
Improvement of Photoautotrophic Algal Biomass Production after Interrupted CO2 Supply by Urea and KH2PO4 Injection


Department of Chemical and Biological Engineering, Korea University, 145 Anam-ro, Seongbuk-gu, Seoul 02841, Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Energies 2021, 14(3), 778; https://0-doi-org.brum.beds.ac.uk/10.3390/en14030778
Submission received: 31 December 2020
/
Revised: 28 January 2021
/
Accepted: 31 January 2021
/
Published: 2 February 2021
(This article belongs to the Section A4: Bio-Energy)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Microalgae-derived biomass is currently considered a sustainable feedstock for making biofuels, including biodiesel and direct combustion fuel. The photoautotrophic cultivation of microalgae using flue gas from power plants has been continuously investigated to improve the economic feasibility of microalgae processes. The utilization of waste CO2 from power plants is advantageous in reducing carbon footprints and the cost of carbon sources. Nonetheless, the sudden interruption of CO2 supply during microalgal cultivation leads to a severe reduction in biomass productivity. Herein, chemical fertilizers including urea and KH2PO4 were added to the culture medium when CO2 supply was halted. Urea (5 mM) and KH2PO4 (5 mM) were present in the culture medium in the form of CO2/NH4+ and K+/H2PO4−, respectively, preventing cell growth inhibition. The culture with urea and KH2PO4 supplementation exhibited 10.02-fold higher and 7.28-fold higher biomass and lipid productivity, respectively, compared to the culture with ambient CO2 supply due to the maintenance of a stable pH and dissolved inorganic carbon in the medium. In the mass cultivation of microalgae using flue gas from coal-fired power plants, urea and KH2PO4 were supplied while the flue gas supply was shut off. Consequently, the microalgae were grown successfully without cell death.
1. Introduction
The emissions of carbon dioxide (CO2) from the use of fossil fuels are increasingly raising the atmospheric levels of CO2. This increase in the levels of CO2 in the atmosphere is associated with climate change [1], rising sea levels [2] and direct risks to human health [3]. The Paris Agreement was adopted at the 2015 United Nations (UN) Conference on Climate Change to foster cooperation between countries in reducing anthropological CO2 emissions. Subsequently, interest in carbon capture, utilization, and storage (CCUS) technologies has increased rapidly [4,5,6]. In particular, biological conversion technologies that capture CO2 using only photosynthetic organisms and light as energy sources are promising technologies [7,8]. Bio-based CO2 conversion processes also have a high potential as a sustainable carbon reduction technology, producing valuable products while simultaneously mitigating CO2 emissions [9,10,11,12].
Microalgae can be utilized as a representative bio-platform for CO2 conversion thanks to their advantages over traditional biomass resources, including: (1) high photosynthetic efficiency; (2) high biomass productivity; (3) no controversy in food and feed ethics; (4) low water consumption for cultivation; (5) availability in non-arable areas [13,14,15,16,17]. It is estimated that there are approximately 20,000 to 800,000 microalgal species on Earth, with around 50,000 having been discovered so far [18]. It is possible to produce hundreds of useful substances, such as biofuels [19,20,21], bioplastics [22,23], antioxidants [24,25,26], and pigments [27,28], by selecting the appropriate microalgal species, taking into consideration the culture conditions and target products based on the diversity of species. Taking into account their potential for CO2 mitigation and the large size of this market, the demand for microalgae-based biofuels is constantly increasing [29]. Despite the advantages of using microalgae as a feedstock for biofuels, large-scale microalgae processes have yet to be adopted commercially because of the high cost of production [30]. Thus, the main obstacles to cost-effective biofuel production are: (1) low net volumetric lipid productivity [31]; (2) high input energy for harvesting processes [32]; (3) high cost for obtaining CO2, nutrients, and water [33]. To overcome these drawbacks and improve the economic feasibility of microalgae processes, numerous efforts have been made to increase the efficiency in both the upstream and downstream processes [34,35,36,37,38,39,40]. In particular, culture systems able to directly utilize waste CO2 emitted from power plants has attracted attention for its potential to capture large amounts of carbon, thereby reducing the cost of carbon sources [41,42].
Depending on the type of fuel used in a given power plant, flue gas contains approximately 2–20% of CO2 [43,44]. These CO2 concentrations can be directly applied to microalgae cultivation with suitable microalgal species and culture systems. The direct use of flue gas can minimize the cost of carbon sources, which is a significant factor that determines the overall cost of biological processes [45]. In outdoor large-scale cultivation of microalgae, if artificial CO2 and organic carbon sources are used as carbon sources, the total cost of the carbon source is very high, but there is no cost when using flue gas emitted from coal-fired power plants. Also, the establishment of microalgae processes near the flue gas emission source can be used to reduce the cost and energy of transporting carbon sources further [46]. Despite these advantages, flue gas supplies can be temporarily interrupted at any time due to their dependence on equipment and machines that are prone to failure. To minimize the use of external energy, the outdoor cultivation of microalgae utilizing sunlight as the sole energy source requires a culture period of 1 to 4 weeks [47,48]. If the CO2 injection is halted during the night (no light) the pH will not change considerably as compared to the daytime (light). However, the supply of CO2 is not only interrupted at night but depending on the situation of the power plant such as emergency stop and over-haul thereby causing pH change in unexpected times. Thus, if the supply of flue gas containing a high concentration of CO2 is halted during this period, a rapid change in the pH of the culture medium occurs, resulting in a decrease in cell growth or cell death [49]. Cultivating microalgae after cell death is expensive and time consuming, due to the need to wash photobioreactors (PBR), creating a culture medium, and preparing microalgal seeds [50]. Thus, the overall cost of the process can be very large.
In this study, we propose a strategy to prevent cell death and improve biomass and lipid production during flue gas supply interruption in microalgal cultivation. Urea and KH2PO4, which are widely used as chemical fertilizers, were introduced into the culture medium to provide additional nutrient sources and alleviate rapid changes in pH. The optimum concentrations of urea and KH2PO4 were investigated to maximize the biomass production. In addition, the possibility of a synergistic effect on increasing biomass productivity was confirmed by injecting the two chemicals simultaneously. To analyze these results, the dissolved inorganic carbon (DIC) concentration and the pH of the medium were analyzed in a chemical fertilizer system and compared with conventional buffer systems supplied with high concentrations of CO2. Additionally, we verified whether the improvement of cell growth in photoautotrophic culture could lead to an improvement in the lipid productivity. Lastly, the system was applied to a large-scale microalgal cultivation process using flue gas emitted from a coal-fired power plant. In conclusion, upon the interruption of the CO2 supply, which can lead to substantial losses in the large-scale process, it was possible to suppress the cell death caused by a rapid change in the medium pH and enhance the biomass productivity.
2. Materials and Methods
2.1. Algal Strains and Culture Conditions
Chlorella sorokiniana UTEX 2714 was acquired from the Culture Collection of Algae at the University of Texas at Austin (TX, USA). In the indoor cultivation, all cells were cultured in a 500-mL graduated cylinder containing 0.05 g L−1 of CaCl2, 0.1 g L−1 of MgSO4, 0.4 g L−1 of NH4Cl, 0.108 g L−1 of K2HPO4, 0.056 g L−1 of KH2PO4, and 1 mL of Hutner’s trace elements solution with 10 mM of potassium hydroxide solution (KOH). This KOH acts as bicarbonate buffer when CO2 is bubbled in the culture. 10 mM KOH solution was aerated with 100% CO2 gas and flue gas from a coal-fired power plant for lab-scale cultivation and outdoor cultivation, respectively [9]. To keep the pH constant in bicarbonate buffer system, CO2 must be continuously supplied. If the bicarbonate buffer system is collapsed, the rapid increase in the pH(up to 9.5) of the culture medium occurs, resulting in a decrease in cell growth or cell death. CO2 was supplied from the bottom by a sparger to create a bubble column for cultivation and preventing sedimentation. Inorganic or organic compounds, including 5 mM of urea and KH2PO4 were also added simultaneously at the interruption of the CO2 supply. For the experiment comparing of different buffer systems, 20 mM of 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) was supplemented instead of KOH. A two-stage culture strategy was applied to enhance the synchronous production of biomass and lipids under photoautotrophic culture conditions. In the growth stage, the cells were fully grown under light of 150 μE m−2 s−1. In the induction stage, the cells were transferred to a nitrogen-depleted medium for inductive growth and lipid production with intense illumination (300 μE m−2 s−1). For the lab-scale experiments, the artificial light was illuminated using LED and the natural sunlight was used at outdoor cultivation.
