and the likelihood of concentrating local deer herds.
4.2. Use of ‘4-Poster’ Feeders by Deer
Some studies have suggested deploying ‘4-Poster’ feeders or supplemental feeding stations at one station per 50–60 ha to reduce deer using multiple bait piles while still effectively covering the area [
21,
26]. Based on recent literature reviews, ‘4-Poster’ densities may need to be much higher to reach the population size needed to achieve effective tick control [
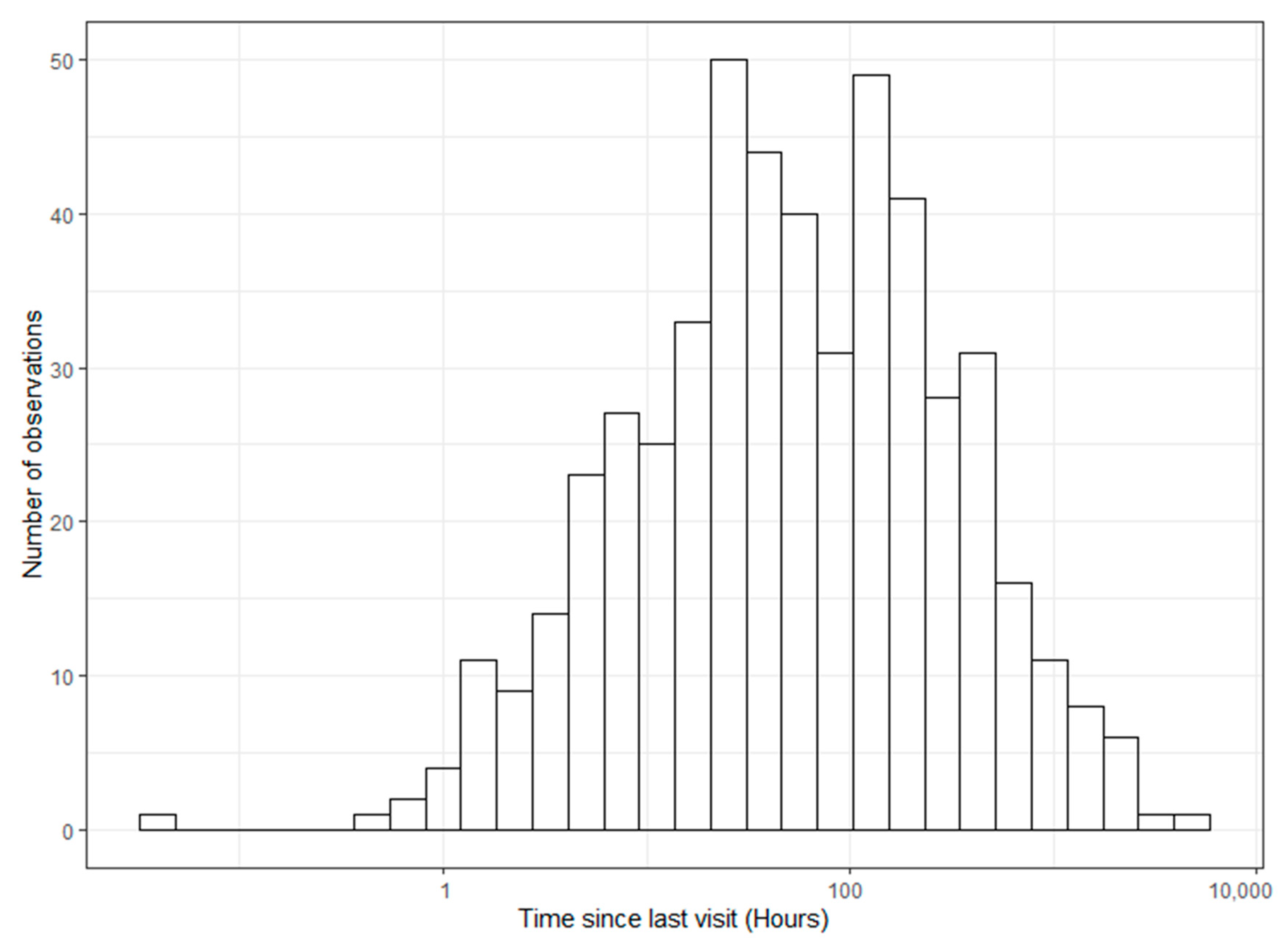
20]. Our densities were approximately one feeder per 15–19 ha, and we saw 83% of collared deer using feeders. We also documented the widespread use of multiple feeders by the same individual deer, and time between visits was highly variable among individuals and difficult to predict using sex or day of the year. Some individuals would consistently use specific feeders for months at a time, whereas others would visit only a few times in the same time period.
