Modelling the Incursion and Spread of a Forestry Pest: Case Study of Monochamus alternatus Hope (Coleoptera: Cerambycidae) in Victoria


Abstract
:1. Introduction
2. Materials and Methods
- A high-priority pest for the forestry industry.
- An incursion of the pest would require an emergency response.
- Historical data of an incursion in a similar environment to Victoria.
- Sufficient information available on the life history and dispersal of the pest.
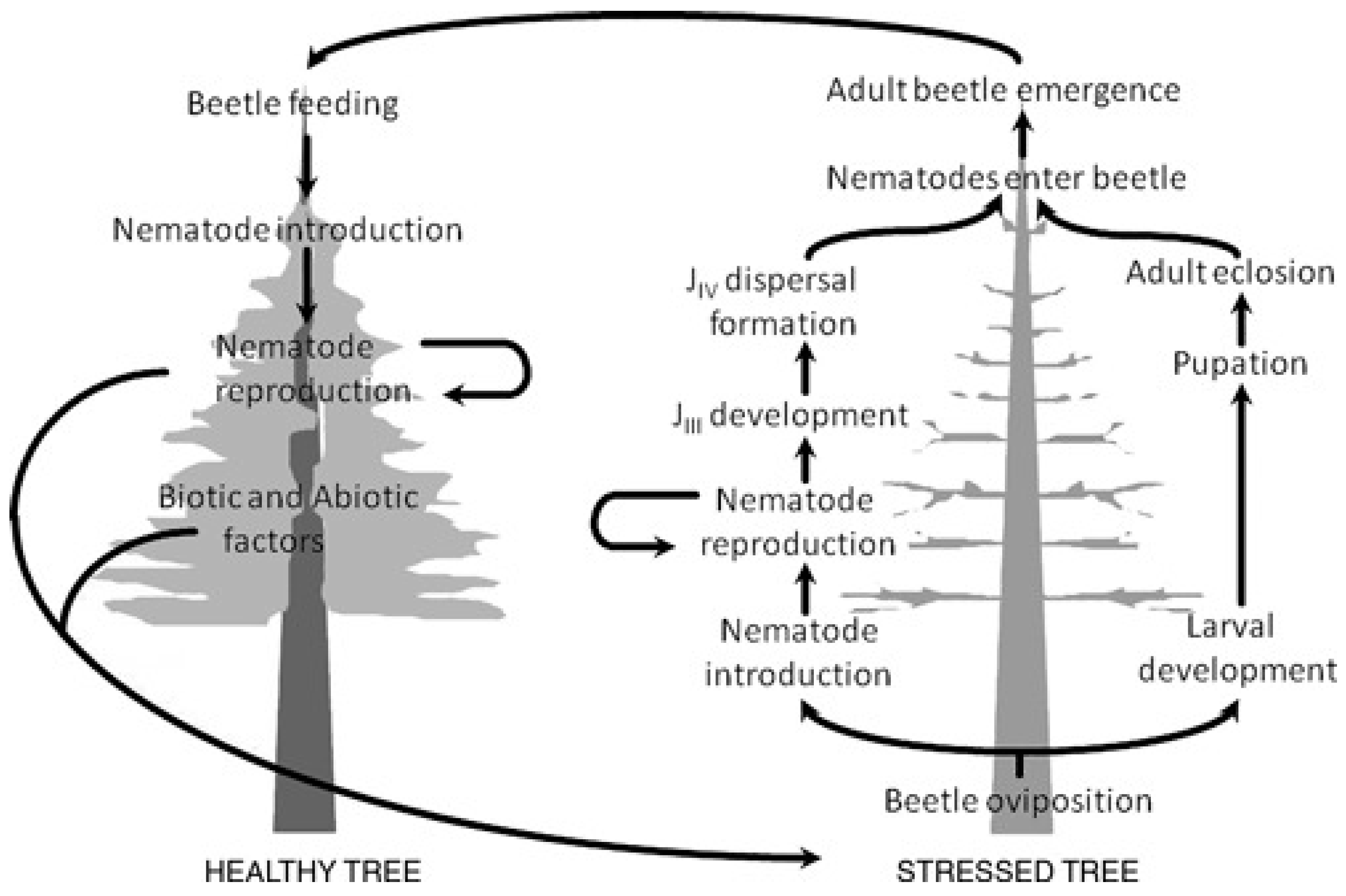
2.1. Life Cycle of Monochamus Species and Bursaphelenchus Xylophilus
2.2. Model Components
- They were considered important influences in the establishment and spread of pine beetles.
- The data were available at a scale that was suitable for inclusion in the model.
- (a)
- Pine beetle life cycle module which simulates the growth from eggs, through the larval stages, pupae and then to adult, dispersal of a proportion of adult beetles to suitable nearby trees, mating and egg production.
- (b)
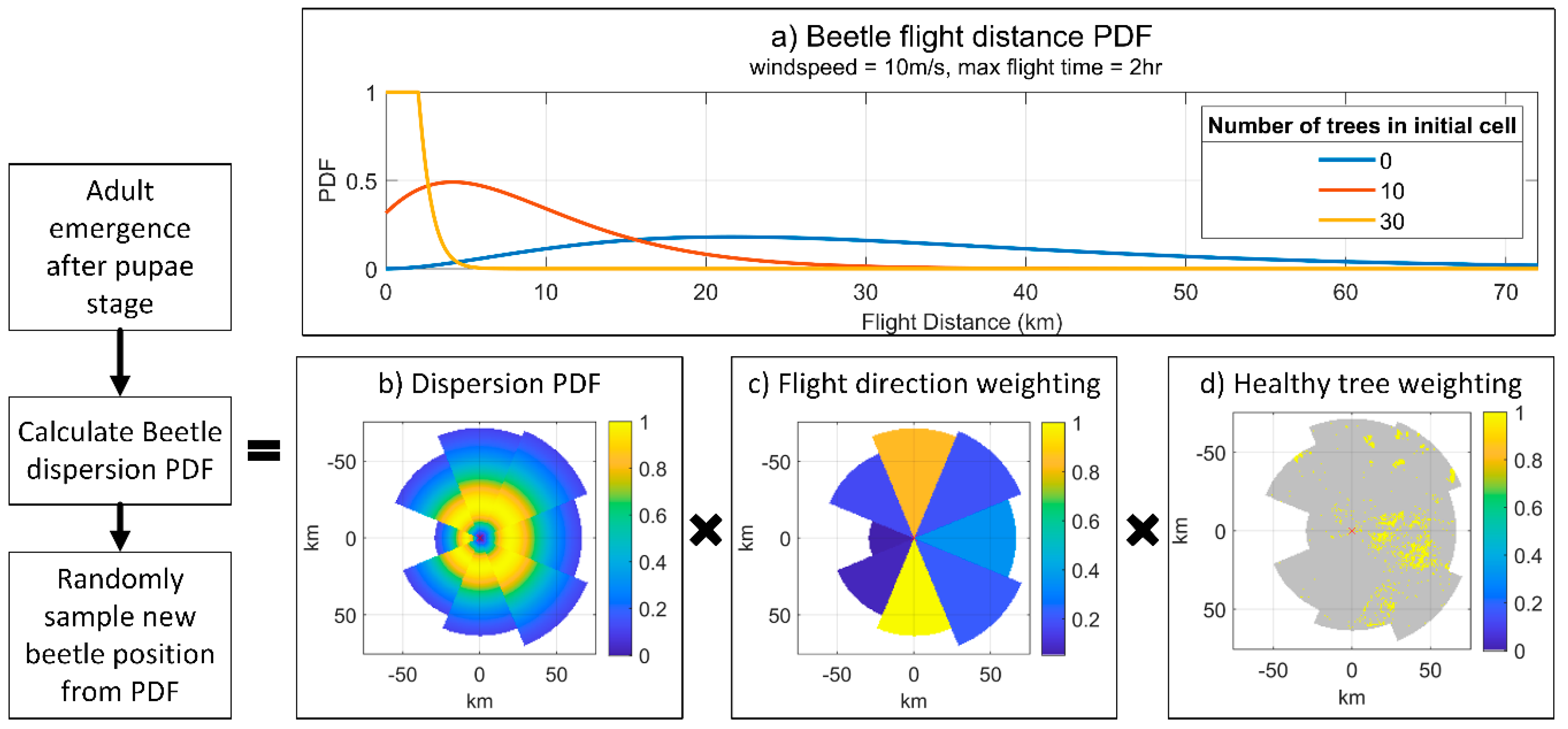
- Wind spread module that disperses a proportion of adult beetles across the landscape during spring and summer, depending on actual or averaged wind direction(s) and its interaction with the dispersal kernel.
