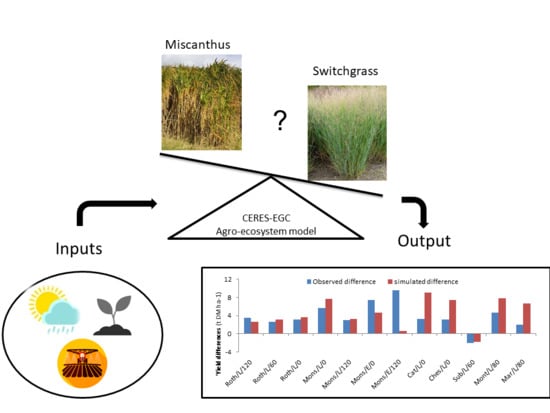
The following paragraphs describe the adaptation of CERES-EGC to the miscanthus and switchgrass crops. Most of the modeling concepts and equations apply to miscanthus, and the switchgrass version was evolved from the miscanthus routines by adjusting the relevant parameters, according to the literature on the ecophysiology of this particular crop.
2.1.3. Crop Development Stages
For miscanthus and switchgrass, five development stages were defined with a base temperature of 6 °C for miscanthus and 10 °C for switchgrass. The developmental stages of miscanthus are described below. They also apply to switchgrass.
Stage 1: Shoot emergence. This stage requires a minimum air temperature above 10 °C and daylengths longer than 12 hours.
Stage 2: Leaf growth. Leaves start growing after a thermal time of 900 GDD6 (Growing Degree Day with a base temperature of 6 °C for miscanthus and 10 °C for switchgrass) has elapsed after emergence. The crop leaf area index (LAI) may increase up to a maximum value of 7.5.
Stage 3: Onset of leaf senescence. Crop LAI diminishes at a daily rate of 0.03 m
2 m
−2, which doubles when the plant enters its overall senescence (stage 4). This occurs after a thermal time of 3000 GDD
6 has elapsed. Photosynthesis continues during Stage 3, with all the photosynthates being allocated to rhizome. This stage may end prematurely if there are six consecutive days with an air temperature under 10 °C, a frost or 30 consecutive drought days [
32]. At the end of this stage, leaves are assumed to have been entirely shed.
Stage 4: Onset of plant senescence. Photosynthesis stops and part of the stem biomass is translocated to the rhizomes. Stems dry up and are gradually shed until the day of harvest.
Stage 5: Dormancy. Plant growth stops until new shoots emerge.
2.1.4. Biomass Production, Partitioning, and Translocation
Plant growth and biomass partitioning. At the beginning of the simulation (stand establishment phase), miscanthus or switchgrass plants are only made up of their rhizomes and roots. Potential dry matter production is calculated through a radiation use efficiency (
RUE) calculation. Potential dry matter production is based on light interception, as follows:
where
PCARB is the potential aboveground dry matter production of the day (g DM m
−2),
PAR are the photosynthetically active radiations (MJ d
−1), and
k the extinction coefficient.
The daily increase in plant dry matter (
CARBO, g DM m
−2) is calculated as:
where
SWDF and
NDEF are 0–1 modifiers accounting for limitations through water and N stress (see corresponding section below), and
REMC (g DM m
−2) is a flux of biomass remobilized from rhizome to the aboveground parts of the plant. During the first two development stages, 45% of the miscanthus rhizome dry matter is transferred to aboveground [
21]. A conversion efficiency of gross to net energy (or biomass) flow of 50% is assumed [
21].
Daily plant growth is the sum of above- and below-ground biomass increments. Photosynthates are partitioned between the various plant compartments according to crop development stage, using partitioning variables for each stage and each compartment of the plant (
Table A1). Before the stand reaches maturity, more photosynthates are allocated to below-ground compartments to ensure crop establishment. When maturity is reached (phenological Stage 2), all photosynthates are apportioned to aboveground biomass pools.
