Inorganic Nitrogen Addition Affects Soil Respiration and Belowground Organic Carbon Fraction for a Pinus tabuliformis Forest

Abstract
:1. Introduction
2. Materials and Methods
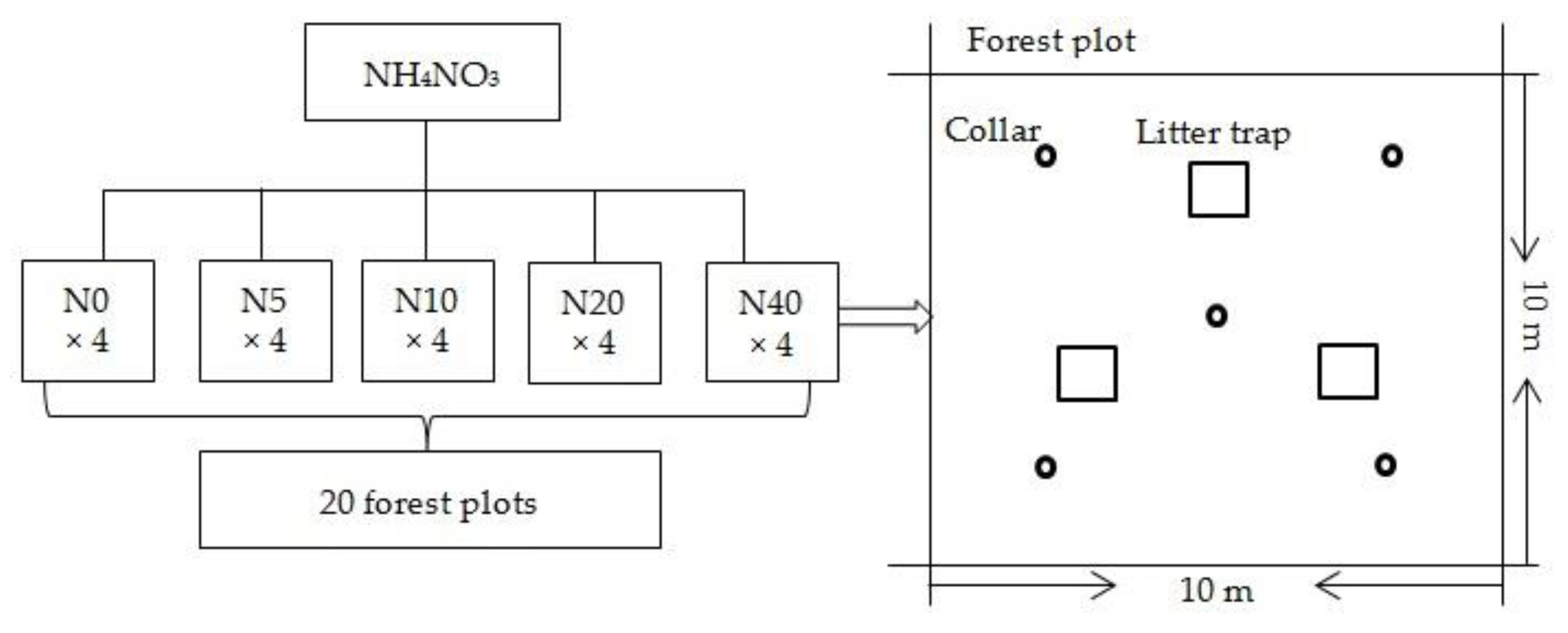
2.1. Site Description and Experimental Design
2.2. Soil Respiration Measurements
2.3. Fine Root, Litter, and Soil Sampling
2.4. Chemical Analysis
2.5. Statistical Analysis
3. Results
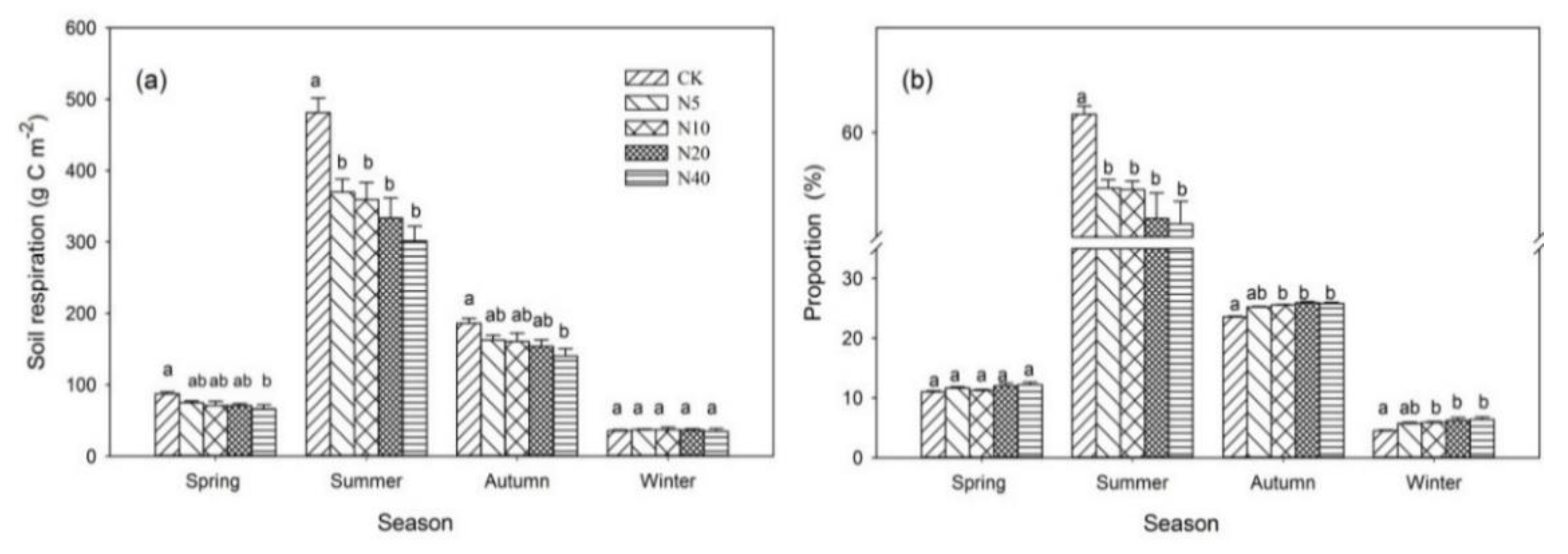
3.1. Soil Respiration Rate over Time
3.2. The Effect of Inorganic Nitrogen Addition on Soil Physicochemical Properties
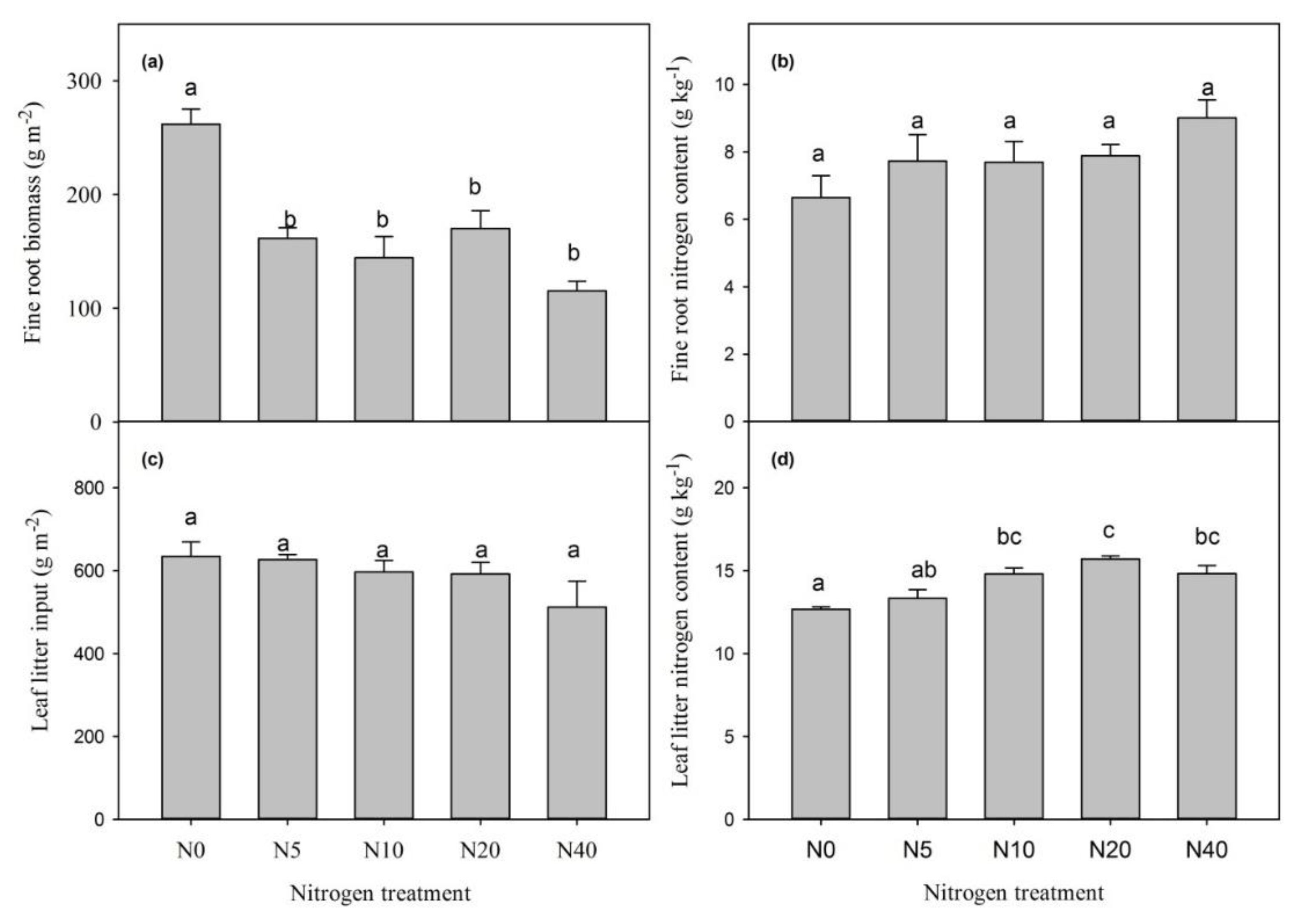
3.3. The Effect of Inorganic Nitrogen Addition on Litter Production and Its Nitrogen Content
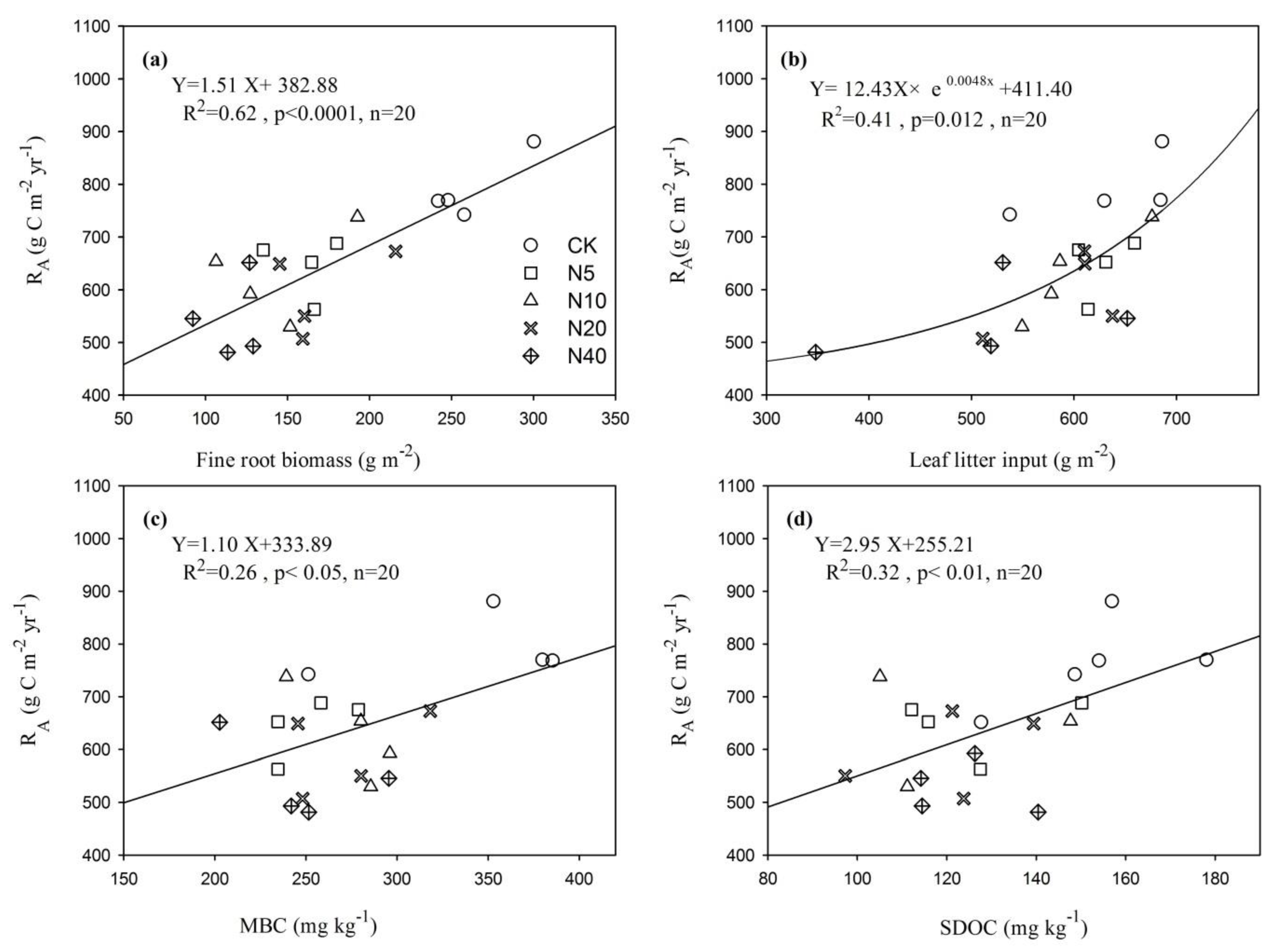
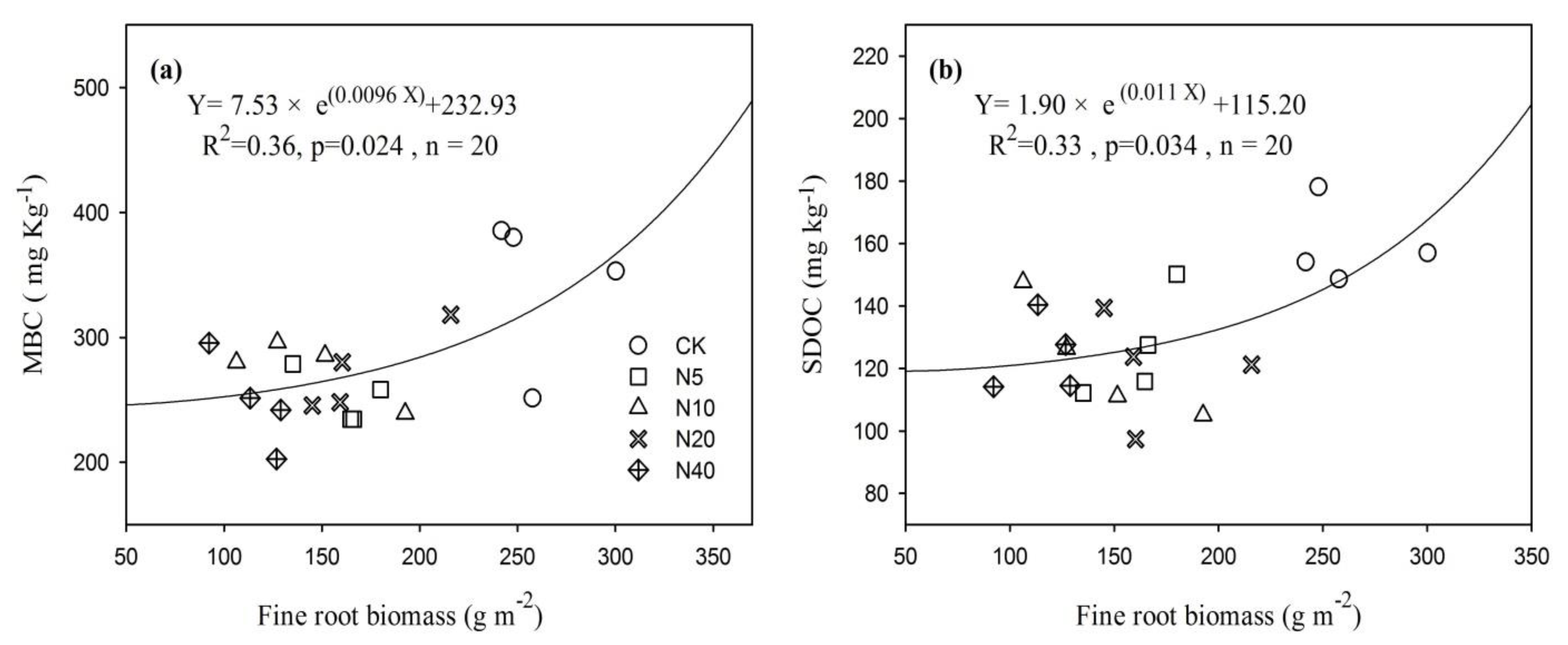
3.4. The Correlation of RA with the SOC Fraction and Litter Input
3.5. Variation of Soil Organic Carbon by Litter Input
4. Discussion
4.1. The Effect of Nitrogen Addition on Soil Respiration Rate at Temporal Scales
4.2. The Effect of Nitrogen Addition on the Soil Carbon Fraction and Plant Litter Input
4.3. The Correlation of RA with SOC Fraction and Plant Litter Input
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nature 2008, 451, 293–296. [Google Scholar] [CrossRef] [Green Version]
