
Assessment of Microbial Diversity during Thermophilic Anaerobic Co-Digestion for an Effective Valorization of Food Waste and Wheat Straw

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates and Inoculum Sources
2.2. Bioreactors and Operating Conditions
2.3. Analytical Methods
2.4. Microbial Community Analysis
3. Results
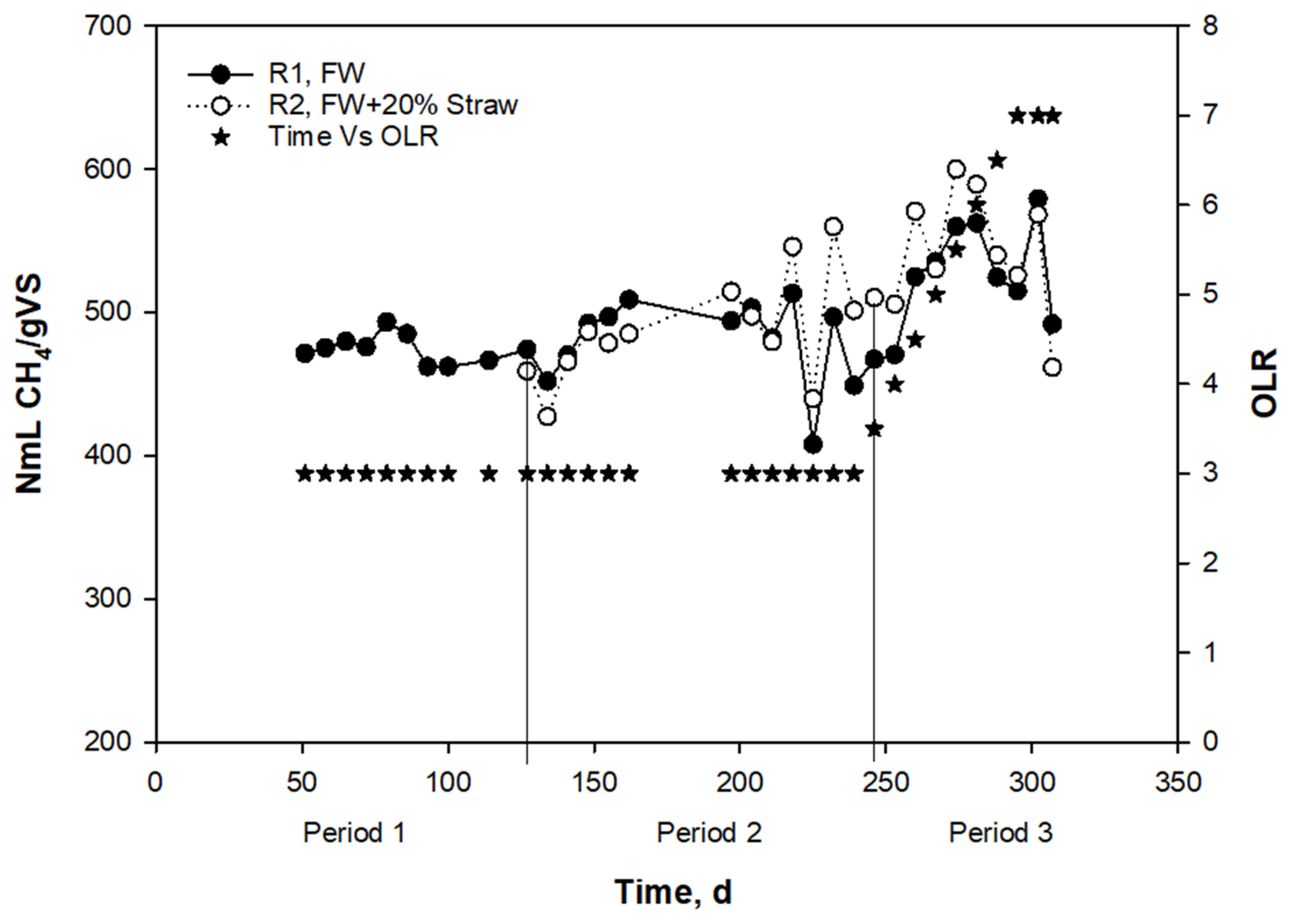
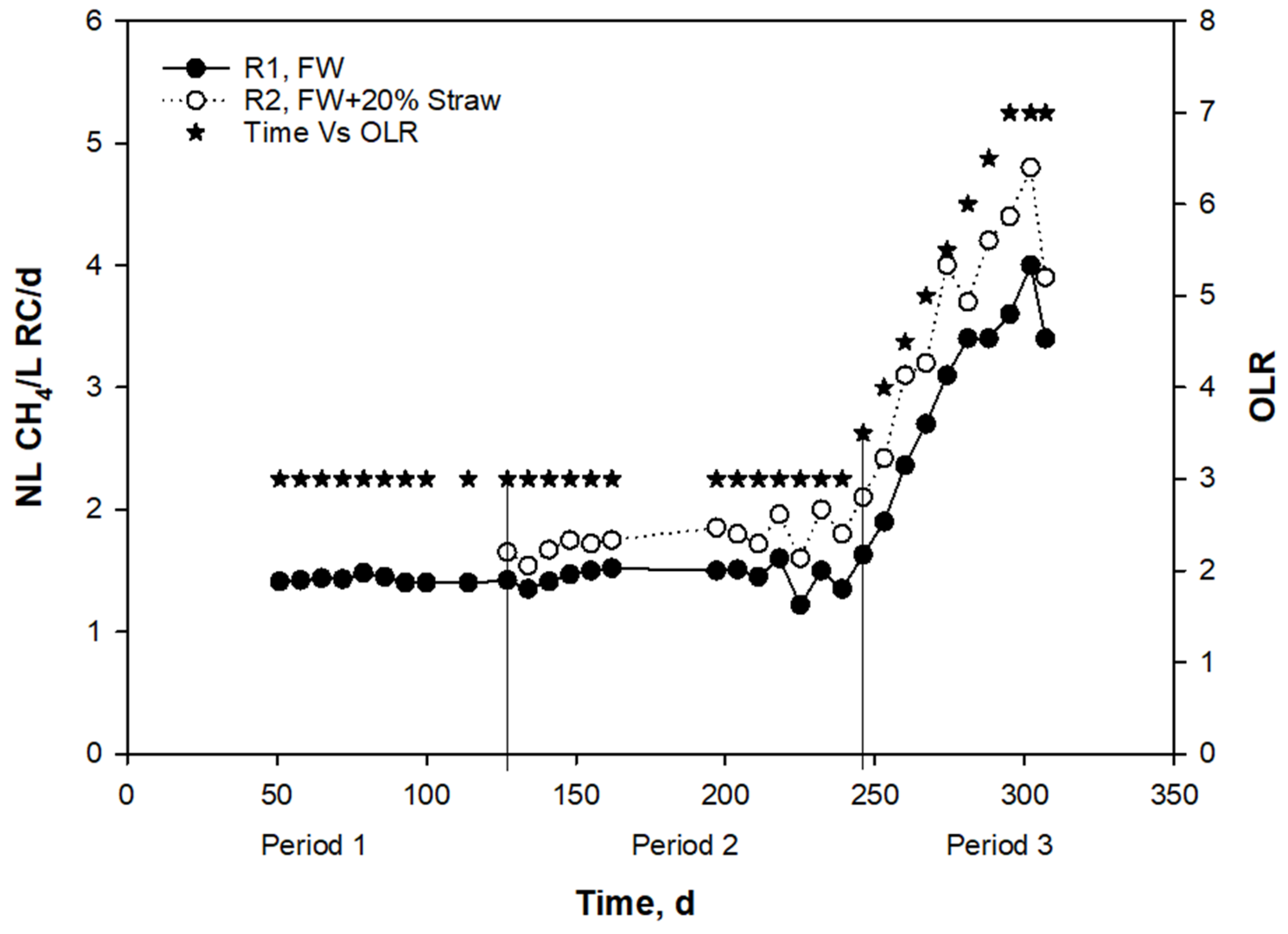
3.1. Effects of Straw Addition during Anaerobic Digestion of Food Waste at Low and High OLRs
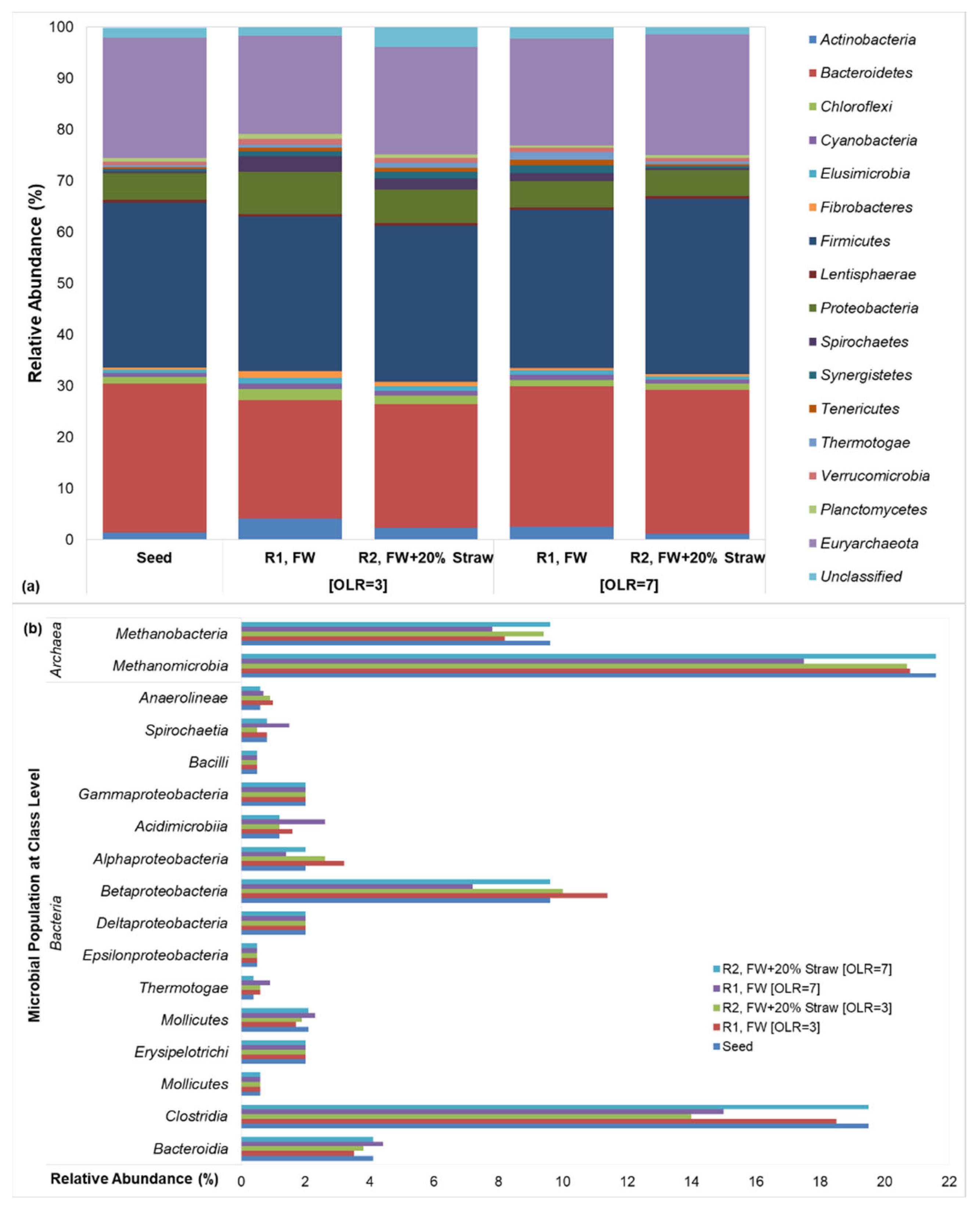
3.2. Assessment of Microbial Diversity
3.2.1. Hydrolytic and Acidogenic Bacteria
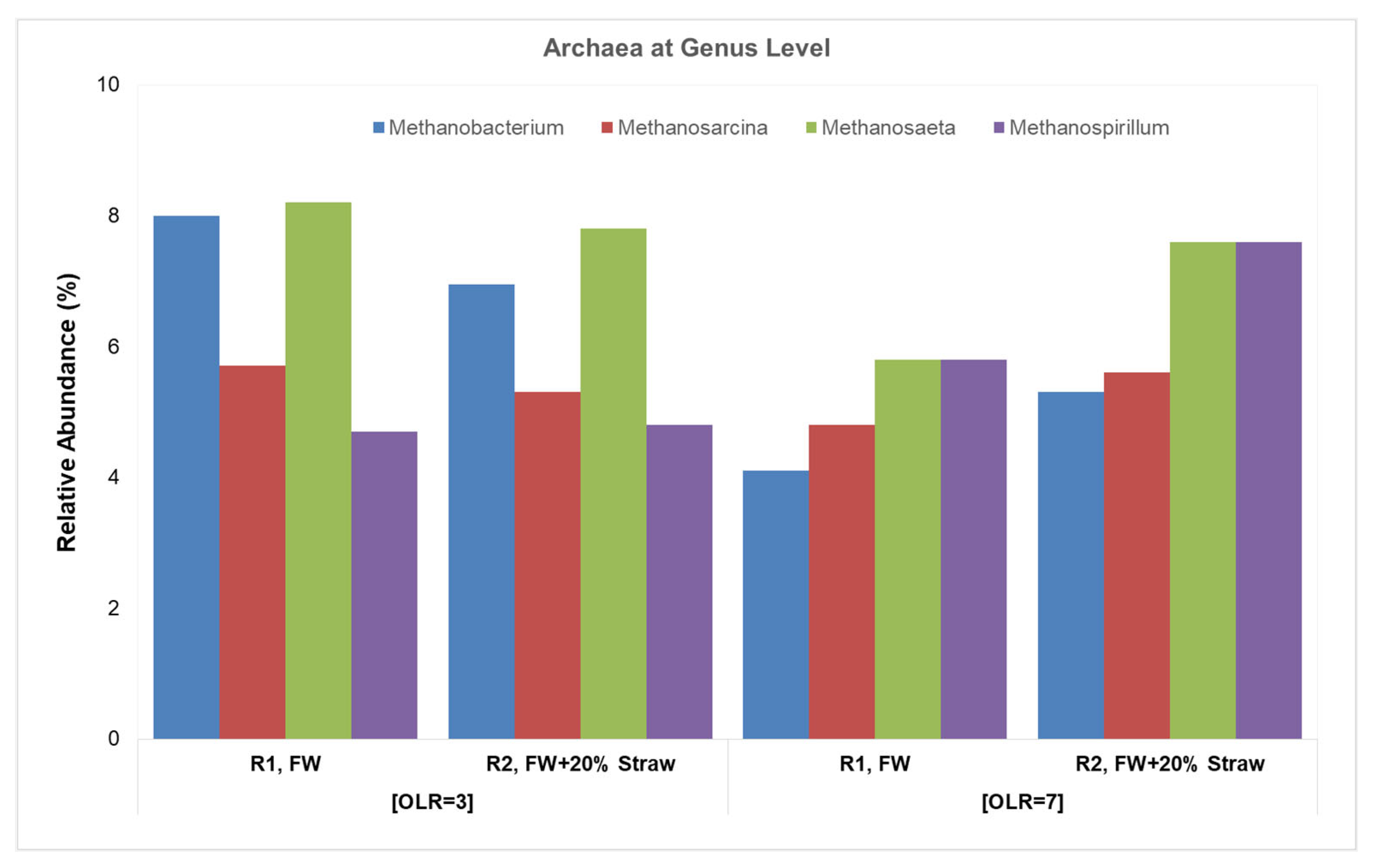
3.2.2. Acetoclastic and Hydrogenotrophic Methanogens
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gomec, C.Y.; Gonuldinc, S.; Eldem, N.; Ozturk, I. Behaviour if an up-flow anaerobic sludge bed reactor (UASB) reactor at extreme salinity. Water Sci. Technol. 2005, 51, 115–120. [Google Scholar] [CrossRef]
- Wang, X.; Yang, G.; Feng, Y.; Ren, G.; Han, X.O. Optimizing feeding composition and carbon–nitrogen ratios for improved methane yield during anaerobic codigestion of dairy, chicken manure and wheat straw. Bioresour. Technol. 2012, 120, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Zeshan Karthikeyan, O.P.; Visvanathan, C. Effect of C/N ratio and ammonia-N accumulation in a pilot-scale thermophilic dry anaerobic digester. Bioresour. Technol. 2012, 113, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Yangin-Gomec, C.; Ozturk, I. Effect of maize silage addition on biomethane recovery from mesophilic co-digestion of chicken and cattle manure to suppress ammonia inhibition. Energy. Convers. Manag. 2013, 71, 92–100. [Google Scholar] [CrossRef]
- Zhang, C.; Su, H.; Baeyens, J.; Tan, T. Reviewing the anaerobic digestion of food waste for biogas production. Renew. Sust. Energy Rev. 2014, 38, 383–392. [Google Scholar] [CrossRef]