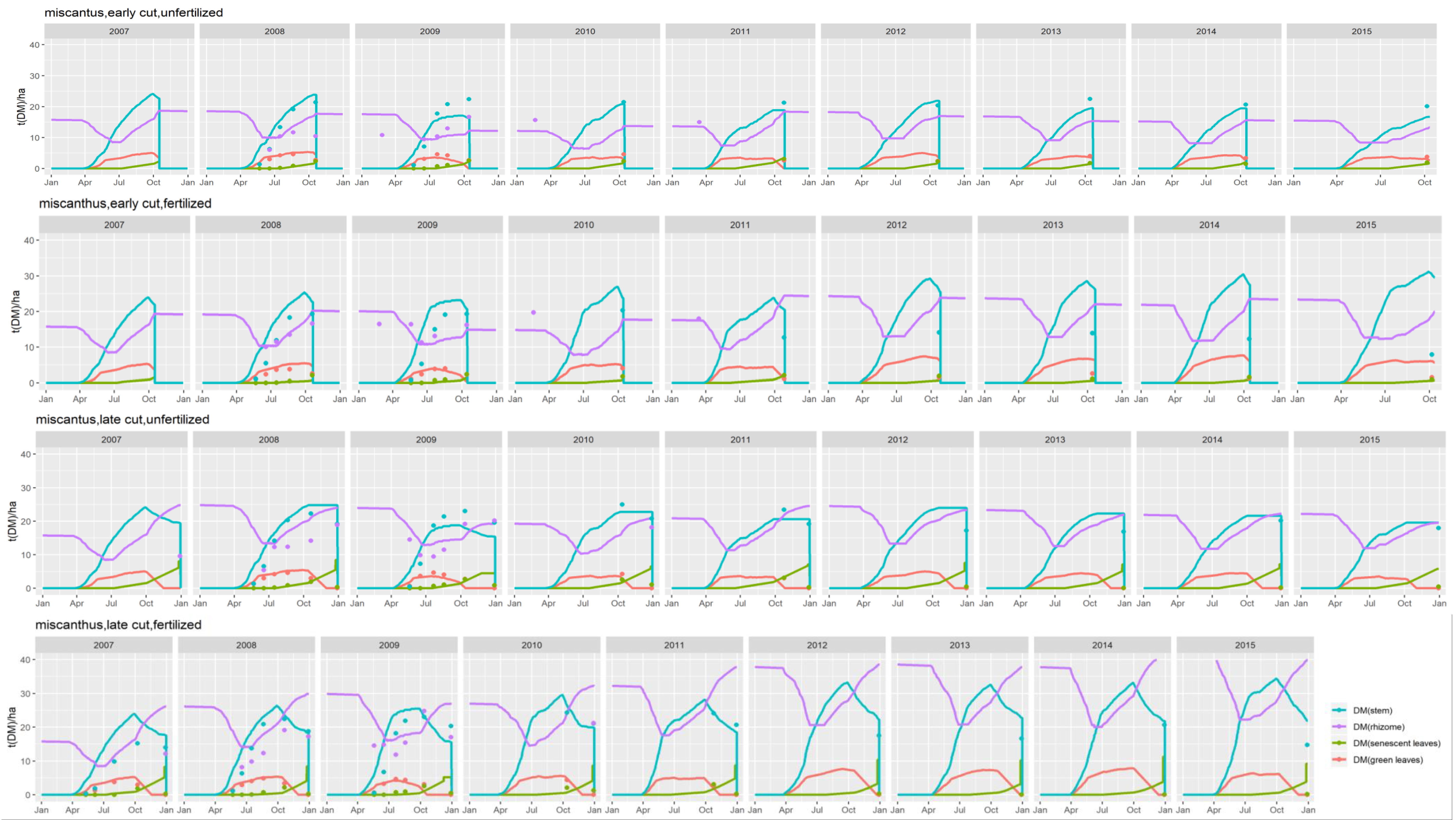
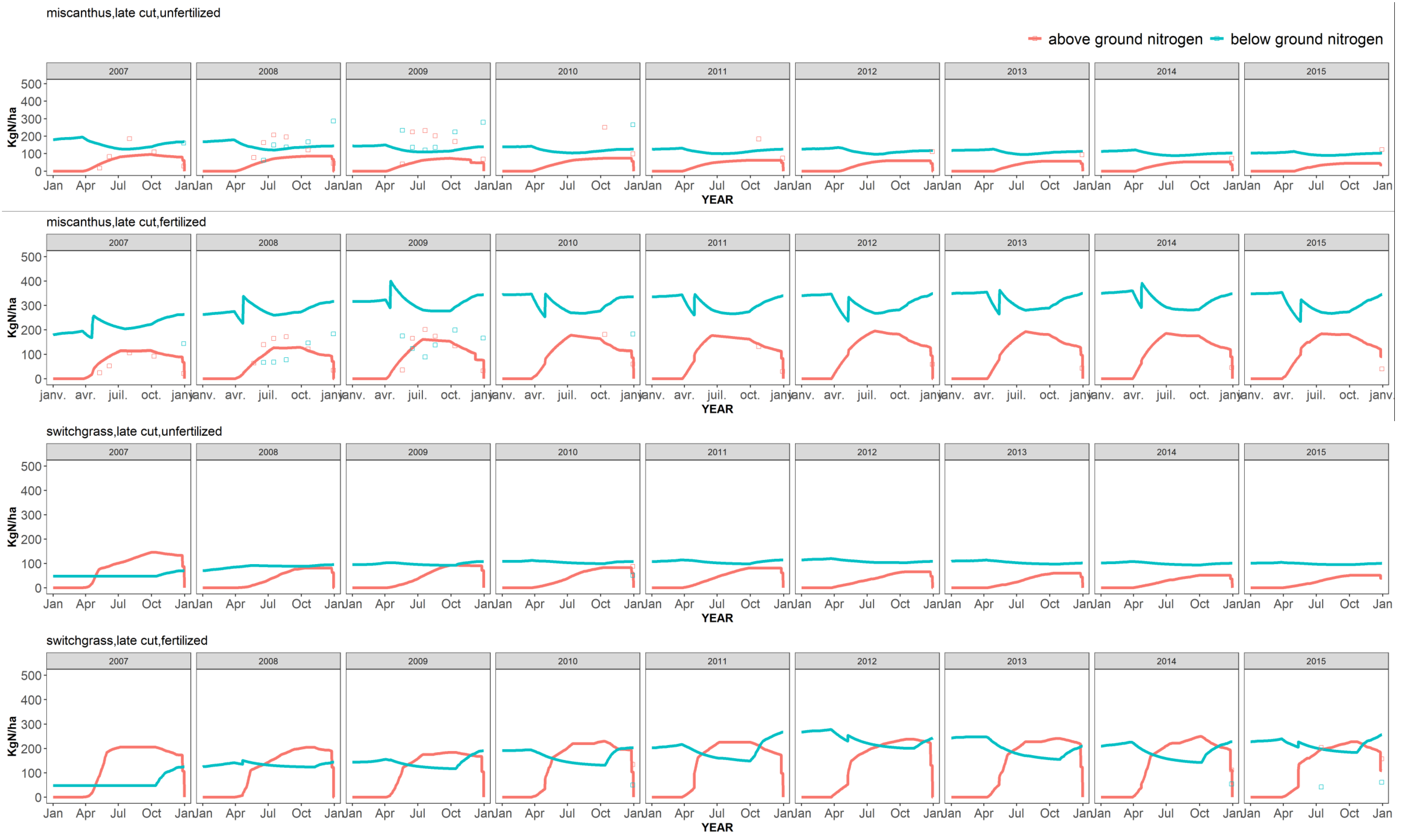
At the end of the first year, miscanthus or switchgrass stems are usually not harvested but cut and left on the soil surface. In the following years, the CERES-EGC model calculates biomass yields as the weight of plant stems and leaves, assuming 10% are left as stubbles and hence returned to the soil.