- (c)
- Host availability and suitability module that accurately describes the distribution, abundance and health status of trees within the Pineacea family (pine, fir and spruce species), based on local, state and federal government datasets available across Victoria (see [16] for a full description of the host availability and suitability dataset development). Most of the local and state government pine survey data were of high quality and spatially accurate. All data sources were curated for data quality and duplicate records from the diverse data sources were removed. Each grid cell contains a count of healthy, healthy (but infected by the nematode), stressed (but uninfected by the nematode), stressed (after being infected by the nematode) and dead pine trees.
2.3. Model Design
- The number and health status of pine trees,
- Average meteorological wind data, relevant to the time frame of the model run,
- The presence, sex, and life stages of any beetles,
- The nematode infection status of any beetles, and
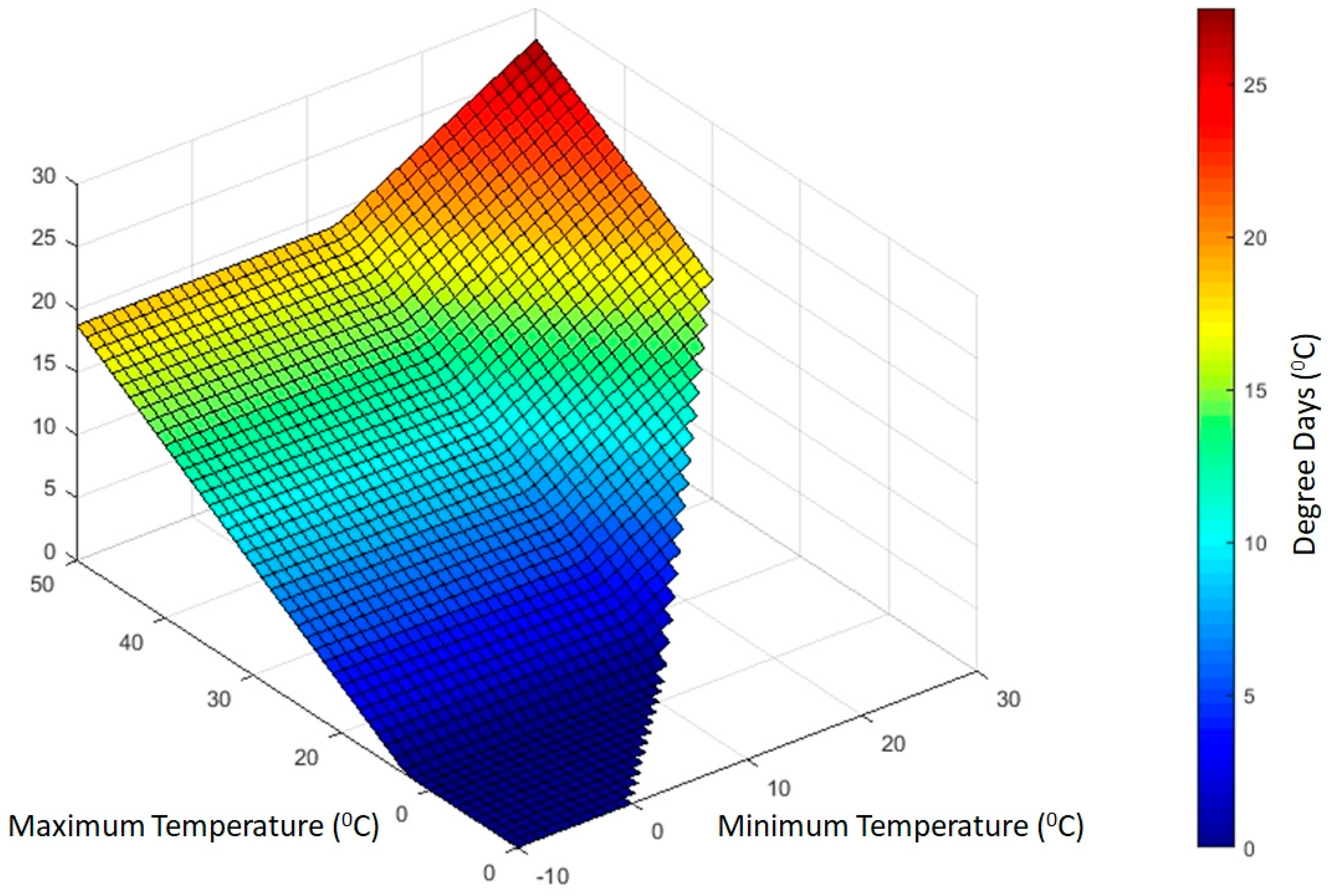
- Degree day information relevant for beetle development.