- Yangin-Gomec, C.; Pekyavas, G.; Sapmaz, T.; Aydin, S.; Ince, B.; Akyol, Ç.; Ince, O. Microbial monitoring of ammonia removal in a UASB reactor treating pre-digested chicken manure with anaerobic granular inoculum. Bioresour. Technol. 2017, 241, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Pekyavas, G.; Yangin-Gomec, C. Response of Anammox bacteria to elevated nitrogen and organic matter in pre-digested chicken waste at a long-term operated UASB reactor initially seeded by methanogenic granules. Bioresour. Technol. Rep. 2019, 7, 100222. [Google Scholar] [CrossRef]
- Mirmohamadsadeghi, S.; Karimi, K.; Tabatabaei, M.; Aghbashlo, M. Biogas production from food wastes: A review on recent developments and future perspectives. Bioresour. Technol. Rep. 2019, 7, 100202. [Google Scholar] [CrossRef]
- Moestedt, J.; Müller, B.; Westerholm, M.; Schnürer, A. Ammonia threshold for inhibition of anaerobic digestion of thin stillage and the importance of organic loading rate. Microb Biotechnol. 2016, 9, 180–194. [Google Scholar] [CrossRef]
- Dhar, H.; Kumar, P.; Kumar, S.; Mukherjee, S.; Vaidya, A.N. Effect of organic loading rate during anaerobic digestion of municipal solid waste. Bioresour. Technol. 2016, 217, 56–61. [Google Scholar] [CrossRef]
- Westerholm, M.; Schnürer, A. Microbial responses to different operating practices for biogas production systems. In Anaerobic Digestion; Banu, J.R., Ed.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Pullammanappallil, P.C.; Chynoweth, D.P.; Lyberatos, G.; Svoronos, S.A. Stable performance of anaerobic digestion in the presence of a high concentration of propionic acid. Bioresour. Technol. 2001, 78, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, W.; Anwar, N.; Ma, Z.; Liu, G.; Zhang, R. Effect of organic loading rate on anaerobic digestion of food waste under mesophilic and thermophilic conditions. Energy Fuel 2017, 31, 2976–2984. [Google Scholar] [CrossRef]