Leaf development. During leaf growth (phenological Stage 2),
LAI expands at a relative rate of 0.018 m
2 m
−2 d
−1 for miscanthus [
21], provided enough dry matter is available to maintain a constant specific leaf area (SLA). Leaf expansion is reduced accordingly if this is not the case. Leaf expansion is also modulated by leaf-specific nutrient and water stress modifiers (see corresponding section below). During leaf senescence
LAI declines at a relative rate of 0.03 m² m
−2 d
−1 [
21].
Roots. The root growth subroutine is adapted from the original CERES routine [
30]. The initial root length density is set to 0.08 cm root cm
−3 soil. The vertical elongation rate of roots is a function of thermal time, soil water and N contents. Root density may increase to a maximum value of 6 cm root cm
−3 soil in the top 10 cm of soil, 3 cm root cm
−3 soil in the 10–30 cm layer, and 1 cm root cm
−3 soil below the 30 cm depth. Maximum rooting depth is set to 3 m.
Plant senescence. Root and rhizome senescence occur throughout the growing season with daily decay rates of 0.05% and 0.001%, respectively. Rhizome starts senescing only after a threshold mass value of 500 g DM m−2 has been reached for this compartment. Senescent leaves that have dropped at the end of Stage 3, stems left on the ground after harvest, senescent roots and rhizomes are returned to the soil as fresh organic matter pools.
N and C fluxes within the plant. At the beginning of the growing season, a pool of rhizome N that can be potentially remobilized to the aerial parts (
REMN, in g N m
−2) is calculated as:
where
RHNIN is the rhizome nitrogen content in kg DM ha
−1.
Every day, rhizomes may contribute up to a maximum of
REMN to the N demand, or until rhizome N has reached a minimum concentration of 0.6%, to prevent its total N depletion. Potential crop N uptake is a function of the amount of nitrate and ammonium available in each soil layer, and the corresponding root density [
30]. From leaf senescence to harvest, if N uptake is larger than plant demand, excess N is stored in the rhizome up to a maximum concentration of about 2% [
33].
During leaf and plant senescence, N is remobilized from the aboveground biomass to the rhizome. The daily N remobilization rates (denoted
RHN, in g N m
−2 d
−1) depend on growing degree-days relative to the duration of the whole period of remobilization. They are calculated as [
11]:
where
LFNIN is the initial N contents of leaves (stems) at the beginning of leaf (stem) senescence, respectively (in g N m
−2).
P4-AP3 is the thermal time from the end of leaf senescence to plant senescence. N remobilization from the stems is supposed to occur between October and February.
P8 is the thermal time from October to February. Stems and leaves keep a structural N concentration around 0.3% (on a mass basis [
10]), and the remobilization is limited by a maximum rhizome N content of 2%.
Water and N stress. Similar to most crop models, the effect of N deficiencies on crop growth is calculated via a 0 to 1 modifier (stress factor) corresponding to a supply-to-demand ratio. Crop N demand is based on the concept of critical N concentration in plant biomass, i.e., the optimal concentration for biomass production depending on plant development stage. The definition of such concentrations for miscanthus is complicated by rhizome remobilization, and there are no data specific to this plant. We therefore used a generic allometric relationship for the aboveground biomass of annual and perennial crops:
where
Ncrit% is the critical N concentration (%, mass basis),
W is the aboveground biomass (g DM m
−2), and
a and
b are parameters set at 3.0 and −0.47 for both crops. An upper limit of 4% was set for
Ncrit% for low
W values, while root N concentration was set to 0.6%. [
10].