2.2. Analytical Methods
2.2.1. Analysis of Algal Cell Growth
Biomass concentration was determined by measuring the dry cell weight (DCW) using pre-dried (at 105 °C in a drying oven) and pre-weighed GF/F glass microfiber filter (Whatman, Cambridge, UK). The microfiber filter was washed with deionized (DI) water before use. The sample was centrifuged and washed with DI water. Then, a 5-mL aliquot of evenly mixed sample was uniformly loaded onto the pre-weighed dry filter. The cell-loaded filter was washed using DI water to remove any excess salt in the sample. The filter was dried in an oven for 2 days to evaporate the DI water. Finally, dry cell weight was determined as the difference between the mass of the sample-loaded filter and the pre-weighed dry filter [45].
2.2.2. Determination of pH and Dissolved Inorganic Carbon in the Medium
To determine the pH and dissolved inorganic carbon (DIC) concentration (mg L−1) of the medium, the evenly mixed sample was centrifuged at 3000 rpm and 23 °C for 10 min to obtain a supernatant. The pH of the supernatant was measured using a digital pH meter (HI 8314 model; Hanna Instruments, Seoul, Republic of Korea). The dissolved inorganic carbon was titrated against standardized hydrochloric acid (HCl, 0.03 N) via phenolphthalein titration. The supernatant (20 mL) was placed in a 50-mL conical tube attached to a pH meter and titrated against HCl using phenolphthalein (pKa = 8.6) indicator to know the first end point of colorless. And then, methyl orange (pKa = 3.8) as the indicator was used to know the second end point. The first end point corresponds to OH− + CO2−3 (total alkalinity), while the second corresponds to OH− + HCO−3 (total acidity). The DIC content was estimated by subtracting the second end point from the first and applying the volumetric relation V1N1 = V2N2 [51]. V1 and N1 are the supernatant volume and DIC concentration (mg/L) of samples. V2 is the subtracted value and N2 is the known concentration of the HCl.
2.2.3. Lipid Extraction and Lipid Content of Algal Cells
At the end of the growth stage (after 4 days), the cells were centrifuged and re-suspended in nitrogen-depleted medium for inductive growth and lipid production with intense illumination (300 μE m−2 s−1). After 6 days of nitrogen starvation, the lipid-accumulated cell suspension was obtained from the graduated cylinder to measure the lipid content of the microalgal cells. Total lipid was first extracted using the modified Bligh and Dyer method for the total lipid assay of C.sorokiniana [52]. Briefly, 3 mL of each culture sample in a 15-mL screw-capped glass tube was centrifuged at 3000 rpm and 23 °C for 10 min. The resulting supernatant was discarded to obtain the cell pellet. Then, the cell pellet was homogeneously mixed with 3.9 mL of a chloroform and methanol mixture (1:2, v/v) utilizing vortexing, and incubated at 50 °C in a heating block. The extraction was repeated until the cell debris became colorless. The cell debris was removed by centrifugation at 3000 rpm and 23 °C for 10 min. 1.3 mL of chloroform and 2.0 mL of DI water were supplemented to the supernatant. Then the samples were centrifuged at 3000 rpm and 23 °C for 10 min to obtain the organic phases. 2.0 mL of the organic phase was loaded to a pre-weighed dry glass tube. The sample-loaded glass tube was dried overnight at 50 °C to remove the organic solvent. Finally, amount of lipids was determined as the difference between the weight of the sample-loaded glass tube and the pre-weighed dry glass tube [53].
2.3. Outdoor Cultivation of Microalgae Using Flue Gas from Coal-Fired Power Plant
To protect the microalgal cultivation system from external hazards, a polycarbonate (PC) greenhouse (width × length × height: 5 m × 10 m × 3.7 m) was installed at the Taean coal power plant (PREMIUM DESIGN, Chungcheongnam-do, Republic of Korea) [44]. The microalgal cells were inoculated into a 10-L thin-film photobioreactor (PBR) constructed with a polymer film (polyethylene terephthalate, octene, and Nylon-6) [54]. The cells were cultivated in the PBR with 0.1 vvm of the flue gas by the ring blower connected to the stack of the power plant. To simulate the interruption of the flue gas supply, after 4 days of cultivation, the flue gas supply line was blocked, and the air was injected using a bypass line. Then, 5 mM of urea and 5 mM of KH2PO4 were injected into the PBR in the form of a solution. Natural sunlight was provided as the only light source.
3. Results and Discussion
3.1. Introduction of Urea and KH2PO4 to Microalgae Culture System for Preventing Cell Growth Inhibition from Interruption in CO2 Supply
Microalgae are photosynthetic organisms that can be photoautotrophically cultivated to reduce carbon source costs and carbon footprint. HEPES and Tris buffer systems are widely used to maintain stable pH in lab-scale photoautotrophic microalgal cultivation. However, the applications of large-scale culture systems are limited by to their high cost [48]. Accordingly, Choi et al. developed an affordable bicarbonate buffer system applicable to large-scale microalgae processes by utilizing flue gas emitted from power plants [9]. To generate the bicarbonate buffer, air containing a high concentration of CO2 is injected into an alkaline solution to decrease the pH and increase the DIC concentration. The overall reaction can be abbreviated as shown in Equations (1) and (2) [19]:
H2O + CO2 → H2CO3 ↔ H+ + HCO3−
HCO3− ↔ H+ + CO32−
Therefore, when photoautotrophically culturing the microalgae using the bicarbonate buffer system, it is essential to constantly supply a high concentration of CO2. In particular, outdoor cultivation using flue gas from a coal-fired power plant is optimized for this system because of the high concentration of CO2 in the flue gas (13–15%). Despite these advantages, an emergency shutdown or the inspection of a power plant can lead to interruptions in the supply of CO2 for hours to weeks during microalgae cultivation. To minimize losses in this situation, a strategy by which to maintain the neutral pH of the culture medium and supply a carbon source to the microalgal cells is required (Figure 1). If the supply of flue gas is stopped, the pH of the culture medium changes rapidly as the bicarbonate buffer system collapses, which can induce cell death. One solution involves injecting artificially produced CO2 by mixing it with air. However, this approach is also limited by its high cost. Due to its high nitrogen content (46.7%), urea is considered a low-cost and efficient form of nitrogen compared to other nitrogen sources [55]. Since urea can be dissolved in the form of CO2 and NH3 in water with the help of urease from microalgal cells, it represents both a carbon source and a nitrogen source to microalgal cells [56]. The overall reaction can be abbreviated as shown in Equations (3) and (4) [55,56].
CO(NH2)2 + H2O → CO2 + 2NH3
NH3 + H2O → NH4+ + OH−
In addition, urea has been widely used as an efficient alternative to nitrogen sources due to its low production cost and low price per kilogram [57].
Phosphate is a necessary nutrient for cellular material, containing phospholipids, nucleic acids and nucleotides. It also provides a significant function in cellular processes, including signal transduction, energy transfer, macromolecule biosynthesis, photosynthesis, and respiration. Polyphosphates plays a main role in the phosphorus storage in microalgae, composed of unbranched chains of PO4− structural units linked together by oxygen bridges. The concentration of polyphosphate present in the microalgae is dependent on the phosphate available in the medium, the degree of available light, and the temperature. A phosphate buffer system has buffer properties because of the following ionizations reactions, as shown in Equations (5) and (6). Buffer systems using KH2PO4 play a key role in maintaining the pH at moderate conditions during cultivation:
KH2PO4− ↔ K+ + H2PO42− (pKa = 6.8)
H2PO42− ↔ HPO42−
Here, we present a strategy that can be applied to microalgal culture systems during an interruption in the CO2 supply. Injecting urea and KH2PO4, which are relatively inexpensive chemical fertilizers, into the microalgal culture solution whose gas supply has stopped, allows the culture to maintain a stable pH and provides sufficient nutrient sources to suppress cell death and increase biomass productivity.