- Guo, X.; Wang, C.; Sun, F.; Zhu, W.; Wu, W. A comparison of microbial characteristics between the thermophilic and mesophilic anaerobic digesters exposed to elevated food waste loadings. Bioresour. Technol. 2014, 152, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xiao, G.; Peng, L.; Su, H.; Tan, T. The anaerobic co-digestion of food waste and cattle manure. Bioresour. Technol. 2013, 129, 170–176. [Google Scholar] [CrossRef]
- Dereli, R.K.; Yangin-Gomec, C.; Ozabali, A.; Ozturk, I. The feasibility of a centralized biogas plant treating the manure produced by an organized animal farmers union in Turkey. Water Sci. Technol. 2012, 66, 556–563. [Google Scholar] [CrossRef]
- Neves, L.; Oliveira, R.; Alves, M.M. Co-digestion of cow manure, food waste and intermittent input of fat. Bioresour. Technol. 2009, 100, 1957–1962. [Google Scholar] [CrossRef] [Green Version]
- Haider, M.R.; Yousaf Zeshan Malik, R.N.; Visvanathan, C. Effect of mixing ratio of food waste and rice husk co-digestion and substrate to inoculum ratio on biogas production. Bioresour. Technol. 2015, 190, 451–457. [Google Scholar] [CrossRef]
- Demirbas, A. Biogas potential of manure and straw mixtures. Energy Sources 2006, 28, 71–78. [Google Scholar] [CrossRef]
- Tian, S.-Q.; Zhao, R.-Y.; Chen, Z.-C. Review of the pretreatment and bioconversion of lignocellulosic biomass from wheat straw materials. Renew. Sustain. Energy Rev. 2018, 91, 483–489. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef]
- Ma, J.; Zhao, Q.B.; Laurens, L.L.M.; Jarvis, E.E.; Nagle, N.J.; Chen, S.; Frear, C.S. Mechanism, kinetics and microbiology of inhibition caused by long-chain fatty acids in anaerobic digestion of algal biomass. Biotechnol. Biofuels. 2015, 8, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sárvári Horváth, I.; Castillo, M.D.; Schnürer, A.; Agnihotri, S.; Ylitervo, P.; Edström, M. Utilization of Straw Pellets and Briquettes as Co-Substrates at Biogas Plants. Energiforsk Rep. 2017, 2017, 1–62. [Google Scholar]