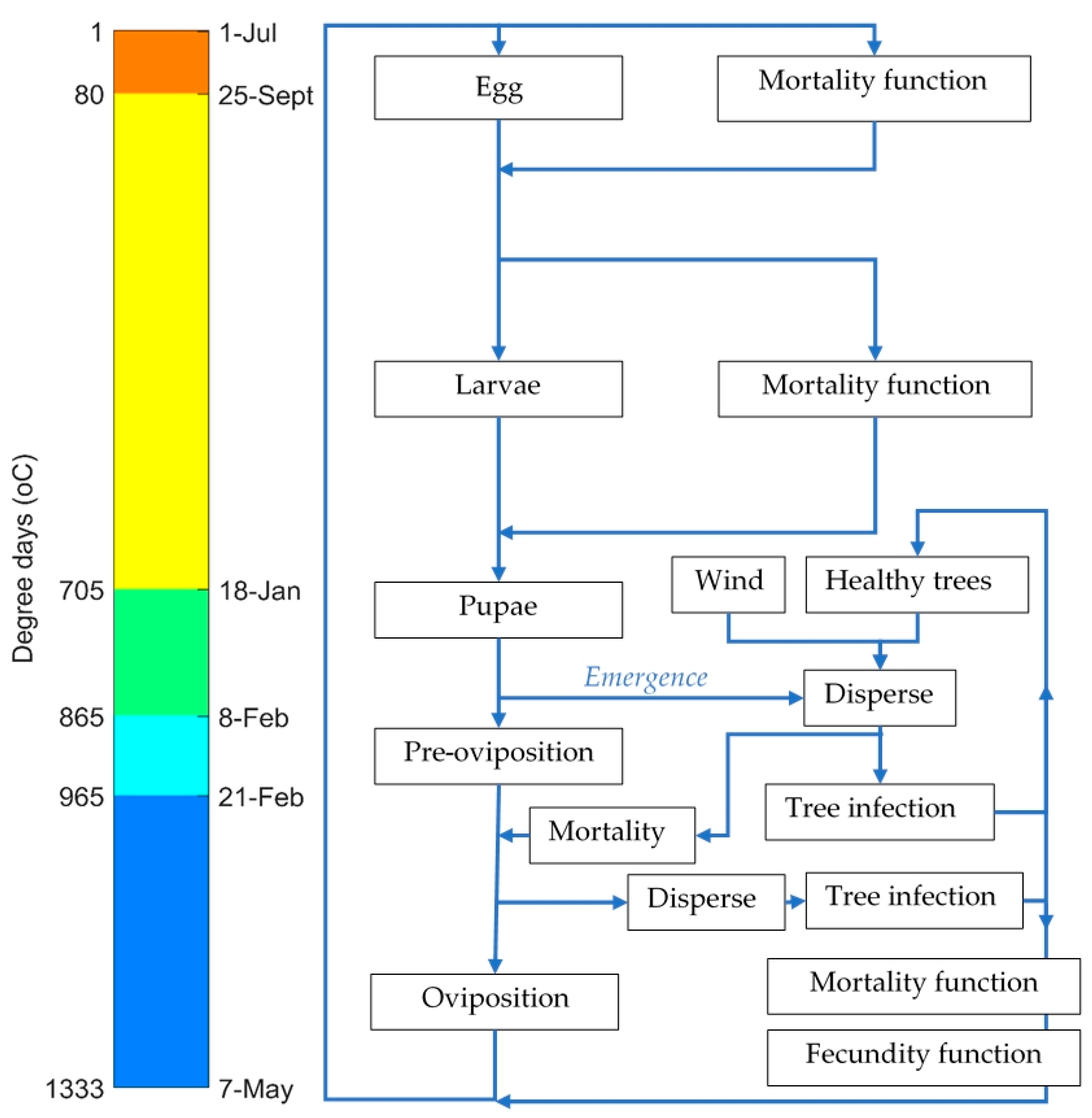
2.4. Life Cycle
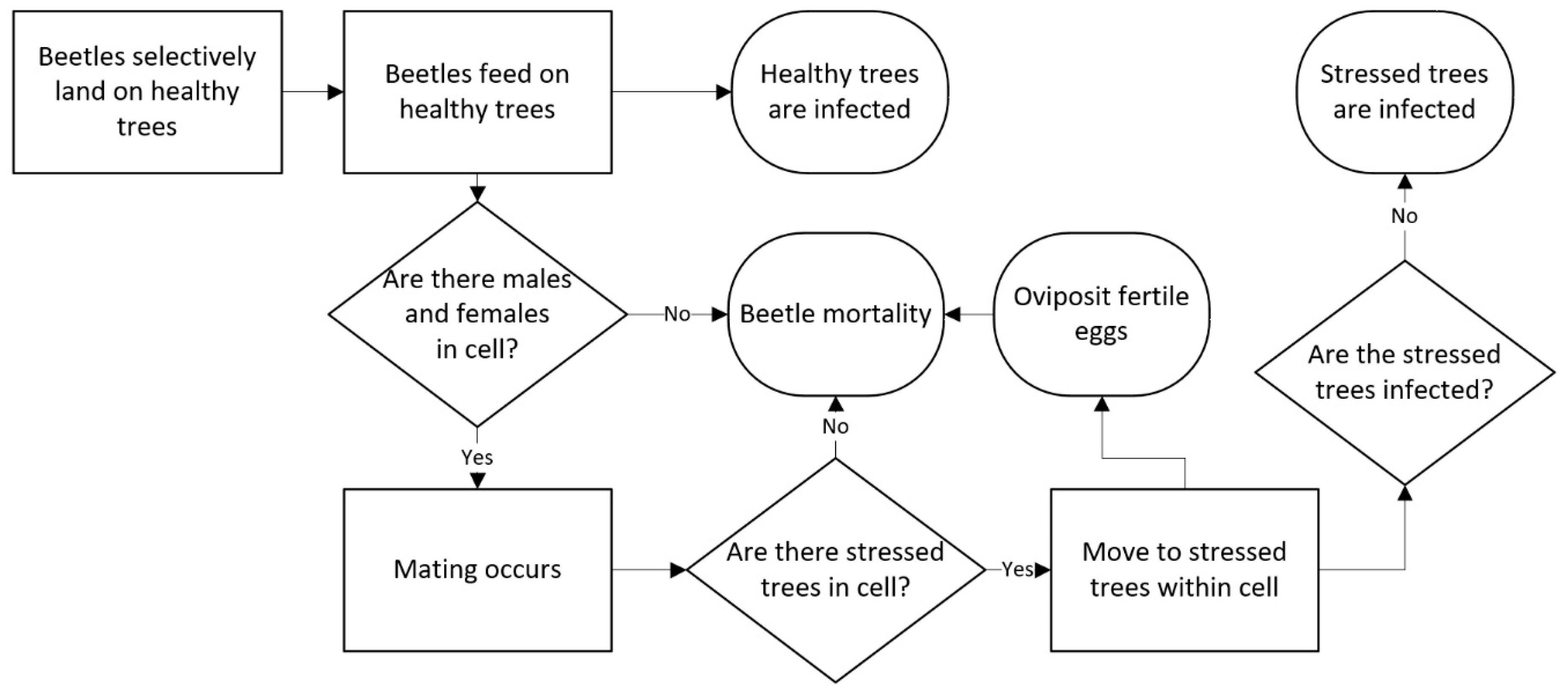
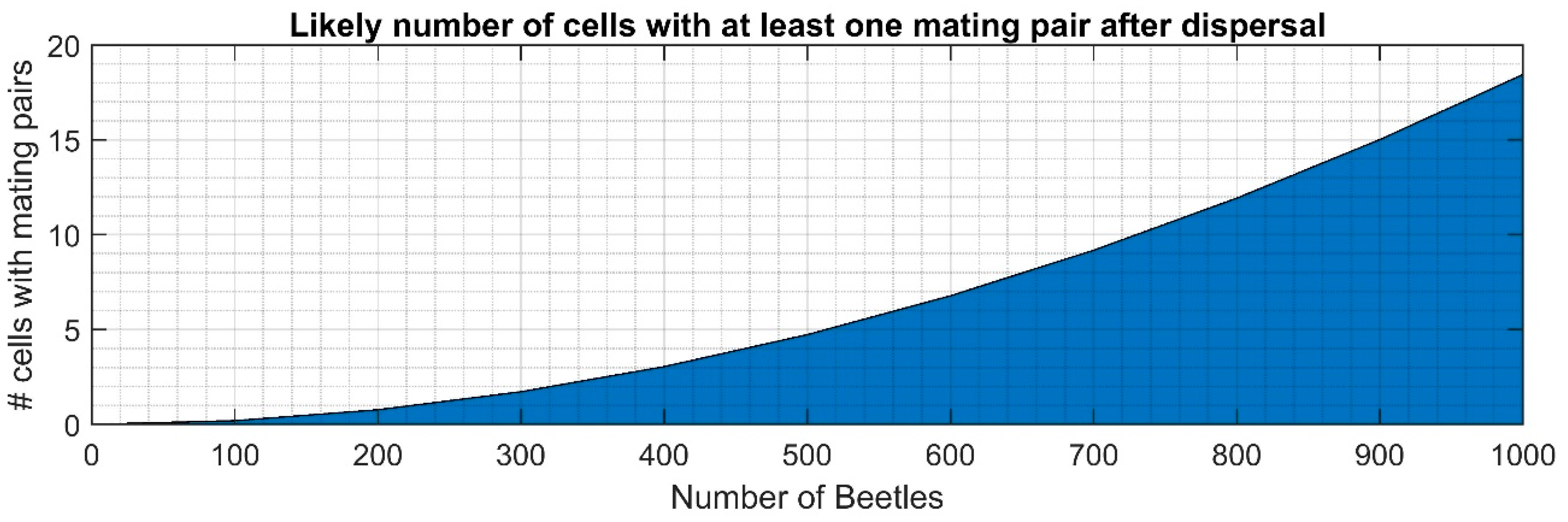
2.5. Feeding, Mating and Ovipositioning
2.6. Phenological Development
2.7. Host Availability and Suitability (Pine Model)
- Healthy: tree is growing robustly (this category is included in the initial input dataset and is based on observations recorded in source data).