3.2. Effect of Urea and KH2PO4 on Microalgal Biomass Production
3.2.1. Effect of Urea Concentration on Biomass Production
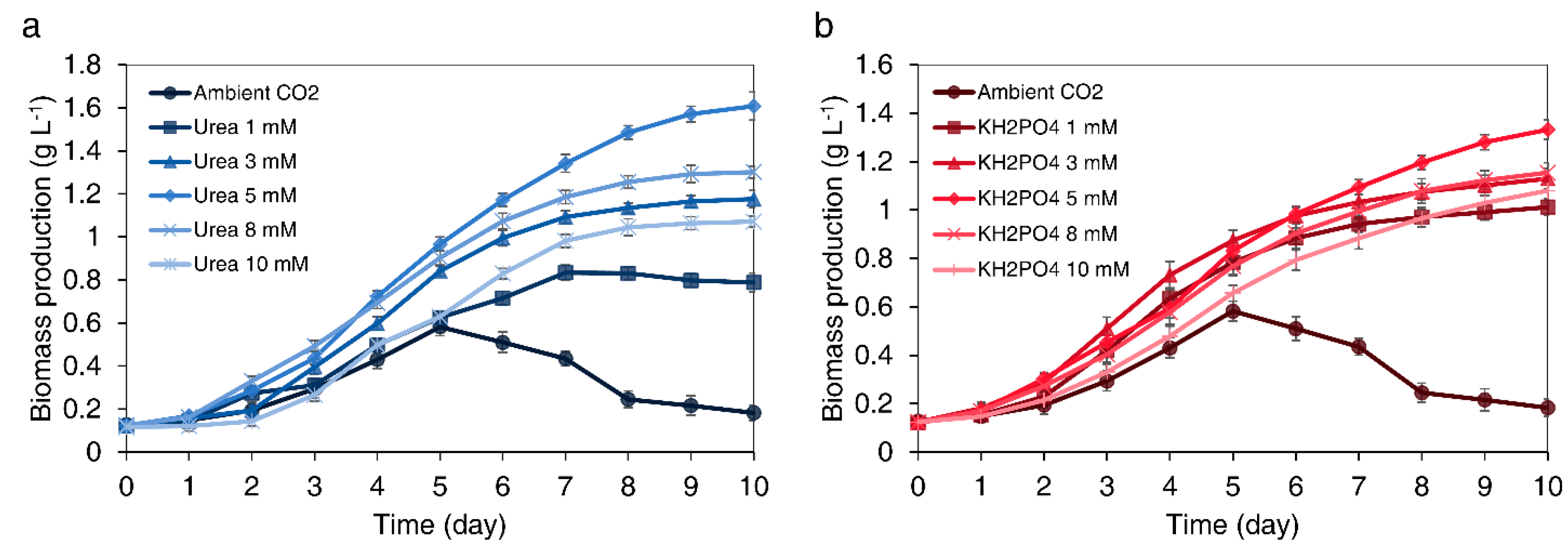
Both ammonia-based chemical fertilizers and urea accumulate ammonia in water in a unionized form. However, in contrast to other ammonia-based chemical fertilizers, urea releases CO2. Therefore, urea can provide microalgal cells a source of carbon when the supply of CO2 is interrupted. To confirm whether cells can grow with a supply of urea alone, biomass production was analyzed according to the concentration of urea injected under ambient CO2 supply conditions. As shown in Figure 2a, C. sorokiniana cells were grown with 0, 1, 3, 5, 8, and 10 mM urea. The biomass concentration increased up to 5 mM as the urea concentration increased. However, higher urea concentrations resulted in biomass loss compared to 5 mM. This is consistent with the results reported by Goswami and Kalita, who found that the excessive supply of urea to microalgal cells interferes with cell growth [58]. As a result, cultures supplied 5 mM of urea exhibited a 8.94-fold higher biomass production (1.61 g L−1) compared to cultures supplied only ambient CO2 without urea (0.18 g L−1) (Figure 2a). The biomass productivity was 0.161 g L−1 day−1 for the cultures supplied with 5 mM of urea and 0.018 g L−1 day−1 for the cultures grown the ambient CO2 without urea. Since there is no available carbon source and the bicarbonate buffer system collapses, the pH of cells cultured in the ambient CO2 without urea was increased rapidly up to 9.74 at day 5, thereby resulting in a decrease in cell growth or cell death after day 5. Consequently, under autotrophic conditions with ambient CO2, C. sorokiniana cells showed the best performance after the supply of 5 mM urea.
3.2.2. Effect of KH2PO4 Concentration on Biomass Production
C. sorokiniana cells were cultured with 0–10 mM KH2PO4 in the growth stage (Figure 2b). The culture supplemented with 5 mM KH2PO4 exhibited 7.28-fold higher biomass compared to the culture with ambient CO2 supply without KH2PO4. Under autotrophic conditions with ambient CO2, C. sorokiniana cells showed the best performance after supplementation with 5 mM KH2PO4. As a result, the cell density (1.33 g L−1) in the cells cultured with 5 mM KH2PO4 was 7.39-fold higher than that of the cells cultured at 0 mM KH2PO4 (0.18 g L−1) (Figure 2b). When KH2PO4 was injected at the optimum concentration, the increase in the biomass was lower than with 5 mM urea. Urea can directly supply both carbon and nitrogen, which are the most essential elements for cell growth. On the other hand, it is inferred that the increase in biomass is relatively low because KH2PO4 only contributes to the supply of phosphorus, while a stable pH only has a relatively auxiliary role in cell growth in culture. Therefore, to achieve optimal biomass productivity when the supply of CO2 is interrupted, we attempted to maximize the synergistic effect by supplying both urea and KH2PO4.
3.3. Synergistic Effect of the Combination of Urea and KH2PO4 on Microalgal Cell Growth
Biomass production was investigated when urea and KH2PO4 were injected simultaneously under low-concentration CO2 supply conditions. Urea and KH2PO4 were injected into the medium by combining various concentrations of urea and KH2PO4 at a concentration of 5 mM (the optimal concentration when injected alone into the culture medium) (Figure 3). As in the previous results, high cell growth rates were observed compared to conditions without urea and KH2PO4. In particular, regardless of the ratio of urea and KH2PO4, the difference in biomass production was only 0.18 g L−1 during the 4 days of cultivation. However, after 10 days of cell culture, when both urea and KH2PO4 were injected at a concentration of 5 mM, the biomass production reached 1.834 g L−1. This indicates an increase of 29.3% in biomass production compared to injection with 2.5 mM/2.5 mM (urea and KH2PO4 injected at a 1:1 ratio at low concentrations). In addition, it was confirmed that using a ratio of urea that is relatively high compared to the ratio of KH2PO4 results in increased cell growth. Specifically, compared to supplementation with urea 2.5 mM/KH2PO4 2.5 mM, when the KH2PO4 ratio was increased to 5.0 mM, the biomass increase rate was only 2.8%, but when the urea ratio increased to 5.0 mM, the biomass increase rate was 17.2% (6.1-fold higher than 5.0 mM of KH2PO4). As a result, when both urea and KH2PO4 concentrations were injected at 5 mM, biomass production (1.834 g L−1) was enhanced by 902.2%, 37.7%, and 14.1% compared to ambient CO2, KH2PO4 alone, and urea alone, respectively. Therefore, in 5% of CO2 supply, the biomass concentration is 1.96 g L−1 and decreases to 0.18 g L−1 when CO2 supply is halted, representing 91% loss of biomass. However, when urea and KH2PO4 is simultaneously provided, the biomass loss is reduced to only 9%.