- Greaver, T.L.; Clark, C.M.; Compton, J.E.; Vallano, D.; Talhelm, A.F.; Weaver, C.P.; Band, L.E.; Baron, J.S.; Davidson, E.A.; Tague, C.L.; et al. Key ecological responses to nitrogen are altered by climate change. Nat. Clim. Chang. 2016, 6, 836–843. [Google Scholar] [CrossRef]
- Zhou, L.Y.; Zhou, X.H.; Zhang, B.C.; Lu, M.; Luo, Y.; Liu, L.L.; Li, B. Different responses of soil respiration and itscomponents to nitrogen addition among biomes: A meta-analysis. Glob. Chang. Biol. 2014, 20, 2332–2343. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Han, W.; Tang, A.; Shen, J.; Cui, Z.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef]
- Liu, L.; Greaver, T.L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol. Lett. 2010, 13, 819–828. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.-A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Chen, Z.M.; Xu, Y.H.; Fan, J.L.; Yu, H.Y.; Ding, W.X. Soil autotrophic and heterotrophic respiration in response to different N fertilization and environmental conditions from a cropland in Northeast China. Soil Biol. Biochem. 2017, 110, 103–115. [Google Scholar] [CrossRef]
- Xu, M.; Shang, H. Contribution of soil respiration to the global carbon equation. J. Plant Physiol. 2016, 203, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Bond-Lamberty, B.; Thomson, A. A global database of soil respiration data. Biogeosciences 2010, 7, 1321–1344. [Google Scholar] [CrossRef]
- Sulzman, E.W.; Brant, J.B.; Bowden, R.D.; Lajtha, K. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2 efflux in an old growth coniferous forest. Biogeochemistry 2005, 73, 231–256. [Google Scholar] [CrossRef]
- Wang, C.; Yang, J. Rhizospheric and heterotrophic components of soil respiration in six Chinese temperate forests. Glob. Chang. Boil. 2007, 13, 123–131. [Google Scholar] [CrossRef]
- Di Lonardo, D.; De Boer, W.; Gunnewiek, P.K.; Hannula, S.; Van Der Wal, A. Priming of soil organic matter: Chemical structure of added compounds is more important than the energy content. Soil Boil. Biochem. 2017, 108, 41–54. [Google Scholar] [CrossRef]
- de Graaff, M.A.; Classen, A.T.; Castro, H.F.; Schadt, C.W. Labile soil carbon inputs mediate the soil microbial community composition and plant residue decomposition rates. New Phytol. 2010, 188, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Smith, P.; Moncrieff, J.B.; Smith, J.U. Similar response of labile and resistant soil organic matter pools to changes in temperature. Nature 2005, 433, 57–59. [Google Scholar] [CrossRef]