- Healthy and infected by the nematode: tree is not showing signs of stress but has been infected by the nematode after beetle feeding (this category is updated following beetle dispersion, feeding and egg laying events within the model).
- Stressed but uninfected by the nematode: tree is showing signs of stress but has not been infected by the nematode (this category is included in the initial input dataset and is based on observations recorded in source data).
- Stressed and infected by the nematode: tree is showing signs of stress and has been infected by the nematode (this category is updated following beetle dispersion, feeding and egg laying events within the model).
- Dead: tree is dead but still present (this category is included in the initial input dataset and is based on observations recorded in source data).
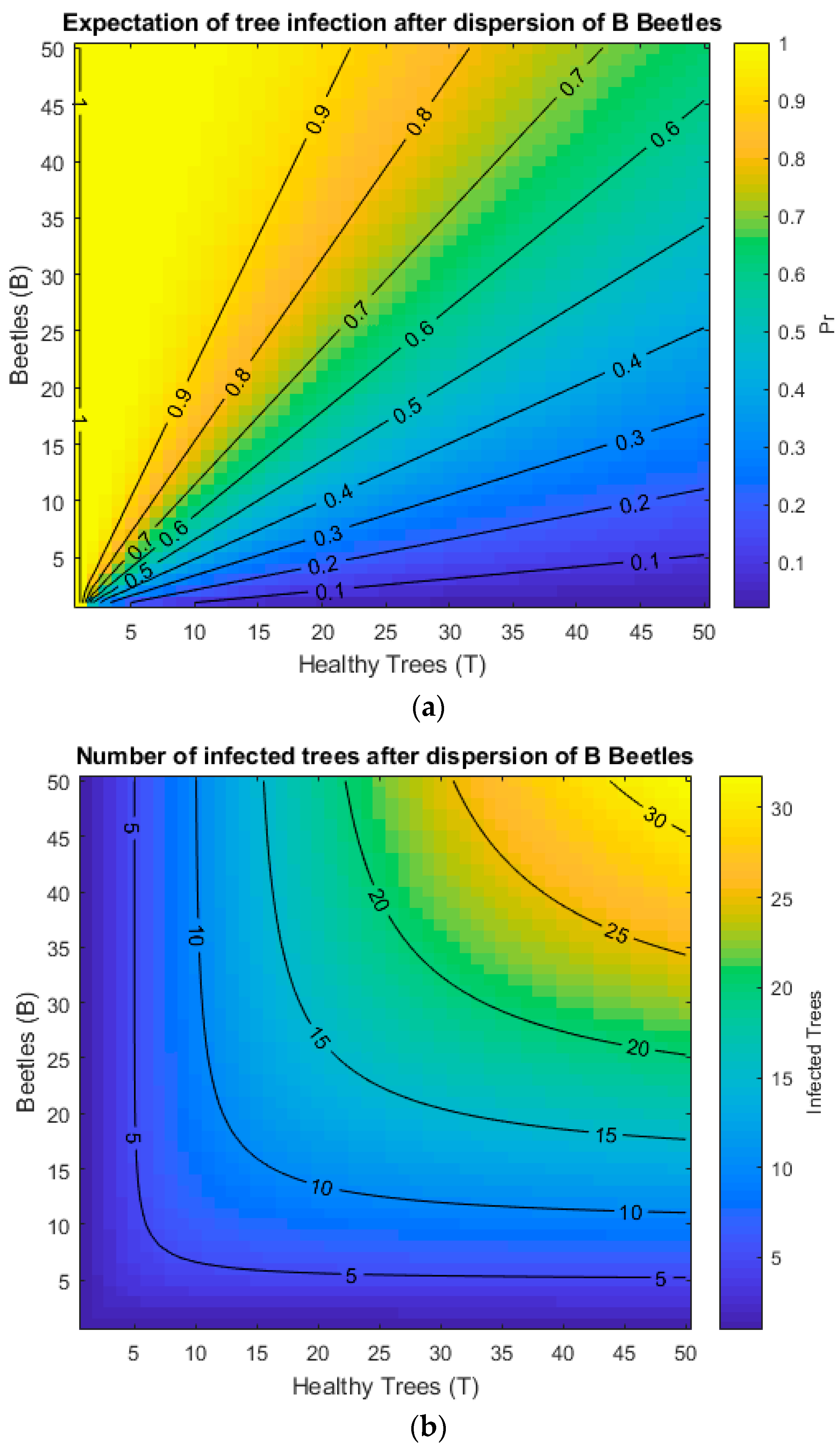
2.8. Dispersal
2.9. Study Area
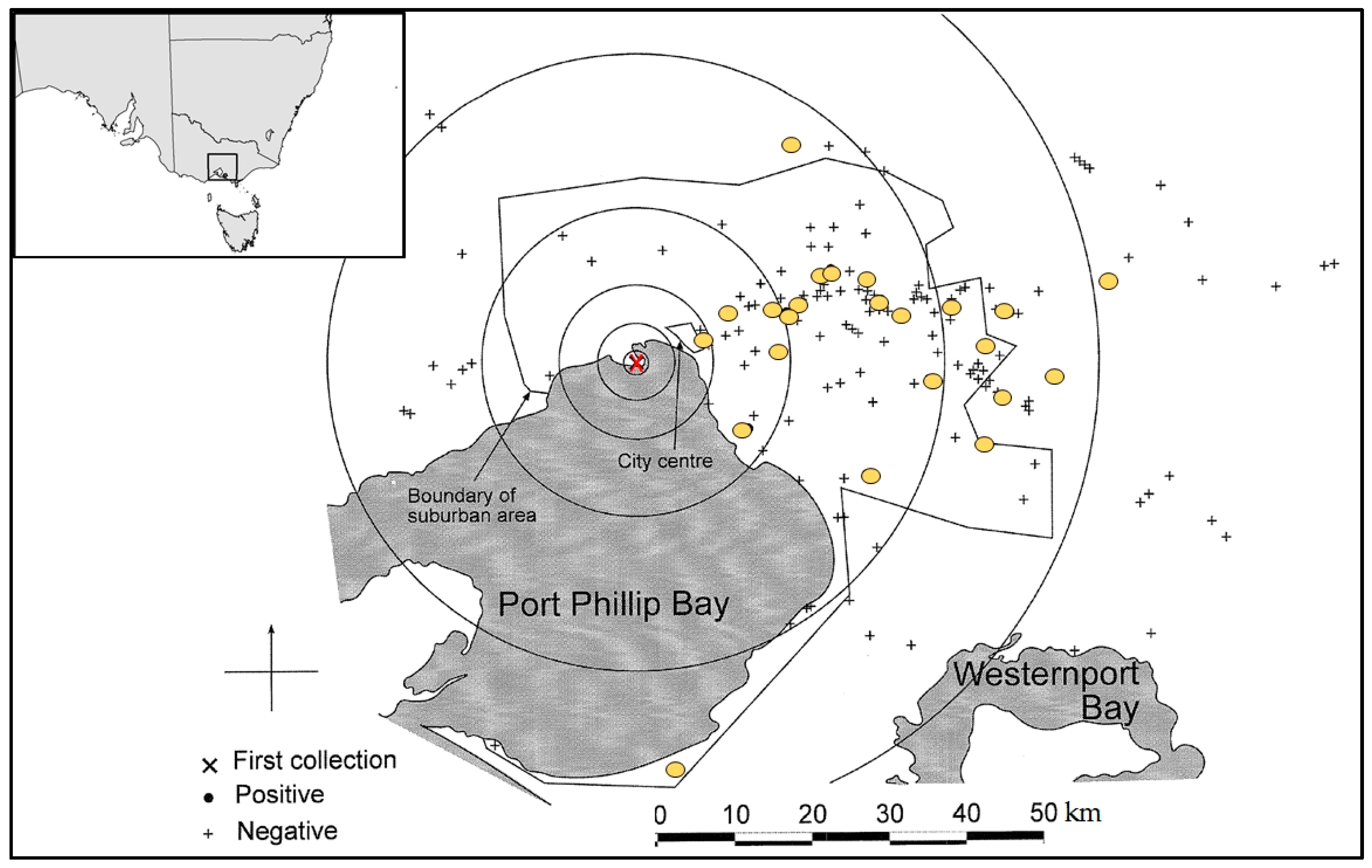
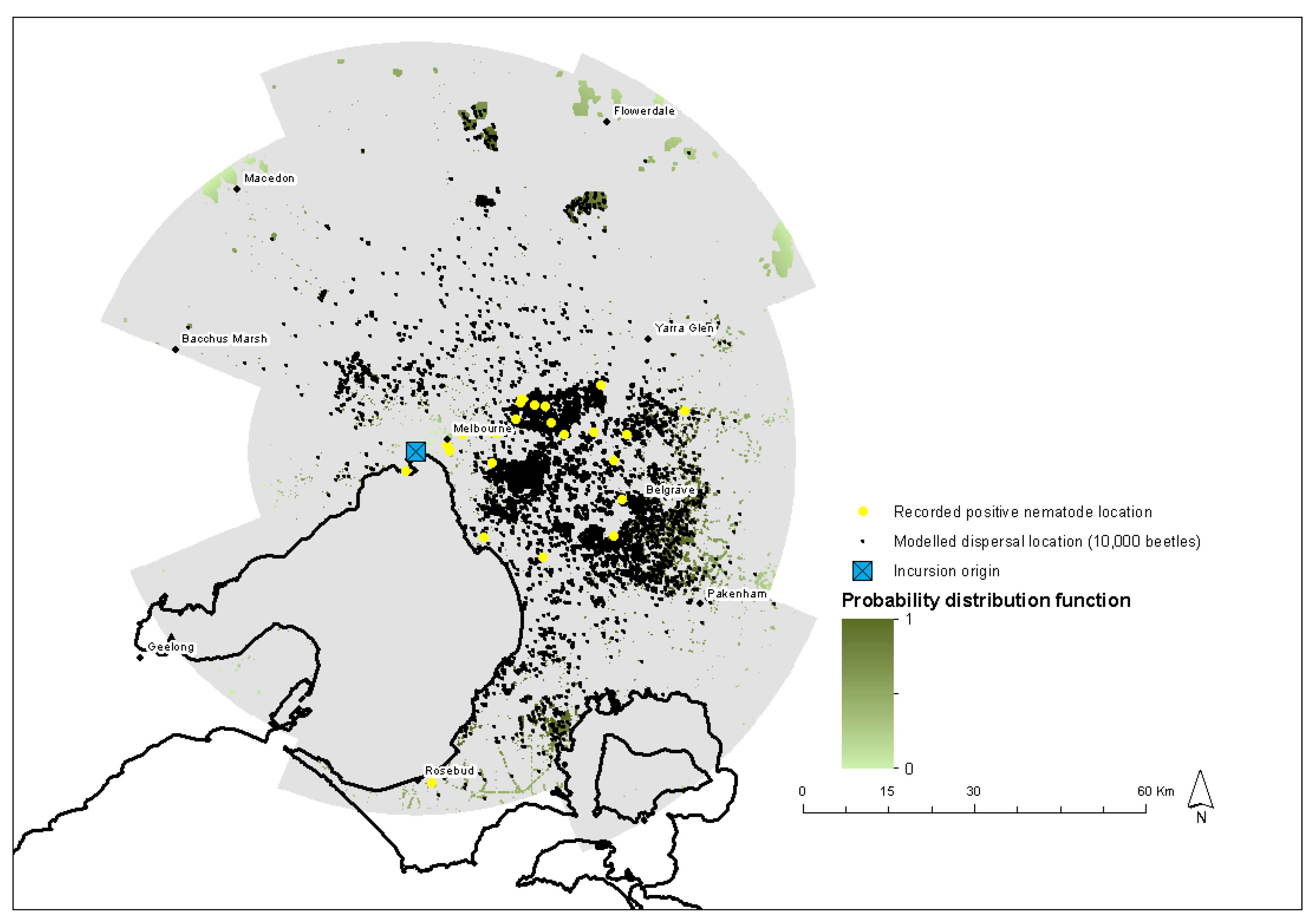
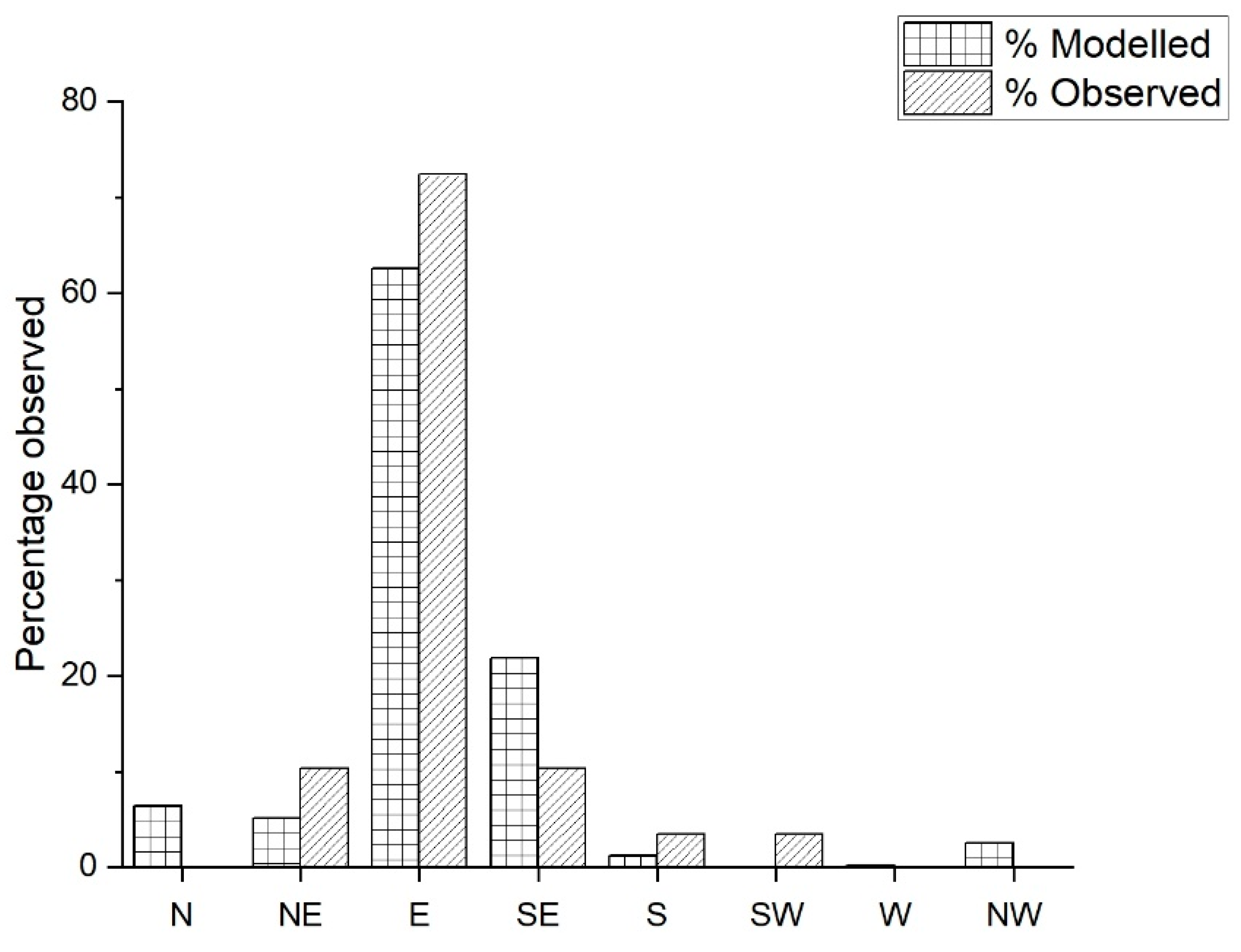
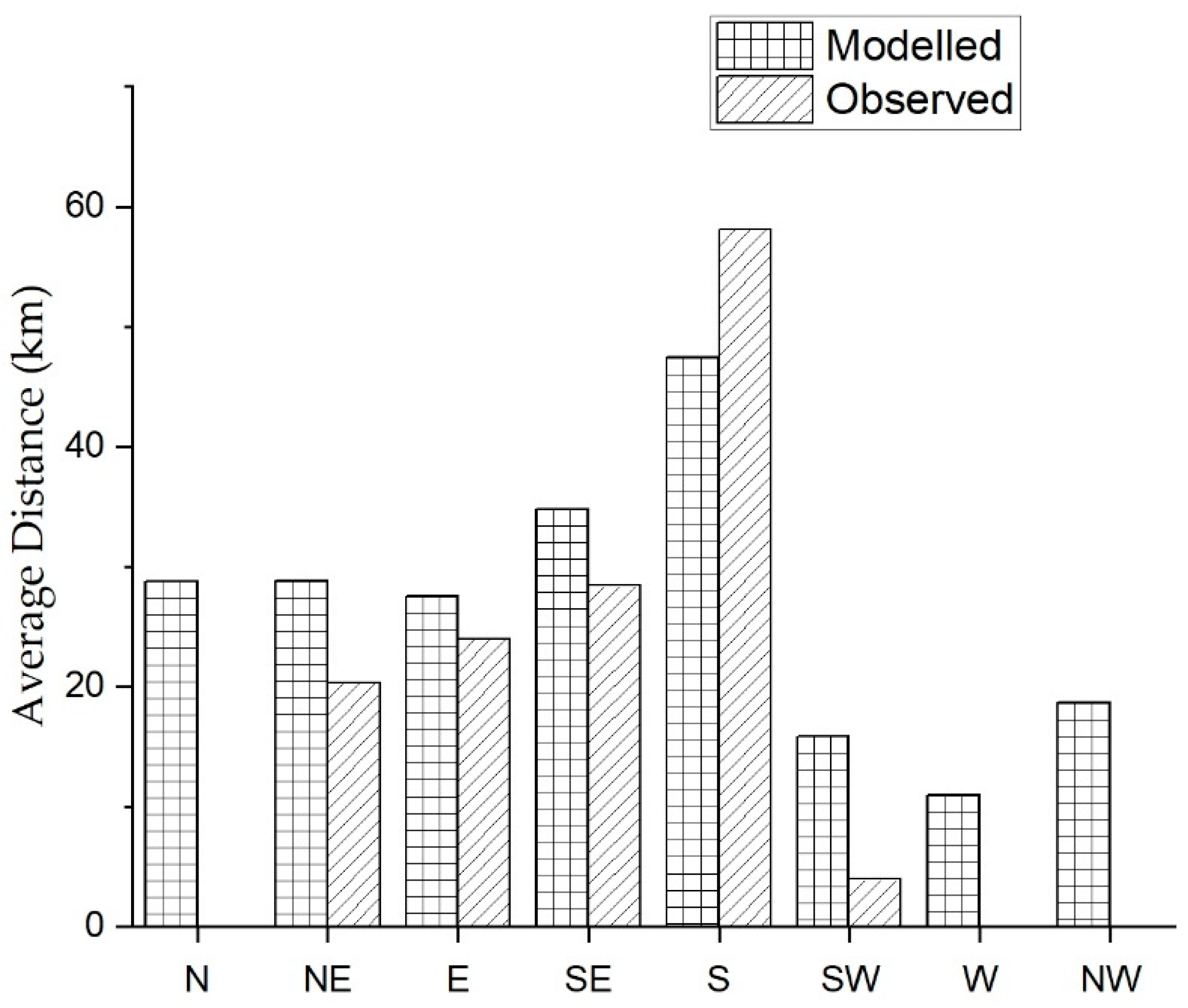
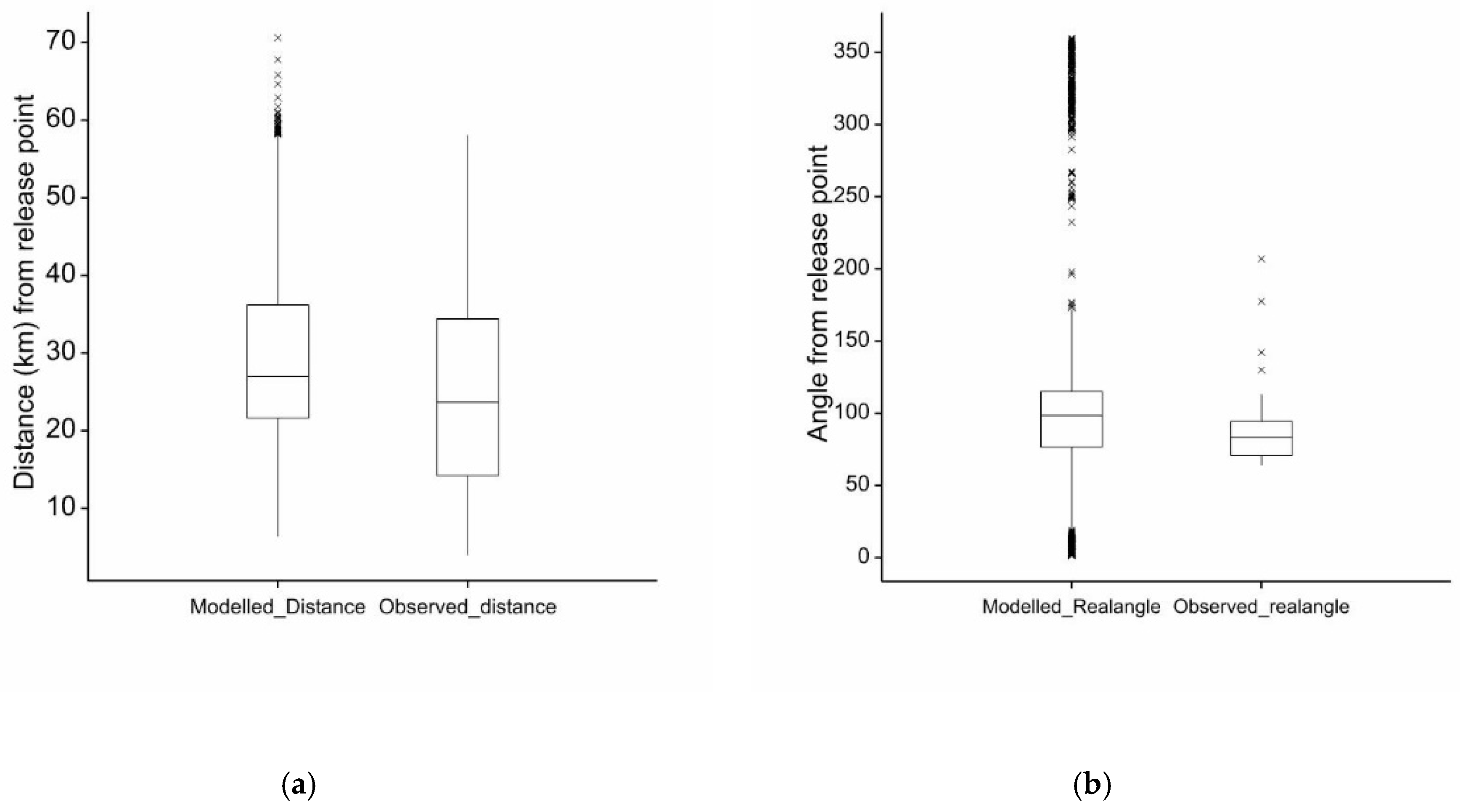
2.10. Verification of Model
Historical Incursion