3.4. Dissolved Inorganic Carbon (DIC) Concentration and pH of the Medium with Different Buffer Systems
3.4.1. DIC Concentration of the Medium
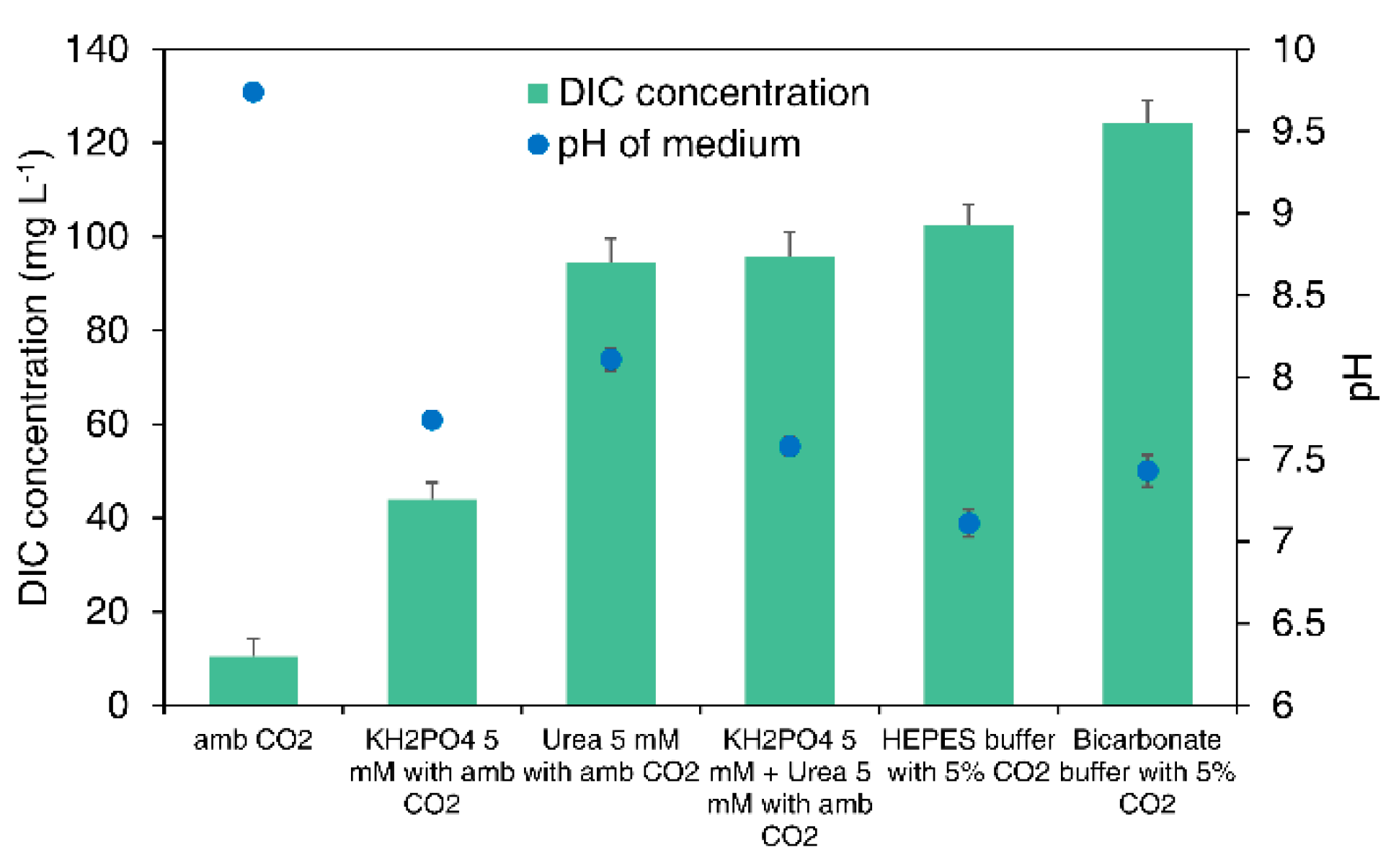
To analyze how urea and KH2PO4 contribute to an increased biomass production at low concentrations of CO2, the DIC concentration and pH in the culture medium were evaluated. Although gaseous CO2 can easily pass through the cell membrane, it is able to just as easily leak out of the cell. Carbonic anhydrase (CA) is involved in the carbon concentrating mechanism (CCM) under low CO2 supply conditions and converts gaseous CO2 into HCO3− ions, storing carbon in the cytoplasm, also known as the internal Ci pool. By consuming adenosine triphosphate (ATP), the cell can transport bicarbonate ions from the outside to the inside of the cell, providing a more stable carbon source for microalgal cells. At low concentrations of CO2, the DIC concentration in the culture medium is low. Therefore, cells obtain the carbon required for their survival via the CCM. However, this requires high energy and complex processes. By contrast, upon the injection of urea into the culture system, the content of inorganic carbon dissolved in the culture medium increases, and the overall DIC concentration rises to 94.4 mg L−1, 9.0 times higher than that without urea (Figure 4). Although this concentration is lower than that of the bicarbonate-based buffer system composed of DIC components, it is similar to the DIC concentration of the culture medium when a high concentration of CO2 (5% v/v) is supplied using a HEPES buffer at a laboratory scale. Therefore, the injection of urea and KH2PO4 can increase the concentration of DIC, allowing cells to utilize carbon sources easily. Increasing the levels of DIC is an efficient strategy to increase biomass and lipid production in autotrophic microalgal cultivation by improving sustained carbon availability and photosynthetic efficiency.
3.4.2. pH of the Medium
Using HEPES buffer and bicarbonate buffer systems with strong buffer capacity, a pH around 7 (7.11–7.43), the optimal pH for cell growth, was maintained despite the continuous supply of high concentrations of CO2. Bicarbonate buffer is produced by dissolving aerial CO2 in a highly alkaline solution, as shown in Equation (7):
CO2 + H2O → H2CO3 → HCO3− + H+ ↔ CO32− + 2H+
The equilibrium of the CO2 entities dissolved in water will highly depend on the pH and mixing system. Physically, some of CO2 can be stripped by CO2 bubbling. However, as the CO2 concentration in the flue gas (13–15%) is much more concentrated than that of atmospheric CO2 (0.04%), the amount of CO2. released from the system can be sufficiently compensated by newly dissolved CO2. However, if the supply of CO2 is interrupted, it is impossible to provide the DIC into the bicarbonate buffer system, causing an imbalance, eventually causing the buffer system to collapse. The disruption of the buffer system can cause severe changes in pH during microalgal culture, resulting in cell death. As shown in Figure 4, the pH increased to 9.74 under ambient CO2 conditions without an additional DIC supply. This pH range exceeds the normal microalgal cell culture conditions and can seriously reduce cell survival. We hypothesized that if urea was added to a culture with a low concentration of CO2, the cells would utilize some DIC as a carbon source for growth, while the remaining DIC would contribute to the bicarbonate buffer system to maintain an appropriate culture pH. However, maintaining a stable pH was difficult since, even with the supplementation of urea, the pH of the culture medium rose to pH 8.11 (Figure 4). Therefore, HPO42−, which is a component of the existing phosphate buffer system, was supplied in the form of KH2PO4 to further increase the buffer capacity. As a result, urea and KH2PO4 were simultaneously injected to minimize changes in the pH of the culture medium to pH 7.58, which was similar to the pH at a high concentration of CO2 in a bicarbonate buffer system. In other words, a stable pH was maintained by increasing the buffer capacity using KH2PO4 while also providing a carbon source and a nitrogen source through the injection of urea when the concentration of CO2 was insufficient.