- Li, L.; He, Q.; Wei, Y.; He, Q.; Peng, X. Early warning indicators for monitoring the process failure of anaerobic digestion system of food waste. Bioresour. Technol. 2014, 171, 491–494. [Google Scholar] [CrossRef]
- Sapmaz, T. Anaerobic Digestion of Organic Wastes with High Nitrogen Contents: An Investigation on Biogas Potential and Microbial Communities. Master’s Thesis, Istanbul Technical University, Istanbul, Turkey, 2017. (In Turkish). [Google Scholar]
- Sluiter, A.; Hames, B.; Hyman, D.; Payne, C.; Ruiz, R.; Scarlata, C.; Wolfe, J. Determination of Total Solids in Biomass and Total Dissolved Solids in Liquid Process Samples. Natl. Renew. Energy Lab. 2008, 9, 1–6. [Google Scholar]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA Analysis. Nucl. Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA GeneDatabase and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanzadeh, M.; Hagen, L.H.; Svensson, K.; Linjordet, R.; Horn, S.J. Biogas production from food waste via co-digestion and digestion- effects on performance and microbial ecology. Sci. Rep. 2017, 7, 17664. [Google Scholar] [CrossRef] [Green Version]
- Murto, M.; Björnsson, L.; Mattiasson, B. Impact of food industrial waste on anaerobic co-digestion of sewage sludge and pig manure. J. Environ. Manag. 2004, 70, 101–107. [Google Scholar] [CrossRef]
- Rozzi, A. Alkalinity considerations with respect to anaerobic digesters. Meded. Fac. Landbouwwet. Rijksuniv. Gent. 1991, 56, 1499–1514. [Google Scholar]
- Ferguson, R.M.W.; Coulon, F.; Villa, R. Organic loading rate: A promising microbial management tool in anaerobic digestion. Water Res. 2016, 100, 348–356. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Yang, Z.-H.; Zheng, Y.; Liu, J.-B.; Xiong, W.-P.; Zhang, Y.-R.; Lu, Y.; Xue, W.-J.; Fan, C.-Z. Organic loading rate and hydraulic retention time shape distinct ecological networks of anaerobic digestion related microbiome. Bioresour. Technol. 