In terms of tick control, other studies have shown that the treatment of 50–70% of the deer population will result in a 60% reduction in infected nymphs after five years [
52,
53]. However, Tickicide
® is permethrin-based, which kills ticks on contact but does immediately degrade in the environment. Depending on exposure to light and precipitation, permethrin has a half-life averaging 39 days in the soil, but ranges from 1 to 113 days [
54,
55,
56]. Given our visitation rates, to maintain consistent continuous treatment, deer may have to visit active feeders a minimum of once every 2 weeks in instances when new questing ticks are picked up after the last treatment of Tickcide
® has degraded naturally.
There are a number of factors that could influence deer use of feeding stations. Natural forage availability can influence feeder use [
13,
57]. Both sexes exhibited slightly lower probabilities of feeder visits during spring and fall compared to the rest of the year, possibly due to changes in food resource availability such as spring green-up or mast production in the fall. Additionally, personal preference and personality between individuals may have a large influence on which individuals use feeding stations and the frequency of use [
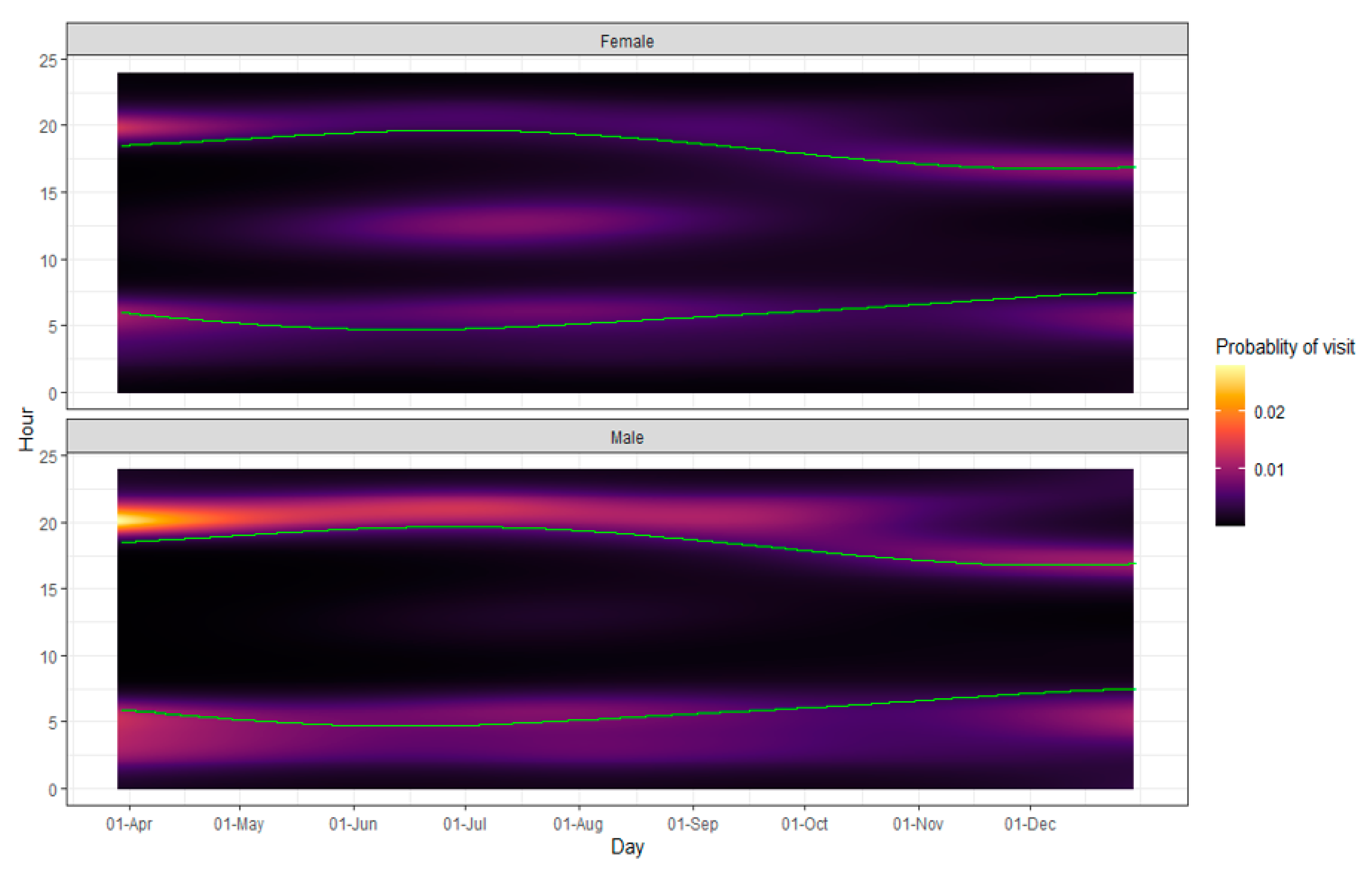
25]. Overall, we found that males used a higher percentage of available feeders and visited them more throughout the deployment periods (
Figure 3). Though we demonstrate that both sexes showed an increased probability in feeder use during crepuscular hours, Bartoskewitz et al. (2003) had also documented males exploited supplemental feed more heavily than females.
Social hierarchy has been shown to influence which deer have access to feeders [
30,
58]. Males and particularly older males will have dominance over resources, but they prefer to use supplemental feed after daylight hours, which may drive females to use feeders more often during the day. Consistent with this premise, we document a mid-day spike in feeder visits by females during the summer months (
Figure 3). As such, it does not appear that social dominance by males alone prevents the use of ‘4-Poster’ feeders by females but may be one influence that shifts females’ use outside of the crepuscular hours, at least during summer. Unfortunately, we could not model the effect of age on feeder use due to the lack of sample size for certain ages, though age and dominance hierarchies can affect which deer have access to feed [
29], and we hypothesized increasing age would increase dominance of use.