3.5. Comparison of Microalgal Lipid Productivity According to Different Carbon Sources and Buffer Systems
The lipid performance of the microalgae was analyzed to determine whether the improved photosynthetic ability of microalgal cells under low-concentration CO2 supply conditions also enhanced lipid accumulation under stress conditions, such as nitrogen deficiency. Under ambient CO2 conditions, during which cell growth was inhibited due to low DIC supply and high pH conditions, 3.88% of lipids were accumulated inside the cells (Figure 5). Cells were found to have a low lipid content compared to normal culture conditions without nitrogen depletion. This is due to the fact that cell death began after 5 days of culture without an additional supply of urea and KH2PO4, as shown in Figure 2 and Figure 3, and metabolites that could be converted into lipids did not remain inside the cells. On the other hand, even under the same ambient CO2 supply condition, when KH2PO4 and urea were injected, metabolite accumulation occurred at the growth stage due to photosynthesis. Moreover, when the culture environment changed due to nitrogen deficiency, these metabolites could be converted into lipids. When KH2PO4 alone was injected (5 mM), the DIC concentration was 46.6% compared to when urea alone was injected (5 mM). However, the difference in lipid content was larger (61.1%) (Figure 5). This suggests that supplying KH2PO4 alone creates limited nutrient environment compared to supplying urea alone. As in the previous results, the highest lipid content (28.2%) was shown when urea and KH2PO4 were supplied simultaneously with low concentrations of CO2 (Figure 5). These results showed a high level of lipid production even when compared with the optimal culture condition (bicarbonate buffer with 5% CO2 supply condition) (33.3%). As a result, when urea and KH2PO4 were injected under ambient CO2 supply conditions, lipid productivity increased by around 7.28-fold compared to conditions without supplementation (Figure 5). This dramatic increase in the lipid productivity is a remarkable result considering the possibility of various applications of microalgae-derived biofuels. In particular, if biomass produced in the current culture system using a large-scale CO2 emission source, such as a power plant, is applied as a direct combustion fuel to a power plant, advantages including high energy yields, high CO2 fixation rates, and ease of fuel transportation can be achieved [4].
3.6. Outdoor Cultivation of Microalgae after Interruption of Flue Gas Supply from Coal-Fired Power Plant
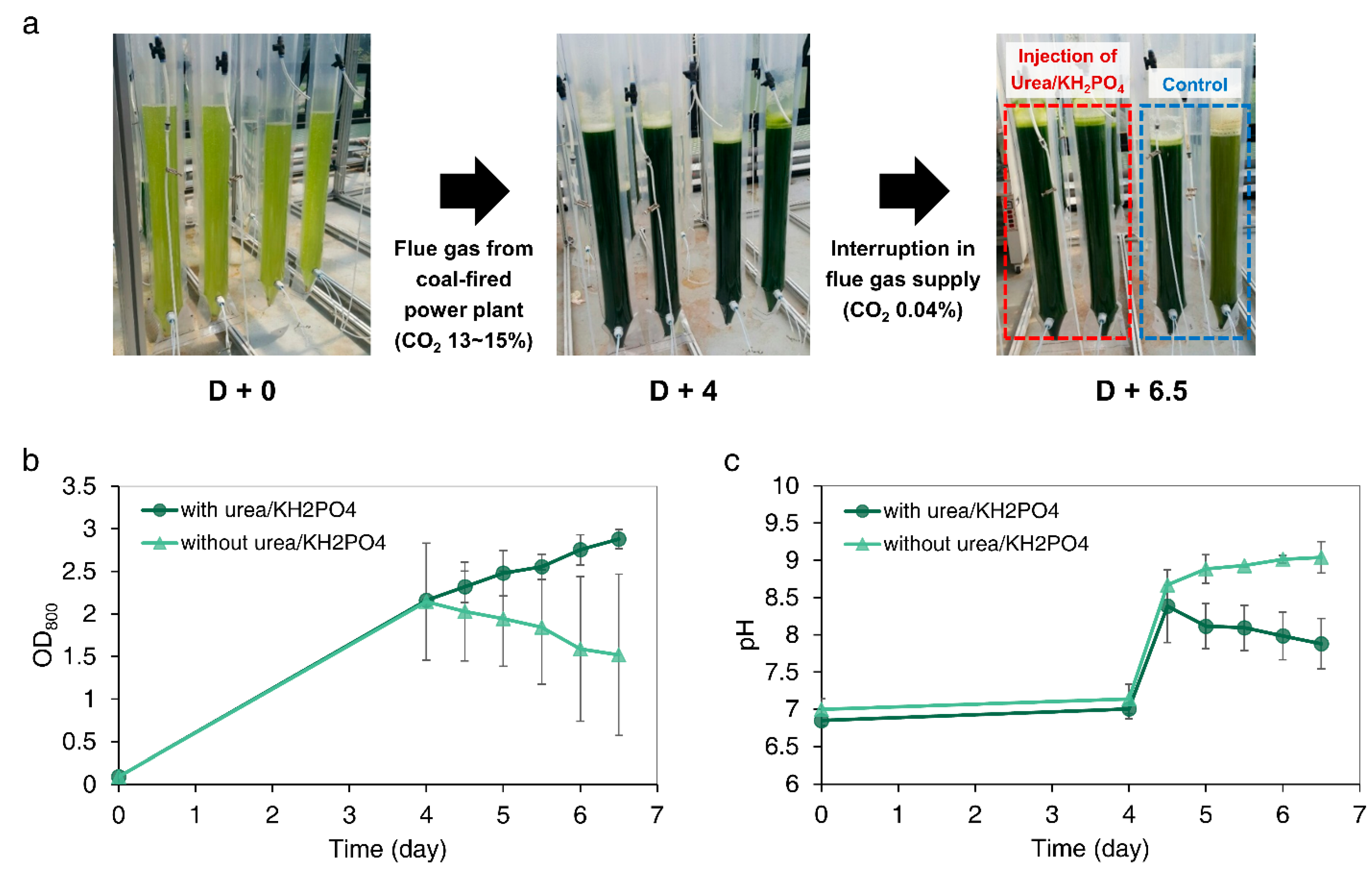
Biomass production was investigated when urea and KH2PO4 were injected simultaneously into a mass cultivation of microalgae near a coal-fired power plant when the flue gas supply was interrupted. A bicarbonate buffer system was formed by directly utilizing an alkaline solution and flue gas in 10 L PBR. C. sorokiniana cells were inoculated into the PBRs at an initial optical density (OD800) of 0.05. Microalgal cells were cultured using only sunlight and flue gas (Figure 6a). The flue gas emitted from coal-fired power plants was comprised of 13–15% of CO2 and a mixture of small amounts of NOx and SOx. However, due to the strong buffer capacity of the bicarbonate buffer system, the pH of the culture medium was maintained around pH 7 during the 4 days of cultivation, which resulted in continued cell growth, resulting in an OD800 of 2.15 (Figure 6a–c). Thereafter, to simulate the sudden interruption of flue gas containing a high concentration of CO2, the connection between the ring blower and the existing flue gas supply line was disconnected and connected to a bypass line that supplied ambient CO2. Although the cells did not sink due to constant aeration, this system provided a low concentration of CO2 (approximately 0.04%). At the same time, urea and KH2PO4 (both 5 mM) were simultaneously injected into the culture solution. As a result, the cells in the PBR injected with the chemical fertilizer showed sustained cell growth. After 2.5 days of additional culture, a 33% increase in the cell growth was observed. On the other hand, in the PBR where the chemical fertilizer was not supplied, a sudden drop in the cell growth of 29.1% was observed (Figure 6b). In particular, in the case of the fourth reactor (Figure 6a, right), the cells died rapidly (within 2.5 days), as the overall color was not bright green, but dark green.
Analyzing the pH of the culture medium, after the interruption of the flue gas supply, the pH of the cultures increased to 8.34~8.67 in 0.5 days. Thereafter, the pH of the PBR supplied with urea and KH2PO4 decreased gradually, reaching a final pH of 7.88, while the pH of the reactor without urea and KH2PO4 increased further, reaching a pH of 9.04. In conclusion, upon the interruption of the flue gas supply during microalgae cultivation, a simultaneous supplementation with urea and KH2PO4 resulted in a stable pH in the medium and an improved microbial cell growth.