- von Lützow, M.; Kögel-Knabner, I.; Ekschmitt, K.; Flessa, H.; Guggenberger, G.; Matzner, E.; Marschner, B. SOM fractionation methods: Relevance to functional pools and to stabilization mechanisms. Soil Biol. Biochem 2007, 39, 2183–2207. [Google Scholar] [CrossRef]
- Zhang, H.H.; Zhou, Z.Y. Recalcitrant carbon controls the magnitude of soil organic matter mineralization in temperate forests of northern China. For. Ecosyst. 2018, 5, 17. [Google Scholar] [CrossRef]
- Bae, K.; Fahey, T.J.; Yanai, R.D.; Fisk, M. Soil Nitrogen Availability Affects Belowground Carbon Allocation and Soil Respiration in Northern Hardwood Forests of New Hampshire. Ecosystems 2015, 18, 1179–1191. [Google Scholar] [CrossRef]
- Huo, C.; Luo, Y.; Cheng, W. Rhizosphere priming effect: A meta-analysis. Soil Boil. Biochem. 2017, 111, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Sokol, N.W.; Kuebbing, S.E.; Karlsen-Ayala, E.; Bradford, M.A. Evidence for the primacy of living root inputs, not root or shoot litter, in forming soil organic carbon. New Phytol. 2019, 221, 233–246. [Google Scholar] [CrossRef]
- Frey, S.D.; Ollinger, S.; Nadelhoffer, K.; Bowden, R.; Brzostek, E.; Burton, A.; Caldwell, B.A.; Crow, S.; Goodale, C.L.; Grandy, A.S.; et al. Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests. Biogeochemistry 2014, 121, 305–316. [Google Scholar] [CrossRef]