2018, 262, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Ziganshina, E.E.; Belostotskiy, D.E.; Shushlyaev, R.V.; Miluykov, V.A.; Vankov, P.Y.; Ziganshin, A.M. Microbial community diversity in anaerobic reactors digesting turkey, chicken, and swine wastes. J Microbiol Biotechnol. 2014, 24, 1464–1772. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Zhou, S.; Luo, R.; Gesang, Z.; Suolang, S. Metagenomic insights into the diversity of carbohydrate-degrading enzymes in the yak fecal microbial community. BMC Microbiol. 2020, 20, 302. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, H.; Zhang, Y.Y.; Si, D.D.; Chen, Q.W. Evolution of microbial community along with increasing solid concentration during high-solids anaerobic digestion of sewage sludge. Bioresour. Technol. 2016, 216, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennehy, C.; Lawlor, P.G.; Gardiner, G.E.; Jiang, Y.; Cormican, P.; McCabe, M.S.; Zhan, X. Process stability and microbial community composition in pig manure and food waste anaerobic co-digesters operated at low HRTs. Front. Environ. Sci. Eng. 2017, 11, 4. [Google Scholar] [CrossRef]
- Venkiteshwaran, K.; Bocher, B.; Maki, J.; Zitomer, D. Relating Anaerobic Digestion Microbial Community and Process Function: Supplementary Issue: Water Microbiology. Microbiol. Insights 2016, 8, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Yang, C.; Zhang, T. Microbial effects of part-stream low-frequency ultrasonic pretreatment on sludge anaerobic digestion as revealed by high-throughput sequencing-based metagenomics and metatranscriptomics. Biotechnol. Biofuels 2018, 11, 47. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Tian, Z.; Liu, J.; Zhou, J.; Yan, Z.; Yong, X.; Jia, H.; Wu, X.; Wei, P. Biogas production and microbial community dynamics during the anaerobic digestion of rice straw at 39–50 °C: A pilot study. Energy Fuels 2018, 32, 5157–5163. [Google Scholar] [CrossRef]