Both urea and KH2PO4 were injected at a concentration of 5 mM in the case of predictable CO2 interruption situation. However, if the interruption of CO2 flow can be unpredictable, an alternative solution should be required. For example, a CO2 sensor that monitors the CO2 concentration of the flue gas supply can be connected to an automated system capable of injecting urea and KH2PO4 to the PBR. When the CO2 concentration becomes much lower than the normal flue gas composition, CO2 interruption can be easily recognized and rapidly feedbacked by the addition of urea and KH2PO. In this way, it would be possible to maintain the normal growth of microalgal culture even during unexpected CO2 supply interruption.
4. Conclusions
Sustaining a neutral and mildly alkaline pH is one of the most important factors during microalgal cultivation. To reduce a large amount of CO2 and produce value-added products, a scaled-up microalgae cultivation system was installed near CO2-emiting companies, such as a coal-fired power plant and Korea District Heating Corporation. In a previous study, we developed a buffer system for maintaining pH using KOH and phosphate. The mixture of bicarbonate and phosphate buffer systems was cost-efficient and supplies nutrients for cell growth. However, flue gas from CO2-emiting companies is not a reliable source of CO2 since CO2 blowers, which drive CO2 from the CO2 source into the cultured cells, periodically require repairs or overhauls. Therefore, in the present study, we proposed a continuous culture strategy based on urea and KH2PO4 in an outdoor autotrophic culture. For the cultivation of 1 ton scale of C. sorokiniana, the cost of artificially provided CO2 is $531.68 (from DONG-A Industrial Gas, Seoul, Republic of Korea). Supplementation of 5 mM urea and 5 mM KH2PO4 for 1 ton cultivation costs $0.58 ($1.93 kg−1; 57-13-6 from DUKSAN science) and $ 30.41 ($44.69 kg−1; 7778-77-0 from DUKSAN science), respectively, thereby reducing the production cost by 99.89% and $94.28, respectively. In total, 94.17% of cost can be reduced by using 5 mM of urea and KH2PO4 simultaneously. Using C. sorokiniana cells, the resulting culture system showed markedly enhanced biomass and lipid productivity upon CO2 supply interruption.
Author Contributions
Conceptualization, B.S.Y., Y.J.S., and S.J.S.; methodology, B.S.Y., Y.J.S. and M.E.H.; writing—original draft preparation, B.S.Y. and Y.J.S.; writing—review and editing, B.S.Y., Y.J.S., and M.E.H.; supervision, S.J.S.; project administration, S.J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the “Carbon to X Project” (no. 2020M3H7A1098295), which was funded by the National Research Foundation (NRF) funded by the Ministry of Science and ICT, Republic of Korea, grant (no. NRF-2019R1A2C3009821/2020R1A5A1018052) from the National Research Foundation of Korea (NRF), a grant from Korea Institute of Energy Technology Evaluation and Planning (KETEP) (no. 20172010202050), and the Korea University Grant.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Korea Western Power Co., Ltd. for providing the cultivation sites and facilities for this study along with their funding support. The authors also would like to thank Hong Ki Yoon for supporting the cultivation of microalgae at the coal-fired power plant (Taean, Republic of Korea).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Solomon, S.; Plattner, G.K.; Knutti, R.; Friedlingstein, P. Irreversible Climate Change Due to Carbon Dioxide Emissions. Proc. Natl. Acad. Sci. USA 2009, 106, 1704–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekwurzel, B.; Boneham, J.; Dalton, M.W.; Heede, R.; Mera, R.J.; Allen, M.R.; Frumhoff, P.C. The Rise in Global Atmospheric CO2, Surface Temperature, and Sea Level from Emissions Traced to Major Carbon Producers. Clim. Chang. 2017, 144, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, T.A.; Kler, J.S.; Hernke, M.T.; Braun, R.K.; Meyer, K.C.; Funk, W.E. Direct Human Health Risks of Increased Atmospheric Carbon Dioxide. Natl. Sustain. 2019, 2, 691–701. [Google Scholar] [CrossRef]
- Choi, H.I.; Lee, J.S.; Choi, J.W.; Shin, Y.S.; Sung, Y.J.; Hong, M.E.; Kwak, H.S.; Kim, C.Y.; Sim, S.J. Performance and Potential Appraisal of Various Microalgae as Direct Combustion Fuel. Bioresour. Technol. 2019, 273, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Liu, Q.; Qi, Y.; Chen, G.; Song, Y.; Kansha, Y.; Kitamura, Y. Absorption-Microalgae Hybrid CO2 Capture and Biotransformation Strategy—A Review. Int. J. Greenh. Gas Control. 2019, 88, 109–117. [Google Scholar] [CrossRef]
- Hong, M.E.; Yu, B.S.; Patel, A.K.; Choi, H.I.; Song, S.; Sung, Y.J.; Chang, W.S.; Sim, S.J. Enhanced Biomass and Lipid Production of Neochloris oleoabundans Under High Light Conditions by Anisotropic Nature of Light-Splitting CaCO3 Crystal. Bioresour. Technol. 2019, 287, 121483. [Google Scholar] [CrossRef] [PubMed]
- Melis, A. Photosynthesis-to-Fuels: From Sunlight to Hydrogen, Isoprene, and Botryococcene Production. Energy Environ. Sci. 2012, 5, 5531–5539. [Google Scholar] [CrossRef]
- Kang, C.D.; Lee, J.S.; Park, T.H.; Sim, S.J. Comparison of Heterotrophic and Photoautotrophic Induction on Astaxanthin Production by Haematococcus pluvialis. Appl. Microbiol. Biotechnol. 2005, 68, 237–241. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Joun, J.M.; Lee, J.; Hong, M.E.; Pham, H.M.; Chang, W.S.; Sim, S.J. Development of Large-Scale and Economic pH Control System for Outdoor Cultivation of Microalgae Haematococcus pluvialis Using Industrial Flue Gas. Bioresour. Technol. 2017, 244, 1235–1244. [Google Scholar] [CrossRef]
- Choi, S.Y.; Wang, J.Y.; Kwak, H.S.; Lee, S.M.; Um, Y.; Kim, Y.; Sim, S.J.; Choi, J.I.; Woo, H.M. Improvement of Squalene Production from CO2 in Synechococcus elongatus PCC 7942 by Metabolic Engineering and Scalable Production in a Photobioreactor. ACS Synth. Biol. 2017, 6, 1289–1295. [Google Scholar] [CrossRef]
- Chen, C.Y.; Kao, P.C.; Tsai, C.J.; Lee, D.J.; Chang, J.S. Engineering Strategies for Simultaneous Enhancement of C-Phycocyanin Production and CO2 Fixation with Spirulina platensis. Bioresour. Technol. 2013, 145, 307–312. [Google Scholar] [CrossRef]
- Hong, M.E.; Hwang, S.K.; Chang, W.S.; Kim, B.W.; Lee, J.; Sim, S.J. Enhanced Autotrophic Astaxanthin Production from Haematococcus pluvialis Under High Temperature via Heat Stress-Driven Haber–Weiss Reaction. Appl. Microbiol. Biotechnol. 2015, 99, 5203–5215. [Google Scholar] [CrossRef]
- Huntley, M.E.; Redalje, D.G. CO2 Mitigation and Renewable Oil from Photosynthetic Microbes: A New Appraisal. Mitig. Adapt. Strat. Glob. Chang. 2007, 12, 573–608. [Google Scholar] [CrossRef]
- Demirbaş, A. Oily Products from Mosses and Algae via Pyrolysis. Energy Sources A 2006, 28, 933–940. [Google Scholar] [CrossRef]
- Tomei, J.; Helliwell, R. Food Versus Fuel? Going Beyond Biofuels. Land Use Policy 2016, 56, 320–326. [Google Scholar] [CrossRef]
