1. Introduction
Loss of tooth dentin not only generates unpleasant pain but ultimately leads to the weakening of whole tooth stability due to reduced dentin thickness. Among its varying causes, dental caries is the most prevalent chronic disease in both children and adults, affecting more than 5 billion people worldwide. Dental caries is caused by bacterial infection and resulting acidic byproducts that demineralize the tooth. Depending on the extent of the infection, the carious lesion either stops at the enamel or penetrates into the dentin [
1]. Exposed dentinal tubules in dental caries serve as a route for bacterial invasion that may result in various pulpal responses [
2]. In severe cases, tooth vitality is permanently lost [
3]. A healthy tooth is composed of nearly 70% dentin enclosing the entire dental pulp, which is a pool of diverse stem cells [
4]. Loss of tooth dentin not only generates unpleasant sensitivity but ultimately leads to the weakening of whole tooth stability due to reduced dentin thickness. Accordingly, incessant efforts have been made to regenerate biologic dentin. However, most of the previously reported reparative tertiary dentin shows features of osteodentin, which is composed of bone-like structures with osteocyte-resembling cells entrapped in lacunae-like spaces [
5]. The formation of osteodentin remains valuable because the damaged tissue is repaired with a newly mineralized barrier. Nonetheless, many reports highlight the significance of dentinal tubules in terms of defense mechanisms [
6,
7,
8]. Moreover, the mineral density of osteodentin is less than that of physiologic tubular dentin [
9]. Therefore, the identification of biological materials capable of restoring dentin with tubular architecture is clinically important.
Based on the knowledge that epithelial–mesenchymal interaction is essential for tooth development, we previously discovered a dental epithelium-derived protein called Copine 7 (CPNE7). CPNE7 is an evolutionarily conserved, calcium-dependent phospholipid-binding protein which consists of two C2 domains in the N terminus and a von Willebrand factor A domain in the C terminus [
10]. Secreted from pre-ameloblasts, CPNE7 induces odontoblast differentiation in vitro and promotes dentin formation ex vivo [
11,
12,
13]. As a result, CPNE7 was suggested as a new molecule with the potential to diffuse across the dentin and induce tertiary dentinogenesis [
14]. The fact that recombinant CPNE7 is a cell-derived soluble bioactive molecule makes it a promising candidate for use in regenerative dental medicine.
As no peptide has yet been reported to form physiologic tubular dentin by stimulating odontoblasts or odontoblastic differentiation, we sought to design and synthesize a stable, cost-efficient, and manipulative functional peptide that mimics the functions of the protein CPNE7. Here, we thoroughly evaluate the tubular dentin forming capacity of CPNE7-derived oligopeptide (Cpne7-DP) and suggest their use as future therapeutic agents for dentinal defects such as dental caries.
2. Materials and Methods
2.1. Tooth Defect Models with Canine Teeth
For in vivo canine studies, a total of 10 beagle dogs (2 y) were operated on, with all of them reaching end points. For each animal, 4 to 6 maxillary premolars and 6 mandibular premolars were used, depending on their periodontal conditions. Control and experimental groups were allocated to each and every animal so that possible biases resulting from individual nutritional, behavioral, and pulpal conditions could be eliminated. Compared to small animal studies, the sample size was necessarily limited for feasibility and ethical reasons. While the exact numbers for each experiment are included in the figure legends, at least 6 premolars and up to 12 premolars were assigned to every control and experimental group. Investigators performing beagle operation and sampling were not blinded, while investigators involved in histological analysis were blinded. All experiments using animals followed protocols approved by the Institutional Animal Care and Use Committee of Seoul National University (SNU-180416-2-1 and SNU-171020-5-2).