- Liang, X.; Whitham, J.M.; Holwerda, E.K.; Shao, X.; Tian, L.; Wu, Y.-W.; Lombard, V.; Henrissat, B.; Klingeman, D.M.; Yang, Z.K.; et al. Development and characterization of stable anaerobic thermophilic methanogenic microbiomes fermenting switchgrass at decreasing residence times. Biotechnol. Biofuels 2018, 11, 243. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Rao, S.; Shen, Z.; Lee, P.K.H. Substrate induced emergence of differenct active bacterial and archaeal assemblages during biomethane production. Bioresour. Technol. 2013, 148, 517–524. [Google Scholar] [CrossRef]
- Marchandin, H.; Jumas-Bilak, E. The Family Veillonellaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Regueiro, L.; Lema, J.M.; Carballa, M. Key microbial communities steering the functioning of anaerobic digesters during hydraulic and organic overloading shocks. Bioresour. Technol. 2015, 197, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Lebedeva, E.V.; Alawi, M.; Maixner, F.; Jozsa, P.G.; Daims, H.; Spieck, E. Physiological and phylogenetic characterization of a novel lithoautotrophic nitrite-oxidizing bacterium,‘Candidatus Nitrospira bockiana’. Int. J. Syst. Evol. Microbiol. 2008, 58, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Pope, P.B.; Eijsink, V.G.H.; Schnürer, A. Characterization of microbial community structure during continuous anaerobic digestion of straw and cow manure. Microb. Biotechnol. 2015, 8, 815–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zealand, A.M.; Mei, R.; Papachristodoulou, P.; Roskilly, A.P.; Liu, W.T.; Graham, D.W. Microbial community composition and diversity in rice straw digestion bioreactors with and without dairy manure. Appl. Microbiol. Biotechnol. 2018, 102, 8599–8612. [Google Scholar] [CrossRef] [Green Version]
- Evans, P.N.; Boyd, J.A.; Leu, A.O.; Woodcroft, B.J.; Parks, D.H.; Hugenholtz, P.; Tyson, G.W. An evolving view of methane metabolism in the Archaea. Nat. Rev. Microbiol. 2019, 17, 219–232. [Google Scholar] [CrossRef]