- Kamble, P.N.; Rousk, J.; Frey, S.D.; Bååth, E. Bacterial growth and growth-limiting nutrients following chronic nitrogen additions to a hardwood forest soil. Soil Boil. Biochem. 2013, 59, 32–37. [Google Scholar] [CrossRef]
- Treseder, K.K. Nitrogen additions and microbial biomass: A meta-analysis of ecosystem studies. Ecol. Lett. 2008, 11, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Chen, H.Y.H.; Ruan, H. Global negative effects of nitrogen deposition on soil microbes. ISME J. 2018, 12, 1817–1825. [Google Scholar] [CrossRef]
- Li, W.B.; Jin, C.J.; Guan, D.X.; Wang, Q.K.; Wang, A.Z.; Yuan, F.H.; Wu, J.B. The effects of simulated nitrogen deposition on plant root traits: A meta-analysis. Soil Biol. Biochem. 2015, 82, 112–118. [Google Scholar] [CrossRef]
- Guo, H.; Wang, B.; Ma, X.Q.; Zhao, G.D.; Xu, S.N. An evaluation on ecological service function of pinus tabuliformis forest in China. Sci. China Life Sci. 2008, 38, 565–572. (In Chinese) [Google Scholar]
- He, C.E.; Liu, X.; Fangmeier, A.; Zhang, F. Quantifying the total airborne nitrogen input into agroecosystems in the North China Plain. Agric. Ecosyst. Environ. 2007, 121, 395–400. [Google Scholar] [CrossRef]