Beagle dogs of 1–2 years of age were used for three independent experiments. After disinfecting the cervical regions of the maxillary and mandibular premolars with 0.5% chlorhexidine, dish-shaped class V cavities were prepared using a #4 high-speed round bur (head diameter, 1.4 mm). For shallow cavities, the drilling stopped when half to two-thirds of the bur head penetrated the tooth structure, depending on the tooth size. For deep cavities, the drilling stopped when the color of the remaining dentin looked reddish gray. The smear layer was removed with a gentle application of 17% EDTA. After sufficient irrigation, surgical sites were dried with a cotton pellet. For CPNE7 function analysis, the cavities were either untreated or treated with a topical application of rCPNE7 (recombinant CPNE7) protein (1 µg total rCPNE7 per tooth in a buffer containing 25 mM Tris-HCl, 100 mM glycine, and 10% glycerol). After a brief incubation for diffusion, the cavities were then filled with either glass ionomer (GI) cement (GC; Fuji II LC; GC America Inc., Alsip, IL, USA), composite resin (Filtek Supreme Ultra Flowable Restorative; 3 M, MN, USA) or intermediate restorative material (IRM; Dentsply Sirona, New York, PA, USA). For peptide function analysis, an HIV-1 Tat-derived cell-penetrating peptide was used as control. In addition to the abovementioned shallow and deep cavity models, complete pulp exposure was performed for peptide analysis. For complete pulp exposure, the drilling stopped when a pin-point exposure of the pulp was generated and bleeding was evident. Cotton pellets soaked in saline were used for bleeding control. Cavities with exposed pulps were divided into two groups for the experiment. Group 1 received only GI cement filling, group 2 received GI cement filling after MTA (ProRoot MTA; Dentsply Sirona, New York, PA, USA) sealing, and group 3 received GI cement filling after MTA mixed with CPNE7-derived peptide (Cpne7-DP) sealing.
The premolar areas were dissected 3, 6, or 12 weeks following surgery, and samples were immersed in 4% paraformaldehyde and kept for an additional 24 h at 4 °C. Decalcification was performed in 10% formic acid, and the specimens were then embedded in paraffin. Serial 5-µm-thick sections were H&E stained.
2.2. TUNEL Assay
Apoptotic cells were detected using the TUNEL kit (Roche Biochemicals, Basel, Switzerland), according to the manufacturer’s instructions. Endogenous peroxidase activity within the tissue sections were blocked in 3% H2O2 prior to enzymatic labeling. To yield a colored reaction product, enzymatically labeled cells were then incubated with 3,3-diaminobenzidine tetrahydrochloride. Visualization was achieved by incubating the sections with diaminobenzidine tetrahydrochloride (DAB), and subsequent counter-staining with hematoxylin. Positive TUNEL signals were converted to red color using IHC Profiler of Image J software (National Institute of Health, Gaithersburg, MD, USA).
2.3. Cell Culture
MDPC-23 cells were provided by Dr. J.E. Nor (University of Michigan, Ann Arbor, MI, USA) and cultured in Dulbecco’s modified Eagle medium (DMEM; Gibco BRL., Carlsbad, CA, USA). C3H10T1/2 cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and cultured in RPMI 1640 medium (Gibco BRL). Both cell lines were supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco BRL) and antibiotic-antimycotic reagents (Gibco BRL) at 37 °C in a 5% CO
2 atmosphere. Impacted human third molars from patients between the ages of 18 and 22 were provided by the Seoul National University Dental Hospital. The experimental protocol was approved by the Institutional Review Board (IRB No: S-D20140007). Informed consent was obtained from every patient. Isolation of whole pulp cells was performed as previously described [
11], and cells were cultured in minimum essential media α (MEM-α; Gibco BRL) for use in in vitro and ex vivo experiments. For human dental pulp cells (hDPCs) and MDPC-23 cell differentiation, 80–90% confluent cells were cultured in corresponding media supplemented with 5% FBS, ascorbic acid (50 µg/mL), and β-glycerophosphate (10 mM) for up to 3 weeks. Passages from 2 to 4 were used for hDPSCs, and 23 to 25 were used for MDPC-23 cells.