- Nishio, N.; Nakashimada, Y. Manufacture of Biogas and Fertilizer from Solid Food Wastes by Means of Anaerobic Digestion. In Food Industry Wastes, 1st ed.; Hosseva, M., Webb, C., Eds.; Assessment and Recuperation of Commodities; Elsevier: Amsterdam, The Netherlands, 2013; Chapter 7; pp. 121–133. [Google Scholar] [CrossRef]
- Demirel, B.; Scherer, P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: A review. Rev. Environ. Sci. Bio/Technol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Monlau, F.; Sambusiti, C.; Barakat, A.; Quemeneur, M.; Trably, E.; Steyer, J.-P.; Carrèree, H. Do furanic and phenolic compounds of lignocellulosic and algae biomass hydrolyzate inhibit anaerobic mixed cultures? A comprehensive review. Biotechnol. Adv. 2014, 32, 934–951. [Google Scholar] [CrossRef]
- Lerm, S.; Kleyboecker, A.; Miethling-Graff, R.; Alawi, M.; Kasina, M.; Liebrich, M.; Würdemann, H. Archaeal community composition affects the function of anaerobic co-digesters in response to organic overload. Waste Manag. 2012, 32, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Karakashev, D.; Batstone, D.J.; Trably, E.; Angelidaki, I. Is methanogenic pathway from acetate in the absence of Methanosaetaceae. Appl. Environ. Microbiol. 2006, 72, 5138–5141. [Google Scholar] [CrossRef] [Green Version]
- Hori, T.; Haruta, S.; Sasaki, D.; Hanajima, D.; Ueno, Y.; Ogata, A.; Ishii, M.; Igarashi, Y. Reorganization of the bacterial and archaeal populations associated with organic loading conditions in a thermophilic anaerobic digester. J. Biosci. Bioeng. 2015, 119, 337–344. [Google Scholar] [CrossRef]
- Francisci, D.D.; Kougias, P.G.; Treu, L.; Campanaro, S.; Angelidaki, I. Microbial diversity and dynamicity of biogas reactors due to radical changes of feedstock composition. Bioresour. Technol. 2015, 176, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yang, J.; Yu, Q.; Yong, X.; Xie, X.; Zhang, L.; Wei, P.; Jia, H. Different organic loading rates on the biogas production during the anaerobic digestion of rice straw: A pilot study. Bioresour. Technol. 2017, 244, 865–871. [Google Scholar] [CrossRef] [PubMed]