- He, C.-E.; Wang, X.; Liu, X.; Fangmeier, A.; Christie, P.; Zhang, F. Nitrogen deposition and its contribution to nutrient inputs to intensively managed agricultural ecosystems. Ecol. Appl. 2010, 20, 80–90. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zheng, C.; Ren, D.; Chou, C.-L.; Liu, J.; Zeng, R.; Wang, Z.; Zhao, F.; Ge, Y. Distribution of potentially hazardous trace elements in coals from Shanxi province, China. Fuel 2004, 83, 129–135. [Google Scholar] [CrossRef]
- van’t Hoff, J.H. Lectures on Theoretical and Physical Chemistry; Lehfeldt, R.A., Ed.; Edwart Arnold: London, UK, 1898; pp. 224–229. (In German) [Google Scholar]
- Zhou, Z.; Xu, M.; Kang, F.; Sun, O.J. Maximum temperature accounts for annual soil CO2 efflux in temperate forests of Northern China. Sci. Rep. 2015, 5, 12142. [Google Scholar] [CrossRef]
- Mo, J.; Zhang, W.; Zhu, W.; Gundersen, P.; Fang, Y.; Li, D.; Wang, H. Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Glob. Chang. Biol. 2008, 14, 403–412. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy and Soil Science Society of American: Madison, WI, USA, 1982; pp. 101–129. [Google Scholar]