2.4. Peptide Synthesis
Cpne7-DP consists of a synthetic peptide corresponding to the 10 amino acid residue 344–353 fragment (KYKQKRRSYK) of the hCPNE7 protein. The peptides were synthesized using the Fmoc (9-fluorenylmethoxycarbonyl)-based solid-phase method and characterized by Lugen Sci. Co., Ltd. (Bucheon, Korea). The purity of the peptides used in this study was greater than 97%, as determined by high-performance liquid chromatography.
2.5. Luciferase Assay
MDPC-23 cells were seeded on a 24-well plate at a density of 5 × 104 cells/wells and transfected with Lipofectamine Plus™ reagent (Invitrogen, Carlsbad, CA, USA) after 24 h. For each transfection, 0.4 µg luciferase reporter plasmid pGL3basic (Control) and pGL3LUC dspp (−750~61) were used. Transfected cells were treated with Cpne7-DP or recombinant CPNE7 protein for 48 h and were then lysed for luciferase activity assessment using the luciferase reporter gene assay system (Roche Applied Science, Indianapolis, IN, USA) according to the manufacturer’s instructions. The measurements were performed with a luminometer (FLUOStar OPTIMA, BMC Laboratory, Offenburg, Germany). Three independent experiments with triplicate samples were analyzed.
2.6. Cytotoxicity Assessment
To evaluate the effects of Cpne7-DP on hDPCs proliferation, an MTT [3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, a tetrazole] assay (Sigma-Aldrich, Saint Louis, MO, USA) was performed. Cells were seeded on a 96-well plate at a density of 3 × 103 cells/well and treated with Cpne7-DP after 24 h. Cells were cultured for up to 5 days, and MTT analyses were performed on days 0, 1, 3, and 5. After washing with phosphate-buffered saline (PBS), 20 µL of MTT was added to each well and incubated for 4 h at 37 °C. After removing the MTT solution, the converted dye was dissolved in Me2SO and measured by reading the absorbance at a wavelength of 540 nm with a microplate reader (Multiskan EX; Thermo Electron Corp., Waltham, MA, USA). Three independent experiments with triplicate samples were analyzed.
2.7. Real-Time Polymerase Chain Reaction Analysis
Total RNA was extracted from cells with TRIzol reagent according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). Three µg of RNA was reverse transcribed using Superscript III reverse transcriptase (Invitrogen) and oligo (dT) primers (Invtrogen). One µL of cDNA was subjected to PCR amplification using the ABI PRISM 7500 sequence detection system (Applied Biosystems, Carlsbad, CA, USA) using SYBR Green PCR Master Mix (Applied Biosystems) according to the manufacturer’s instructions. PCR was performed using the following conditions: 94 °C for 1 min, 95 °C for 15 s, and 60 °C for 34 s, for 40 cycles. All reactions were performed in triplicate and normalized to reactions using the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Relative changes in gene expression were calculated using the comparative threshold cycle (CT) method. The sequences of the real-time PCR primers used in the study are listed in
Table 1.
2.8. Western Blot Analysis
Whole cell lysates were harvested in a lysis buffer consisting of 50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% Nonidet P-40, 1 mM EDTA, and 1 mM PMSF supplemented with protease inhibitors (Roche Molecular Biochemicals, Mannheim, Germany). The supernatant after centrifugation at 13,000×
g for 30 min was collected for analysis. DCTM protein assay system (Bio-Rad Laboratories, Hercules, CA, USA) was used to measure protein concentrations. Twenty-five μg of proteins were resolved using 10% polyacrylamide gel electrophoresis and transferred to a PVDF membrane, which was then blocked with PBST (10 mM phosphate-buffered saline, pH 7.0, and 0.1% Tween-20) buffer containing 5% non-fat dry milk for 1 h at room temperature. After washing, the blots were incubated with primary antibodies indicated in
Table 2, overnight at 4 °C with gentle shaking. Before incubation with anti-rabbit or anti-mouse immunoglobulin G conjugated to horseradish peroxidase in PBST for 1 h at room temperature, blots were washed 3 times for 10 min in PBST. Labeled protein bands were detected under an enhanced chemi-luminescence reagent (ECL; Santa Cruz Biotechnology, Dallas, TX, USA) according to the manufacturer’s guidelines. Semi-quantitative analyses were performed using Image J software (National Institute of Health).