As days shortened, mid-day feeder use declined, and may have contributed to the decreased ability of females to access ‘4-Poster’ feeders. Unfortunately, the decrease in female mid-day visits September through October overlaps with peak activity season for adult black-legged ticks. Furthermore, rutting activity, which occurs primarily from 15 October to 15 November in this area, may disrupt feeder usage, as we see an increase in deer speed and activity coinciding with a lower probability of crepuscular ‘4-Poster’ visits during this time [
59,
60]. As a result, we recommend keeping ‘4-Poster’ feeders active until the end of December to allow more access for deer as the probability of use begins to increase again (
Figure 3).
Many past studies only looked at baiting impacts for short periods within a single year such as hunting season or winter supplemental feeding. However, Jerina (2012) postulated that the length and history of supplemental feeding may be an important factor because it takes several years of continuous baiting to observe clear responses in home range size. Feeders for this study were first deployed in 2017, but there was a legacy of baiting and ‘4-Poster’ feeder use by County Park personnel for management activities in previous years, which may have made the local deer population more inclined to use supplemental feed. Deer use of ‘4-Poster’ stations may have been intensified because we were filling them on a timeline that allowed feeders to be emptied between feeder service dates. Thompson et al. (2008) found that deer use of feeders was more intense for rationed feed versus unlimited amounts, and our ‘4-Poster’ feeders were often found empty during most feeder service dates, creating a timeline of empty–full–empty bait availability. Based on past work and as 83.3% of our collared deer visited a feeder within a 3-year period and revisited feeders an average of every five days, we recognize our protocol may provide adequate coverage for tick control. When deemed necessary, we recommend keeping ‘4-Poster’ feeders active continuously throughout tick seasons for multiple years to achieve the best results in tick reductions. Unfortunately, in many regions, active tick seasons can occur year-round depending on climate and tick species present. Therefore, we recommend strategically restricting access at certain times (e.g., snow cover, air temp < 1.67 °C) to decrease costs, reduce pressure on immediate habitat, and intensify deer use of feeders once refilled.







{kind=link}
{kind=link}
{kind=link}