- Gouveia, L.; Oliveira, A.C. Microalgae as a Raw Material for Biofuels Production. J. Ind. Microbiol. 2009, 36, 269–274. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Moheimani, N.R. Sustainable Biofuels from Algae. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 13–25. [Google Scholar] [CrossRef]
- Suganya, T.; Varman, M.; Masjuki, H.H.; Renganathan, S. Macroalgae and Microalgae as a Potential Source for Commercial Applications Along with Biofuels Production: A Biorefinery Approach. Renew. Sustain. Energy Rev. 2016, 55, 909–941. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Hong, M.E.; Chang, W.S.; Sim, S.J. Autotrophic Biodiesel Production from the Thermotolerant Microalga Chlorella sorokiniana by Enhancing the Carbon Availability with Temperature Adjustment. Biotechnol. Bioproc. E 2019, 24, 223–231. [Google Scholar] [CrossRef]
- Mondal, M.; Goswami, S.; Ghosh, A.; Oinam, G.; Tiwari, O.N.; Das, P.; Gayen, K.; Mandal, M.K.; Halder, G.N. Production of Biodiesel from Microalgae Through Biological Carbon Capture: A Review. 3 Biotech 2017, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.Y.; Hong, M.E.; Chang, W.S.; Sim, S.J. Enhanced Biodiesel Production in Neochloris oleoabundans by a Semi-Continuous Process in Two Stage Photobioreactors. Bioprocess Biosyst. Eng. 2015, 38, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Cinar, S.O.; Chong, Z.K.; Kucuker, M.A.; Wieczorek, N.; Cengiz, U.; Kuchta, K. Bioplastic Production from Microalgae: A Review. Int. J. Environ. Res. Public Health 2020, 17, 3842. [Google Scholar] [CrossRef] [PubMed]
- Hempel, F.; Bozarth, A.S.; Lindenkamp, N.; Klingl, A.; Zauner, S.; Linne, U.; Steinbüchel, A.; Maier, U.G. Microalgae as Bioreactors for Bioplastic Production. Microb. Cell Factories 2011, 10, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant Potential of Microalgae in Relation to Their Phenolic and Carotenoid Content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Kang, C.D.; Sim, S.J. Direct Extraction of Astaxanthin from Haematococcus Culture Using Vegetable Oils. Biotechnol. Lett. 2008, 30, 441–444. [Google Scholar] [CrossRef]
- Hong, M.E.; Choi, Y.Y.; Sim, S.J. Effect of Red Cyst Cell Inoculation and Iron(II) Supplementation on Autotrophic Astaxanthin Production by Haematococcus pluvialis Under Outdoor Summer Conditions. J. Biotechnol. 2016, 218, 25–33. [Google Scholar] [CrossRef]
- Mulders, K.J.M.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic Pigment Production with Microalgae: Biological Constraints and Opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef]
- Christaki, E.; Bonos, E.; Florou-Paneri, P. Innovative Microalgae Pigments as Functional Ingredients in Nutrition. Biotechnol. Adv. 2015, 233–243. [Google Scholar]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Bošnjaković, M.; Sinaga, N. The Perspective of Large-Scale Production of Algae Biodiesel. Appl. Sci. 2020, 10, 8181. [Google Scholar] [CrossRef]
- Bekirogullari, M.; Fragkopoulos, I.S.; Pittman, J.K.; Theodoropoulos, C. Production of Lipid-Based Fuels and Chemicals from Microalgae: An Integrated Experimental and Model-Based Optimization Study. Algal Res. 2017, 23, 78–87. [Google Scholar] [CrossRef]
- Griffiths, M.J.; Harrison, S.T.L. Lipid Productivity as a Key Characteristic for Choosing Algal Species for Biodiesel Production. J. Appl. Phycol. 2009, 21, 493–507. [Google Scholar] [CrossRef]
- Bleeke, F.; Milas, M.; Winckelmann, D.; Klöck, G. Optimization of Freshwater Microalgal Biomass Harvest Using Polymeric Flocculants. Int. Aquat. Res. 2015, 7, 235–244. [Google Scholar] [CrossRef] [Green Version]
- Slade, R.; Bauen, A. Micro-Algae Cultivation for Biofuels: Cost, Energy Balance, Environmental Impacts and Future Prospects. Biomass Bioenergy 2013, 53, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Sung, Y.J.; Patel, A.K.; Yu, B.S.; Choi, H.I.; Kim, J.R.; Jin, E.S.; Sim, S.J. Sedimentation Rate-Based Screening of Oleaginous Microalgae for Utilization as a Direct Combustion Fuel. Bioresour. Technol. 2019, 293, 122045. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, W.; Mao, X.; Li, Y.; Wu, T.; Chen, F. High-Value Biomass from Microalgae Production Platforms: Strategies and Progress Based on Carbon Metabolism and Energy Conversion. Biotechnol. Biofuels 2018, 11, 227. [Google Scholar] [CrossRef] [Green Version]
- Ramanna, L.; Rawat, I.; Bux, F. Light Enhancement Strategies Improve Microalgal Biomass Productivity. Renew. Sustain. Energy Rev. 2017, 80, 765–773. [Google Scholar] [CrossRef]
- Hong, M.E.; Choi, H.I.; Kwak, H.S.; Hwang, S.W.; Sung, Y.J.; Chang, W.S.; Sim, S.J. Rapid Selection of Astaxanthin-Hyperproducing Haematococcus Mutant via Azide-Based Colorimetric Assay Combined with Oil-Based Astaxanthin Extraction. Bioresour. Technol. 2018, 267, 175–181. [Google Scholar] [CrossRef]
- Pham, H.M.; Kwak, H.S.; Hong, M.E.; Lee, J.W.; Chang, W.S.; Sim, S.J. Development of an X-Shape Airlift Photobioreactor for Increasing Algal Biomass and Biodiesel Production. Bioresour. Technol. 2017, 239, 211–218. [Google Scholar] [CrossRef]
- Praveenkumar, R.; Lee, K.B.; Lee, J.Y.; Oh, Y.K. Breaking Dormancy: An Energy-Efficient Means of Recovering Astaxanthin from Microalgae. Green Chem. 2015, 17, 1226–1234. [Google Scholar] [CrossRef]
- Gabrielli, P.; Gazzani, M.; Mazzotti, M. The Role of Carbon Capture and Utilization, Carbon Capture and Storage, and Biomass to Enable a Net-Zero-CO2 Emissions Chemical Industry. Ind. Eng. Chem. Res. 2020, 59, 7033–7045. [Google Scholar] [CrossRef] [Green Version]
- Takht Ravanchi, M.T.; Sahebdelfar, S. Carbon Dioxide Capture and Utilization in Petrochemical Industry: Potentials and Challenges. Appl. Petrochem. Res. 2014, 4, 63–77. [Google Scholar] [CrossRef] [Green Version]
- House, K.Z.; Harvey, C.F.; Aziz, M.J.; Schrag, D.P. The Energy Penalty of Post-Combustion CO2 Capture & Storage and Its Implications for Retrofitting the U.S. Installed Base. Energy Environ. Sci. 2009, 2, 193–205. [Google Scholar]
- Sung, Y.J.; Lee, J.S.; Yoon, H.K.; Ko, H.J.; Sim, S.J. Outdoor Cultivation of Microalgae in a Coal-Fired Power Plant for Conversion of Flue Gas CO2 into Microalgal Direct Combustion Fuels. SMAB 2021, 1, 1–10. [Google Scholar]
- Yoo, J.J.; Choi, S.P.; Kim, J.Y.H.; Chang, W.S.; Sim, S.J. Development of Thin-Film Photo-Bioreactor and Its Application to Outdoor Culture of Microalgae. Bioproc. Biosyst. Eng. 2013, 36, 729–736. [Google Scholar] [CrossRef]
- Sayre, R. Microalgae: The Potential for Carbon Capture. J. Biosci. 2010, 60, 722–727. [Google Scholar] [CrossRef]
- Yadav, G.; Dubey, B.K.; Sen, R. A Comparative Life Cycle Assessment of Microalgae Production by CO2 Sequestration from Flue Gas in Outdoor Raceway Ponds Under Batch and Semi-Continuous Regime. J. Clean. Prod. 2020, 258. [Google Scholar] [CrossRef]
- Hong, M.E.; Chang, W.S.; Patel, A.K.; Oh, M.S.; Lee, J.J.; Sim, S.J.; Carbon, M.-B. Microalgal-Based Carbon Sequestration by Converting LNG-Fired Waste CO2 into Red Gold Astaxanthin: The Potential Applicability. Energies 2019, 12, 1718. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Jiang, J.; Fa, Y. Overcoming the Biological Contamination in Microalgae and Cyanobacteria Mass Cultivations for Photosynthetic Biofuel Production. Molecules 2020, 25, 5220. [Google Scholar] [CrossRef]