2.9. Ex Vivo Transplantation and Histological Analysis
Human root segments (<5 mm in thickness) were prepared from extracted human teeth after pulp tissues were removed according to our previously reported protocol [
15]. Human DPCs (2 × 10
6) were mixed with hydroxy apatite/tricalcium phosphate (HA/TCP) ceramic powder (Zimmer, Warsaw, IN, USA) alone or with Cpne7-DP (10 µg) in an 0.5% fibrin gel and then transplanted subcutaneously into immunocompromised mice (NIH-bg-nu-xid; Harlan Laboratories, Indianapolis, IN, USA) for 6 or 12 weeks. Dentin/pulp-like tissue formation in the emptied root canal space was evaluated after mixing hDPCs (2 × 10
6) with Cpne7-DP (10 µg) in a 0.5% fibrin gel and inserting the mixture into emptied root canal space of the human root segments for 6 weeks. Harvested samples were fixed in 4% in paraformaldehyde, decalcified in 10% EDTA (pH 7.4), embedded in paraffin, and stained with hematoxylin and eosin (H&E) (Vector Laboratory, Burlingame, CA, USA), Masson’s Trichrome Stain (Polysciences, Warrington, PA, USA), or processed for immunohistochemical analysis. For immunohistochemistry, the sections were incubated overnight at 4 °C with rabbit polyclonal DSP and BSP produced, as described previously [
16] at a dilution of 1:150. Biotin-labeled goat anti-rabbit IgG (Vector Laboratory) was incubated with the sections at room temperature for 30 min, which were then reacted with the avidin-biotin-peroxidase complex (Vector Laboratory). Signals were converted using a diaminobenzidine kit (Vector Laboratory). Nuclei were stained with hematoxylin.
2.10. Immunofluorescence Staining
For peptide translocation and localization analysis, a Cy5 Fast Conjugation Kit (ab188288, Abcam, Cambridge, MA, USA) was used to tag Cpne7-DP according to the manufacturer’s instructions. MDPC-23 cells were treated with Cy5-labeled Cpne7-DP (10 µg/mL) and fixed 3 h later. Before fixing with 4% paraformaldehyde in PBS, cells in Laboratory-Tek chamber slides (Nunc, Rochester, NY, USA) were washed with PBS. Cells were then visualized using fluorescence microscopy (AX70, Olympus, Tokyo, Japan). DAPI (Sigma-Aldrich) was used to identify the chromosomal DNA in the nucleus (1:1000 dilution). Reagents used are listed in
Table 3.
2.11. Transient Transfection
C3H10T1/2 cells or hDPCs were seeded on 60 mm culture plates at a density of 1.0 × 106 cells. The cells were transiently transfected with DDK (Flag)-tagged CPNE7 using the Metafectene Pro reagent (Biontex Laboratories GmbH, Munich, Germany) according to the manufacturer’s instructions.
2.12. Scanning Electron Microscopic Analysis
Samples were fixed in 0.1 M cacodylate buffer (pH 7.3) containing 2.5% glutaraldehyde for 30 min and in 0.1 M cacodylate buffer (pH 7.4) containing 1% osmium tetroxide for 1 h. Samples were then rapidly dehydrated through an ethanol gradient. After sputter coating with gold, samples were observed under a scanning electron microscope (S-4700, HITACHI, Tokyo, Japan).
2.13. Specimen Preparation
Human third molars were collected at the Seoul National University Dental Hospital (Seoul, Korea), and the experimental protocol was approved by the Institutional Review Board (IRB No: S-D20140007). Teeth were decoronated to 2–3 mm using a safe diamond disc (Isomet, Buehler Ltd., Düsseldorf, Germany) to eliminate the coronal enamel layer and expose the dentin surface. Teeth were then treated with 5 mL of 32% phosphoric acid solution for 5 min to completely open the dentinal tubules. To remove the residual smear layer, specimens were ultrasonicated in deionized water twice for 5 min each and then rinsed with PBS three times.