- Gallaher, R.N.; Weldon, C.O.; Boswell, F.C. A Semiautomated Procedure for Total Nitrogen in Plant and Soil Samples1. Soil Sci. Soc. J. 1976, 40, 887. [Google Scholar] [CrossRef]
- Tirol-Padre, A.; Ladha, J.K. Assessing the Reliability of Permanganate-Oxidizable Carbon as an Index of Soil Labile Carbon. Soil Sci. Soc. J. 2004, 68, 969. [Google Scholar] [CrossRef]
- Chen, X.; Liu, J.; Deng, Q.; Yan, J.; Zhang, D. Effects of elevated CO2 and nitrogen addition on soil organic carbon fractions in a subtropical forest. Plant Soil 2012, 357, 25–34. [Google Scholar] [CrossRef]
- Vance, E.; Brookes, P.; Jenkinson, D. An extraction method for measuring soil microbial biomass C. Soil Boil. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Bu, X.; Wang, L.; Ma, W.; Yu, X.; McDowell, W.H.; Ruan, H. Spectroscopic characterization of hot-water extractable organic matter from soils under four different vegetation types along an elevation gradient in the Wuyi Mountains. Geoderma 2010, 159, 139–146. [Google Scholar] [CrossRef]
- Balaria, A.; Johnson, C.E.; Groffman, P.M.; Fisk, M.C. Effects of calcium silicate treatment on the composition of forest floor organic matter in a northern hardwood forest stand. Biogeochemistry 2014, 122, 313–326. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 23 April 2018).