Following the generic approach of the CERES models [
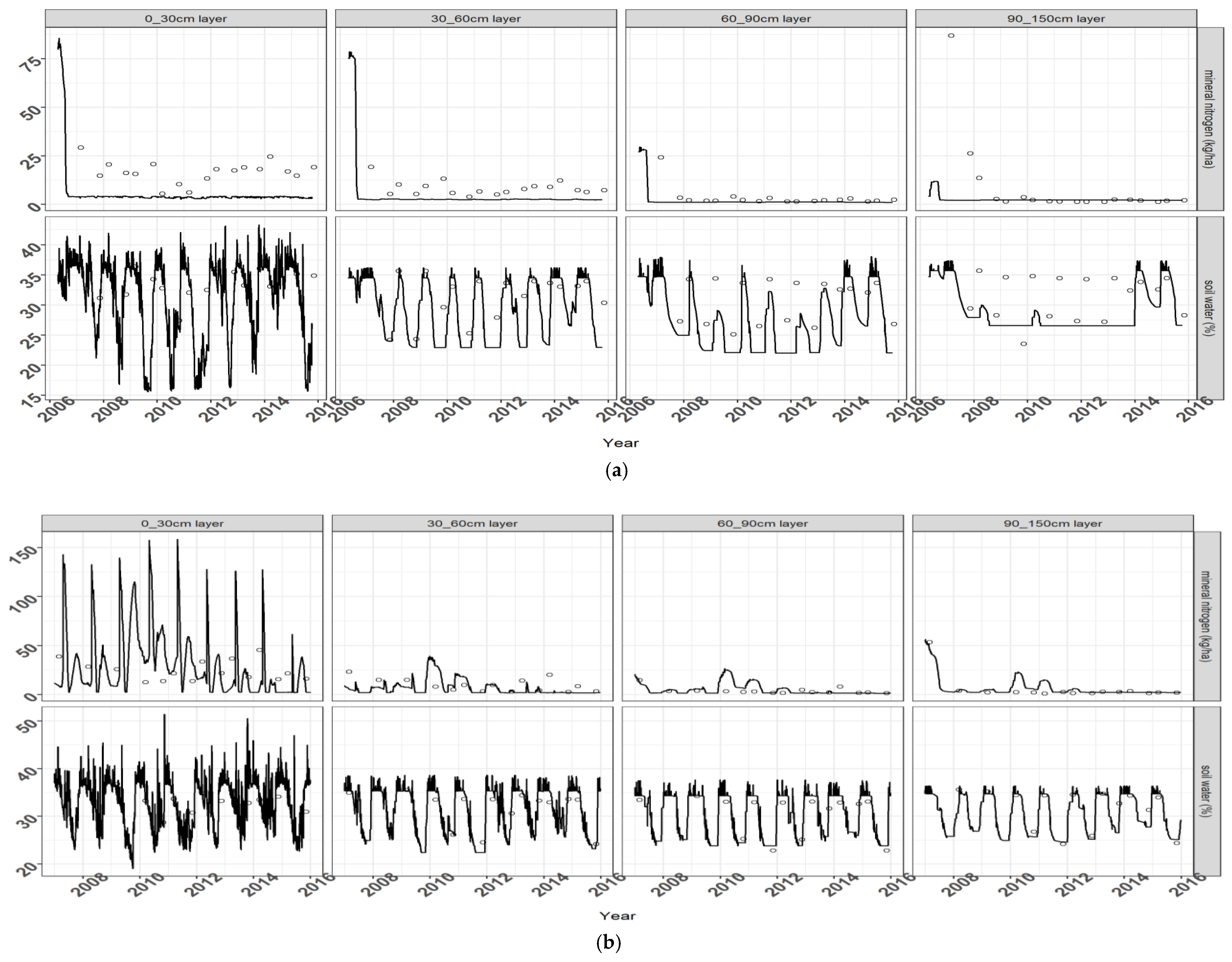
30], the water stress effects on plant photosynthesis and development depend on the ratio of actual to potential plant transpiration, as modulated by an unitless parameter (WSTRSS) accounting for the particular sensitivity of miscanthus to water stress. Maximum plant transpiration is calculated from a Penman potential evapotranspiration rate (which is an input for the model), with a crop coefficient ranging between 0.8 and 1.1 depending on crop leaf area index. Root water uptake depends on soil available water and root density in each soil layer.