- De Vrieze, J.; Hennebel, T.; Boon, N.; Verstraete, W. Methanosarcina: The rediscovered methanogen for heavy duty biomethanation. Bioresour. Technol. 2012, 112, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pap, B.; Györkei, Á.; Boboescu, I.Z.; Nagy, I.K.; Bíró, T.; Kondorosi, É.; Maróti, G. Temperature-dependent transformation of biogas-producing microbial communities points to the increased importance of hydrogentrophic methanogenesis under thermophilic operation. Bioresour. Technol. 2015, 177, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Zamanzadeh, M.; Hagen, L.H.; Svensson, K.; Linjordet, R.; Horn, S.J. Anaerobic digestion of food waste—Effect of recirculation and temperature on performance and microbiology. Water Res. 2016, 96, 246–254. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yangin-Gomec, C.; Agnihotri, S.; Ylitervo, P.; Sárvári Horváth, I. Assessment of Microbial Diversity during Thermophilic Anaerobic Co-Digestion for an Effective Valorization of Food Waste and Wheat Straw. Energies 2023, 16, 55. https://0-doi-org.brum.beds.ac.uk/10.3390/en16010055
Yangin-Gomec C, Agnihotri S, Ylitervo P, Sárvári Horváth I. Assessment of Microbial Diversity during Thermophilic Anaerobic Co-Digestion for an Effective Valorization of Food Waste and Wheat Straw. Energies. 2023; 16(1):55. https://0-doi-org.brum.beds.ac.uk/10.3390/en16010055
Chicago/Turabian StyleYangin-Gomec, Cigdem, Swarnima Agnihotri, Päivi Ylitervo, and Ilona Sárvári Horváth. 2023. "Assessment of Microbial Diversity during Thermophilic Anaerobic Co-Digestion for an Effective Valorization of Food Waste and Wheat Straw" Energies 16, no. 1: 55. https://0-doi-org.brum.beds.ac.uk/10.3390/en16010055