2.14. CLSM Specimen Preparation and Analyses
Cpne7-DP was tagged with Rhodamin B dye using the Rhodamine Fast Conjugation kit (ab188286, Abcam, Cambridge, MA, USA) according to the manufacturer’s instructions. Rhodamin B-labeled Cpne7-DP (20 µg) was applied to the dentin surfaces of specimens with the help of disposable microbrushes for 1 min, and then specimens were washed with PBS three times. Subsequently, the specimens were longitudinally sectioned at a thickness of 0.5 mm using a safe diamond disc. The sections were mounted onto glass slides and scanned under a confocal laser scanning microscope (LSM 700; Carl Zeiss, Jena, Germany).
2.15. Alizarin Red S Staining
MDPC-23 cells were seeded on 60 mm culture plates at a density of 1.0 × 106 cells and cultured in differentiation medium for 14 days with or without Cpne7-DP. The formation of mineralized nodules was evaluated by staining with alizarin red S (Sigma-Aldrich) solution in 0.1% NH4OH at pH 4.2 for 20 min at room temperature.
2.16. Microleakage Test
The extent of tubule occlusion was measured with the previously reported nano-fluid movement measuring device [
17]. Apical 3 mm of extracted beagle incisors was cut with a high-speed diamond bur (TF-13, MANI, Tokyo, Japan) to expose the root canals. A 0.9 mm metal tube was inserted, while 32% phosphoric acid, adhesive agent (Singlebond Universal, 3M ESPE, St Paul, MN, USA) and flowable composite resin (Filtek Supreme Ultra Flowable Restorative; 3M, Alexandria, MN, USA) were applied to bond with the metal tube. All areas except for the defect region were covered with nail varnish several times. The prepared specimens were kept in distilled water. A nano-fluid movement measuring device (NanoFlow, IB Systems, Seoul, Korea) recognizes the movement of bubbles due to leakage when distilled water is left to flow from the tooth apex to the exposed dentin at 70 cm H
2O. All measurements were taken 40 min after the specimens were connected, while the first 20 min of outflow was excluded.
2.17. Statistical Analysis
All data were expressed as the mean ± standard deviation of triplicate experiments. Statistical significance was analyzed using a non-parametric Mann–Whitney U test for comparisons between two groups, and one-way analysis of variance (ANOVA) with Bonferroni correction for comparisons between more than two groups by the SPSS software version 25. p values less than 0.05 were considered statistically significant.
4. Discussion
Dentin has a number of similarities with bone both in its chemical composition and its mode of formation; however, it does not undergo lifelong remodeling and cannot be replaced after loss [
19,
20]. Odontoblasts are ectomesenchyme-derived post-mitotic cells that are responsible for dentin development and eventually line dentin [
21]. Although odontoblasts have recently been reported to have sensory and immune cell capacities [
22,
23], their primary role is the secretion of the organic matrix that will be progressively mineralized [
24]. Noticeably, odontoblasts elongate their cellular processes as they secrete and form predentin. These odontoblastic processes ultimately get embedded in the mineralized dentin matrix, establishing the dentinal tubule structure [
25]. The secretory activity of a mature odontoblast that has completed primary and secondary dentin formation diminishes under healthy conditions. These resting state odontoblasts typically shrink in size and accumulate lipofuscin due to decreased autophagic activity [
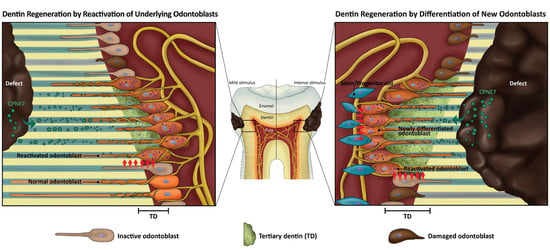
26]. In response to dentinal defects, either underlying odontoblasts are reactivated to generate physiologic reactionary dentin or other pulp cell sources are recruited and differentiate into odontoblast-like cells to produce pathologic reparative dentin [
27]. These endogenous processes may be a key to regenerative medicine for tooth dentin.