- Ma, M.; Yuan, D.; He, Y.; Park, M.S.; Gong, Y.; Hu, Q. Effective Control of Poterioochromonas malhamensis in Pilot-Scale Culture of Chlorella sorokiniana GT-1 by Maintaining CO2-Mediated Low Culture pH. GT. Algal Res. 2017, 26, 436–444. [Google Scholar] [CrossRef]
- Aishvarya, V.; Pradhan, N.; Nayak, R.R.; Sukla, L.B.; Mishra, B.K. Enhanced Inorganic Carbon Uptake by Chlorella sp. IMMTCC-2 under Autotrophic Conditions for Lipid Production and CO2 Sequestration. J. Appl. Phycol. 2012, 24, 1455–1463. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, S.J.; Hong, M.E.; Chang, W.S.; Sim, S.J. Repeated-Batch Production of omega-3 Enriched Biomass of Chlorella sorokiniana via Calcium-Induced Homeoviscous Adaptation. Bioresour. Technol. 2020, 303, 122944. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.Y.; Hong, M.E.; Jin, E.S.; Woo, H.M.; Sim, S.J. Improvement in Modular Scalability of Polymeric Thin-Film Photobioreactor for Autotrophic Culturing of Haematococcus pluvialis Using Industrial Flue Gas. Bioresour. Technol. 2018, 249, 519–526. [Google Scholar] [CrossRef]
- Dhup, S.; Kannan, D.C.; Dhawan, V. Understanding Urea Assimilation and Its Effect on Lipid Production and Fatty Acid Composition of Scenedesmus sp. SOJ Biochem. 2016, 2, 7. [Google Scholar] [CrossRef] [Green Version]
- Giordano, C.; Erpen, C.; Yao, W.; Antonietti, M. Synthesis of Mo and W Carbide and Nitride Nanoparticles via a Simple “Urea Glass” Route. Nano Lett. 2008, 8, 4659–4663. [Google Scholar] [CrossRef]
- Radkova, M.; Stoyneva-Gärtner, M.S.; Dincheva, I.; Stoykova, P.; Uzunov, B.; Dimitrova, P.; Borisova, C.; Gärtner, G. Chlorella vulgaris H1993 and Desmodesmus communis H522 for Low-Cost Production of High-Value Microalgal Products. Biotechnol. Biotechnol. Equip. 2019, 33, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Goswami, R.C.D.; Kalita, M.C. Scenedesmus dimorphus and Scenedesmus quadricauda: Two Potent Indigenous Microalgae Strains for Biomass Production and CO2 Mitigation—A Study on Their Growth Behavior and Lipid Productivity under Different Concentration of Urea as Nitrogen Source. J. Algal Biomass Utln 2011, 2, 42–49. [Google Scholar]
Figure 1.
Injection of chemical fertilizers, such as cost-effective urea ($1.93 kg−1; 57-13-6 from DUKSAN science) and KH2PO4 ($44.69 kg−1; 7778-77-0 from DUKSAN science), to improve microalgal biomass production upon interruption of CO2 supply.
Figure 1.
Injection of chemical fertilizers, such as cost-effective urea ($1.93 kg−1; 57-13-6 from DUKSAN science) and KH2PO4 ($44.69 kg−1; 7778-77-0 from DUKSAN science), to improve microalgal biomass production upon interruption of CO2 supply.

Figure 2.
Biomass production of Chlorella sorokiniana UTEX 2714 using chemical fertilizers. Biomass production according to concentrations (0, 1, 3, 5, 8, and 10 mM) of (a) urea and (b) KH2PO4 injections with ambient CO2 supply. Results represent the mean ± standard deviation (SD) of three replicates.
Figure 2.
Biomass production of Chlorella sorokiniana UTEX 2714 using chemical fertilizers. Biomass production according to concentrations (0, 1, 3, 5, 8, and 10 mM) of (a) urea and (b) KH2PO4 injections with ambient CO2 supply. Results represent the mean ± standard deviation (SD) of three replicates.

Figure 3.
Biomass concentration of Chlorella sorokiniana UTEX 2714 with different combinations of urea and KH2PO4 concentrations. Results represent the mean ± standard deviation (SD) of three replicates.
Figure 3.
Biomass concentration of Chlorella sorokiniana UTEX 2714 with different combinations of urea and KH2PO4 concentrations. Results represent the mean ± standard deviation (SD) of three replicates.

Figure 4.
Comparison of dissolved inorganic carbon (DIC) concentration and pH of the medium with different CO2 concentrations (ambient CO2 and 5% CO2), chemical fertilizers (urea and KH2PO4), and buffer systems (HEPES buffer and bicarbonate buffer) after 4 days of microalgae cultivation. Results represent the mean ± standard deviation (SD) of three replicates.
Figure 4.
Comparison of dissolved inorganic carbon (DIC) concentration and pH of the medium with different CO2 concentrations (ambient CO2 and 5% CO2), chemical fertilizers (urea and KH2PO4), and buffer systems (HEPES buffer and bicarbonate buffer) after 4 days of microalgae cultivation. Results represent the mean ± standard deviation (SD) of three replicates.

Figure 5.
Lipid content and productivity of Chlorella sorokiniana UTEX 2714 after 6 days of nitrogen starvation with different concentrations of CO2 supplementation (ambient CO2 and 5% CO2) using urea, KH2PO4, and bicarbonate buffer system. Results represent the mean ± standard deviation (SD) of three replicates.
Figure 5.
Lipid content and productivity of Chlorella sorokiniana UTEX 2714 after 6 days of nitrogen starvation with different concentrations of CO2 supplementation (ambient CO2 and 5% CO2) using urea, KH2PO4, and bicarbonate buffer system. Results represent the mean ± standard deviation (SD) of three replicates.

Figure 6.
Outdoor cultivation of Chlorella sorokiniana UTEX 2714 using flue gas from coal-fired power plant. (a) Images of 10-L bubble column polymeric photobioreactor during cell culture. Changes of (b) optical density (OD) and (c) pH of the medium for 6.5 days of cell cultivation. Algal cells were cultivated using flue gas (CO2 13~15%) for 4 days and with supplementation of urea and KH2PO4 under interruption in flue gas supply for 2.5 days. Results represent the mean ± standard deviation (SD) of three replicates.
Figure 6.
Outdoor cultivation of Chlorella sorokiniana UTEX 2714 using flue gas from coal-fired power plant. (a) Images of 10-L bubble column polymeric photobioreactor during cell culture. Changes of (b) optical density (OD) and (c) pH of the medium for 6.5 days of cell cultivation. Algal cells were cultivated using flue gas (CO2 13~15%) for 4 days and with supplementation of urea and KH2PO4 under interruption in flue gas supply for 2.5 days. Results represent the mean ± standard deviation (SD) of three replicates.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yu, B.S.; Sung, Y.J.; Hong, M.E.; Sim, S.J. Improvement of Photoautotrophic Algal Biomass Production after Interrupted CO2 Supply by Urea and KH2PO4 Injection. Energies 2021, 14, 778. https://0-doi-org.brum.beds.ac.uk/10.3390/en14030778
AMA Style
Yu BS, Sung YJ, Hong ME, Sim SJ. Improvement of Photoautotrophic Algal Biomass Production after Interrupted CO2 Supply by Urea and KH2PO4 Injection. Energies. 2021; 14(3):778. https://0-doi-org.brum.beds.ac.uk/10.3390/en14030778
Chicago/Turabian StyleYu, Byung Sun, Young Joon Sung, Min Eui Hong, and Sang Jun Sim. 2021. "Improvement of Photoautotrophic Algal Biomass Production after Interrupted CO2 Supply by Urea and KH2PO4 Injection" Energies 14, no. 3: 778. https://0-doi-org.brum.beds.ac.uk/10.3390/en14030778
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.