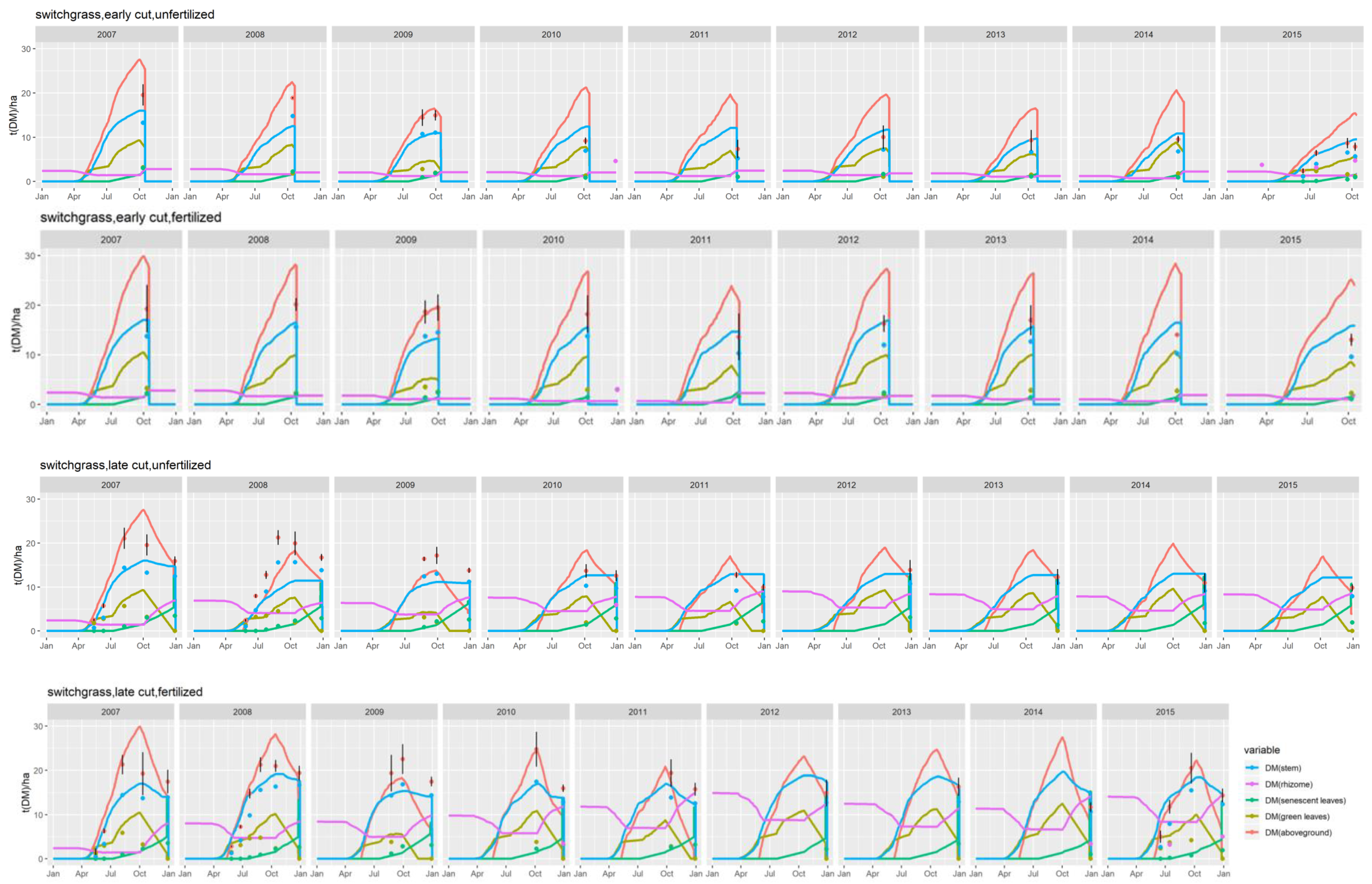
Adaptation to switchgrass. Except for the difference in base temperatures, all the above equations and concepts apply equally to miscanthus and switchgrass. For lack of similar literature and modeling concepts pertaining to switchgrass, the latter crop was modeled with the same equations, whether for crop development stages, photosynthesis or
LAI development. The simulation of drought kill gave one exceptional major difference between the two crops. On any given year miscanthus was considered to fail if plant available water remained at zero for more than one month at a time (without compromising next year’s harvest), and the crop was terminated if this condition extended for more than two months [
32]. This condition was deactivated for switchgrass, for which no such effects were reported, resulting in a higher tolerance to water stress for this crop.