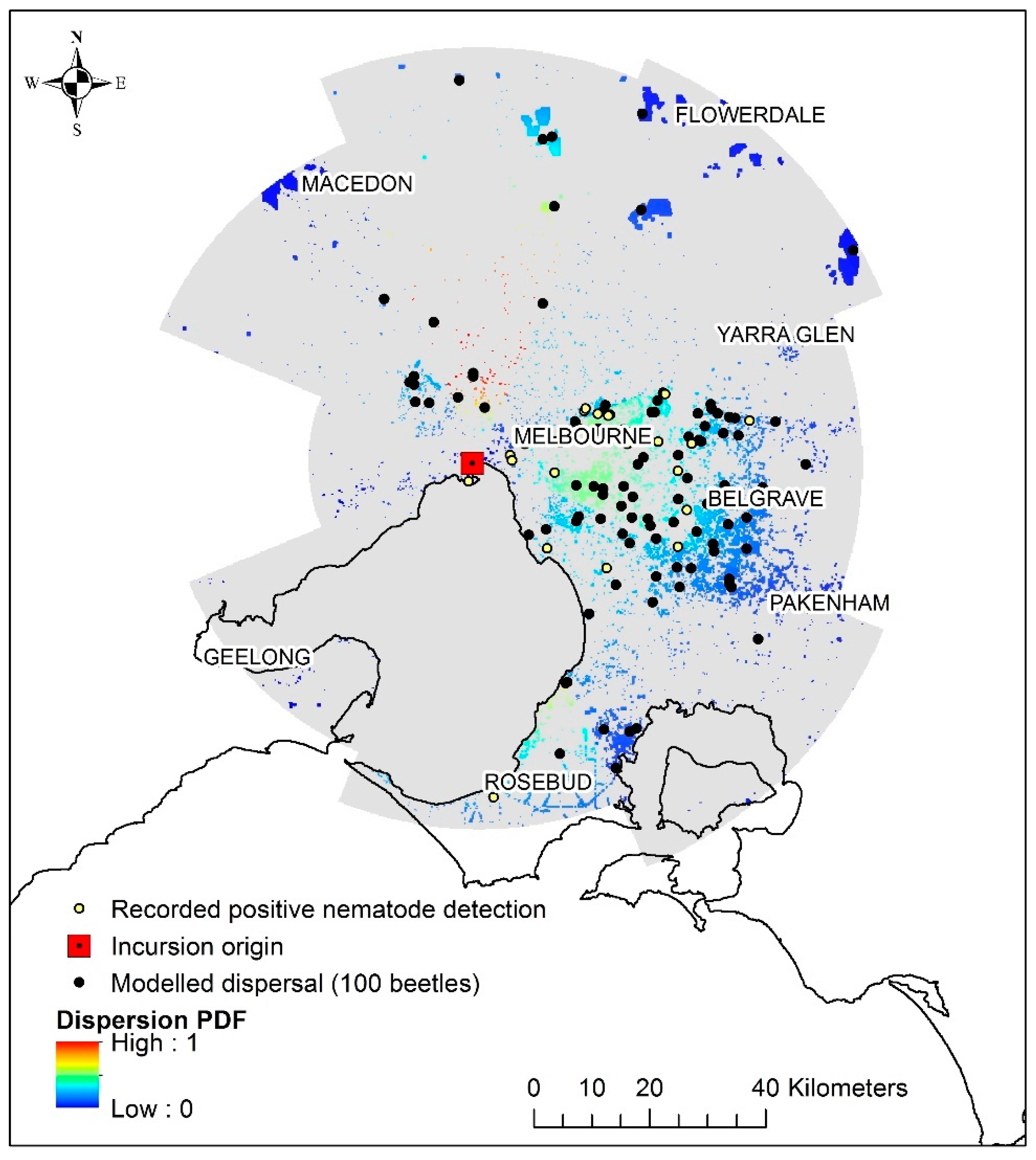
3. Results
- (1)
- 10,000 simulated beetles for comparative analysis against the observed data;
- (2)
- 100 simulated beetles as an indication to what was thought to happen in 1999/2000.
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DAWR. National Forest Biosecurity Surveillance Strategy; Plant Health Australia: Canberra, Australia, 2018; p. 32.