Shallow and mild injuries to dentin stimulate the matrix-secreting activity of underlying resting state odontoblasts. The newly formed tertiary dentin is continuous with remaining dentin through the dentinal tubule structure and is called “reactionary” dentin [
27]. In the case of deep and severe dentin injuries, underlying odontoblasts partially or massively undergo cell apoptosis, and a sub-odontoblastic population is recruited to differentiate into odontoblast-like cells [
28,
29,
30]. Whether these pulp cells have really differentiated into odontoblasts can be examined by the morphology of the newly generated mineralized tissue called “reparative dentin.” In the present study, treatment of CPNE7 or its derivative peptide Cpne7-DP resulted in the regeneration of tubular dentin in both the shallow and deep cavity models, indicating their abilities to not only induce new odontoblast differentiation, but to reactivate the matrix-secretion of underlying odontoblasts.
In addition to causing hypersensitivity, exposed dentinal tubules can be a route for the invasion of foreign substances [
31]. The current treatment for dentin hypersensitivity patients incorporates insoluble precipitate formation in the open tubules [
32,
33]. The ability of Cpne7-DP to stimulate underlying odontoblasts to resume matrix-secreting activity results in peritubular dentin formation. In addition to the newly generated tubular dentin along the pulp side, our SEM analysis revealed that peritubular dentin is deposited above the odontoblast process near the dentin-restoration interface. Consequently, it resulted in the physiologic occlusion of dentinal tubules. Integrity of the tubule occlusion was further confirmed by a nano-fluid movement test.
A number of synthetic peptides, especially those derived from proteins in an enamel or dentin matrix, have been developed to promote remineralization of enamel and dentin [
34,
35,
36,
37]. Most were capable of inducing hydroxyapatite deposition, and a DSP-derived peptide was reported to act on pulp cells and induce osteodentin formation [
38]. In the present study, Cpne7-DP highly reproduced the in vitro effects of CPNE7 by upregulating odontoblast marker genes,
DSPP, and
Nestin. Moreover, subcutaneous transplantation of Cpne7-DP-treated hDPCs resulted in the formation of a dentin-pulp-like complex ex vivo. Unlike CPNE7, which is internalized via nucleolin-mediated endocytosis, Cpne7-DP seems to act on odontoblasts by directly penetrating the cell membrane. Exactly how odontoblast marker gene transcription is regulated by Cpne7-DP is an area of future exploration.
Although the efficiency of hard tissue regeneration differs in model animals, regeneration studies in large mammals (e.g., dogs) have been reported to be more challenging than in small animals (e.g., mice) [
39,
40]. Nevertheless, large mammals provide a more appropriate model for mimicking human disease and are thus more transferable to a human model. In our canine model, both CPNE7 and Cpne7-DP showed successful regeneration of tubular dentin. As the results were based on a single topical application, the effects of multiple repetitive applications should also be analyzed to determine their additive effects. The thickness of the newly formed tubular dentin increased with time, and 6-week samples demonstrated greater new dentin volume than 3-week samples. Such correlation implies that the secretory activity of either newly differentiated odontoblast-like cells or existing odontoblasts is prolonged once triggered. Nonetheless, one of the limitations of this study includes the lack of an exact quantification of newly formed tertiary dentin.
The findings of the current study suggest that Cpne7-DP promotes the formation of new tubular dentin by inducing odontoblast differentiation of dental pulp stem cells in both dentin and pulp exposure models (
Figure 6,
Table 4). The development of Cpne7-DP, a synthetic oligopeptide derived from CPNE7, proved to be advantageous not only in that it perfectly reproduces the functions of CPNE7, but it is a more stable and potent cell-penetrating peptide. Coherent observations in in vitro, ex vivo, and in vivo models establish the groundwork for the clinical translation of Cpne7-DP, which shows a promising ability to arrest the demineralization process, compensate for lost dentin in dentin defects, and occlude exposed dentinal tubules to reduce dentin hypersensitivity.

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}