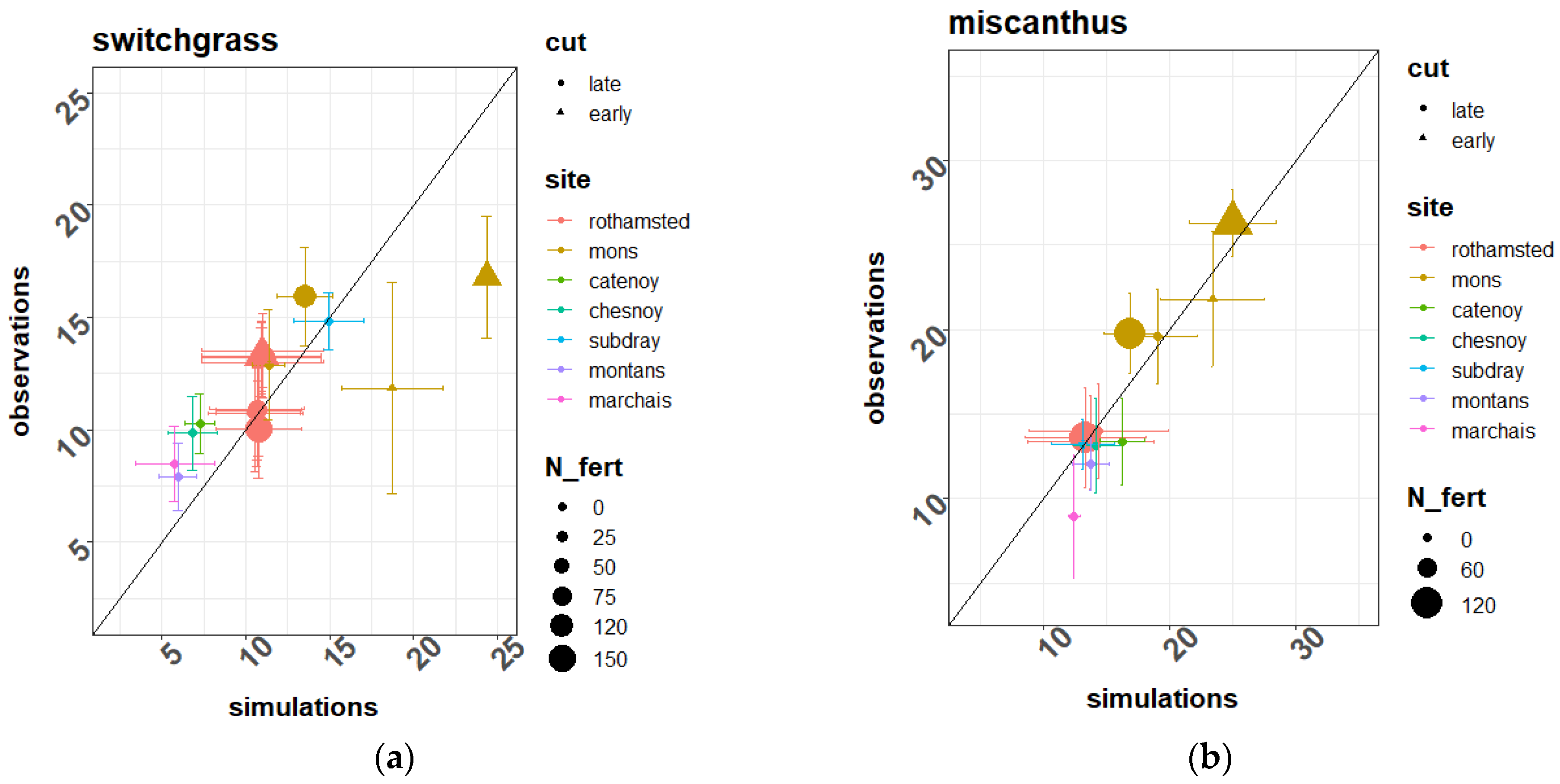
The parameters used to simulate the two crops on the Estrées-Mons site were mostly extracted from the literature, as can be seen in the
Appendix A,
Table A1.