- Mohammed, C.; Walshe, T.; Wardlaw, T.; Stone, C.; Beadle, C.; Lawson, S. An Audit of Forest Biosecurity Arrangements and Preparedness in Australia; Forest & Wood Products Australia Ltd.: Melboune, Australia, 2011; p. 109. [Google Scholar]
- Bashford, R. The development of a port surrounds trapping system for the detection of exotic forest insect pests in Australia. In New Advances and Contributions to Forestry Research; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar]
- Carnegie, A. A decade of forest health surveillance in Australia: An overview. Aust. For. 2008, 71, 161–163. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Venn, T.; Lawson, S.; Nagel, M.; Wardlaw, T.; Cameron, N.; Last, I. An analysis of pest risk and potential economic impact of pine wilt disease to Pinus plantations in Australia. Aust. For. 2018, 81, 24–36. [Google Scholar] [CrossRef]
- Lawson, S.A.; Carnegie, A.J.; Cameron, N.; Wardlaw, T.; Venn, T.J. Risk of exotic pests to the Australian forest industry. Aust. For. 2018, 81, 3–13. [Google Scholar] [CrossRef]
- Fan, J.T. Invasive Species Compendium. Datasheet Report for Monochamus alternatus (Japanese pine sawyer). Available online: https://www.cabi.org/isc/datasheet/34719 (accessed on 23 February 2018).
- Australian Government Department of Agriculture and Water Resources. National Priority Plant Pests 2016. Available online: http://www.agriculture.gov.au/pests-diseases-weeds/plant/national-priority-plant-pests-2016 (accessed on 23 February 2018).
- Akbulut, S.; Linit, M.J. Flight performance of Monochamus carolinensis (Coleoptera: Cerambycidae) with respect to nematode phoresis and beetle characteristics. Environ. Entomol. 1999, 28, 1014–1020. [Google Scholar] [CrossRef]
- Togashi, K.; Shigesada, N. Spread of the pinewood nematode vectored by the Japanese pine sawyer: Modeling and analytical approaches. Popul. Ecol. 2006, 48, 271–283. [Google Scholar] [CrossRef]
- Mota, M.M.; Vieira, P. Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Springer, Dordrecht: Tokyo, Japan, 2008. [Google Scholar]
- Wang, Q.E. Cerambycidae of the World: Biology and Pest Management; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Akbulut, S.; Stamps, W. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Grimm, V.; Berger, U.; Bastiansen, F.; Eliassen, S.; Ginot, V.; Giske, J.; Goss-Custard, J.; Grand, T.; Heinz, S.K.; Huse, G. A standard protocol for describing individual-based and agent-based models. Ecol. Model. 2006, 198, 115–126. [Google Scholar] [CrossRef]
- Grimm, V.; Berger, U.; DeAngelis, D.L.; Polhill, J.G.; Giske, J.; Railsback, S.F. The ODD protocol: A review and first update. Ecol. Model. 2010, 221, 2760–2768. [Google Scholar] [CrossRef] [Green Version]
- Weiss, J.; Sheffield, K.; Weeks, A.; Smith, D. Modelling the Incursion and Spread of a Forestry Pest within Victoria; Technical report, June 2018; Agriculture Victoria Research, Department of Economic Development, Jobs, Transport and Resources: Melbourne, Australia, 2018; p. 49. [CrossRef]
- Jeffrey, S.J.; Carter, J.O.; Moodie, K.B.; Beswick, A.R. Using spatial interpolation to construct a comprehensive archive of Australian climate data. Environ. Model. Softw. 2001, 16, 309–330. [Google Scholar] [CrossRef]
- Stephens, P.A.; Sutherland, W.J.; Freckleton, R.P. What is the allee effect? Oikos 1999, 87, 185–190. [Google Scholar] [CrossRef]
- Coop, L. Monochamus alternatus (Japanese Pine Sawyer Beetle) Degree-Day Model Documentation. Available online: http://uspest.org/wea/Monochamus_alternatus_model.pdf (accessed on 23 February 2018).
- Bodart, J. Flight activity, oviposition pit distribution, and emergence densities of Monochamus titillator and M. carolinensis in the Ozark-St. Francis national forest in Arkansas; University of Arkansas: Fayetteville, AR, USA, 2017. [Google Scholar]
- Hodda, M.; Smith, D.; Smith, I.; Nambiar, L.; Pascoe, I. Incursion management in the face of multiple uncertainties: A case study of an unidentified nematode associated with dying pines near Melbourne, Australia. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 15–40. [Google Scholar]
- Evans, H.F.; McNamara, D.G.; Braash, H.; Chadoeuf, J.; Magnusson, C. Pest risk analysis (PRA) for the territories of the European Union (as PRA area) on Bursaphelenchus xylophilus and its vectors in the genus Monochamus. EPPO Bull. 1996, 26, 199–249. [Google Scholar] [CrossRef]
- Allison, J.D.; Borden, J.H.; Seybold, S.J. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- Cameron, N.L.; Carnegie, A.J.; Wardlaw, T.; Lawson, S.; Venn, T. Economic appraisal of sirex wood wasp (Sirex noctilio) control in Australian pine plantations. Aust. For. 2018, 81, 37–45. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | Property | Description |
| X | X-coordinate (meters) of agent | |
| Y | Y-coordinate (meters) of agent | |
| PhenoStage | Current phenological stage of agent [1,2,3,4,5] | |
| DegDayCount | Current cumulative degree count for agent | |
| AdultDayCount | Current number of days since emergence | |
| Male | Male/Female (true/false) | |
| Mated | Whether agent has mated (true/false) | |
| Infected | Is agent infected with nematode (true/false) |
| Days Required to Complete Each Phenological Stage at DD 12.5 | Victoria | Melbourne | |||
|---|---|---|---|---|---|
| Completion of Stage (median) | Stage Length (days) | Completion of Stage (median) | Stage Length (days) | ||
| Eggs | 80 | 12 September | 73 | 25 September | 86 |
| Larvae | 705 | 2 January | 112 | 18 January | 115 |
| Pupae | 865 | 19 January | 17 | 8 February | 21 |
| Pre-oviposition | 965 | 30 January | 11 | 21 February | 13 |
| Oviposition | 1333 | 20 March | 49 | 7 May | 75 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weiss, J.; Sheffield, K.; Weeks, A.; Smith, D. Modelling the Incursion and Spread of a Forestry Pest: Case Study of Monochamus alternatus Hope (Coleoptera: Cerambycidae) in Victoria. Forests 2019, 10, 198. https://0-doi-org.brum.beds.ac.uk/10.3390/f10020198
Weiss J, Sheffield K, Weeks A, Smith D. Modelling the Incursion and Spread of a Forestry Pest: Case Study of Monochamus alternatus Hope (Coleoptera: Cerambycidae) in Victoria. Forests. 2019; 10(2):198. https://0-doi-org.brum.beds.ac.uk/10.3390/f10020198
Chicago/Turabian StyleWeiss, John, Kathryn Sheffield, Anna Weeks, and David Smith. 2019. "Modelling the Incursion and Spread of a Forestry Pest: Case Study of Monochamus alternatus Hope (Coleoptera: Cerambycidae) in Victoria" Forests 10, no. 2: 198. https://0-doi-org.brum.beds.ac.uk/10.3390/f10020198