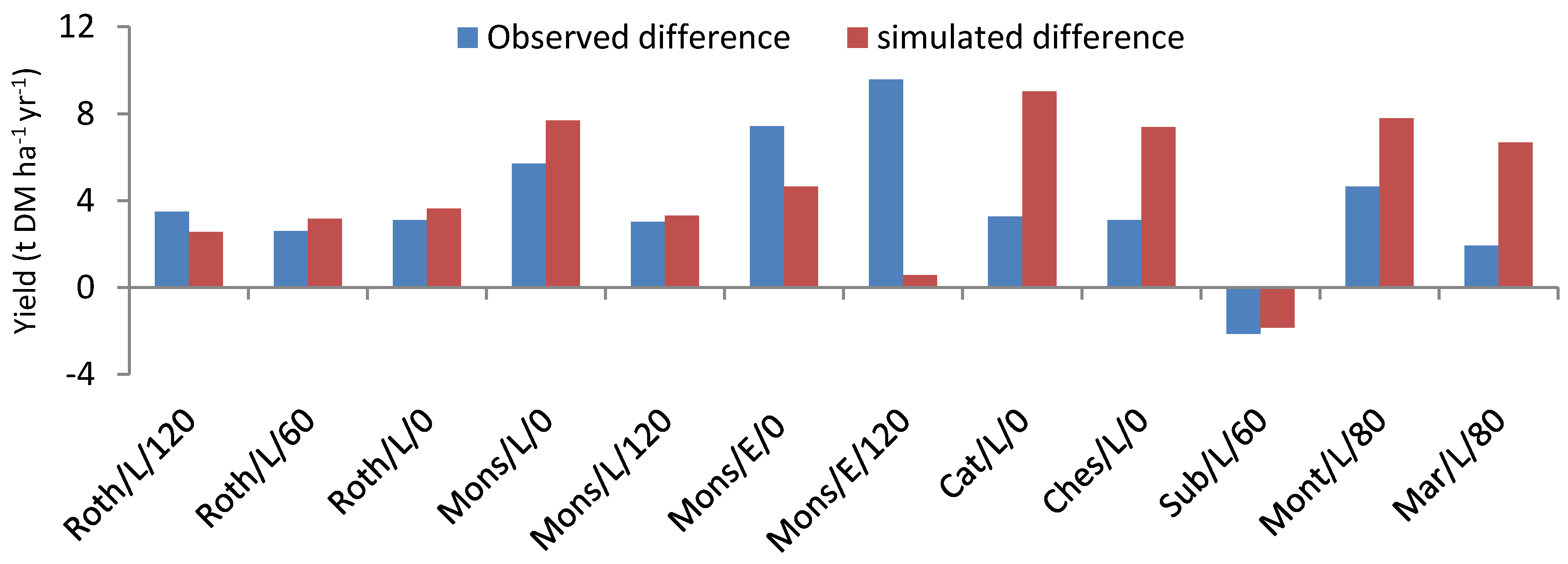
Some of them (especially for switchgrass) were calibrated by fitting the model against the experimental data of this site. Such was the case for the radiation use efficiency, for example (
Table A1). The selection of parameters undergoing such calibration (done by trial and error and for a single parameter at a time) was based on a preliminary sensitivity analysis of the times series of simulated biomass and crop N content.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