- Fan, H.; Wu, J.; Liu, W.; Yuan, Y.; Huang, R.; Liao, Y.; Li, Y. Nitrogen deposition promotes ecosystem carbon accumulation by reducing soil carbon emission in a subtropical forest. Plant Soil 2014, 379, 361–371. [Google Scholar] [CrossRef]
- Maaroufi, N.; Nordin, A.; Hasselquist, N.J.; Bach, L.H.; Palmqvist, K.; Gundale, M.J. Anthropogenic nitrogen deposition enhances carbon sequestration in boreal soils. Glob. Chang. Boil. 2015, 21, 3169–3180. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, L.; Zhang, C.; Bu, W.; Zhao, X.; Gadow, K.V. Combined effects of nitrogen addition and organic matter manipulation on soil respiration in a Chinese pine forest. Environ. Sci. Pollut. 2016, 23, 22701–22710. [Google Scholar] [CrossRef]
- Du, Y.; Guo, P.; Liu, J.; Wang, C.; Yang, N.; Jiao, Z. Different types of nitrogen deposition show variable effects on the soil carbon cycle process of temperate forests. Glob. Chang. Boil. 2014, 20, 3222–3228. [Google Scholar] [CrossRef]
- Fang, C.; Ye, J.-S.; Gong, Y.; Pei, J.; Yuan, Z.; Xie, C.; Zhu, Y.; Yu, Y. Seasonal responses of soil respiration to warming and nitrogen addition in a semi-arid alfalfa-pasture of the Loess Plateau, China. Sci. Total. Environ. 2017, 590, 729–738. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Z.; Lin, C.; Giardina, C.P.; Xiong, D.; Lin, W.; Chen, S.; Xu, C.; Chen, G.; Xie, J.; et al. Will nitrogen deposition mitigate warming-increased soil respiration in a young subtropical plantation? Agric. For. Meteorol. 2017, 246, 78–85. [Google Scholar] [CrossRef]
- Zhou, X.; Zhang, Y. Seasonal pattern of soil respiration and gradual changing effects of nitrogen addition in a soil of the Gurbantunggut Desert, northwestern China. Atmos. Environ. 2014, 85, 187–194. [Google Scholar] [CrossRef]
- Tao, B.; Song, C.; Guo, Y. Short-term Effects of Nitrogen Additions and Increased Temperature on Wetland Soil Respiration, Sanjiang Plain, China. Wetlands 2013, 33, 727–736. [Google Scholar] [CrossRef]
- Sampson, D.A.; Janssens, I.A.; Yuste, J.C.; Ceulemans, R. Basal rates of soil respiration are correlated with photosynthesis in a mixed temperate forest. Glob. Chang. Boil. 2007, 13, 2008–2017. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Z.; Zha, T.; Luo, Z.; Zheng, J.; Sun, O.J. Predicting soil respiration using carbon stock in roots, litter and soil organic matter in forests of Loess Plateau in China. Soil Boil. Biochem. 2013, 57, 135–143. [Google Scholar] [CrossRef]
- Tu, L.H.; Hu, T.X.; Zhang, J.A.; Li, R.H.; Dai, H.Z.; Luo, S.H. Short-term simulated nitrogen deposition increases carbon sequestration in a Pleioblastus amarus plantation. Plant Soil 2011, 340, 383–396. [Google Scholar] [CrossRef]
- Chen, H.; Tian, H.-Q. Does a General Temperature-Dependent Q10 Model of Soil Respiration Exist at Biome and Global Scale? J. Integr. Plant Boil. 2005, 47, 1288–1302. [Google Scholar] [CrossRef]
- McDowell, W.H.; Magill, A.H.; A Aitkenhead-Peterson, J.; Aber, J.D.; Merriam, J.L.; Kaushal, S.S. Effects of chronic nitrogen amendment on dissolved organic matter and inorganic nitrogen in soil solution. Ecol. Manag. 2004, 196, 29–41. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Z.; Hu, Y.; Wang, Q.; Wang, S.; Zhang, Z.; Zhao, Y. Climate change affects soil labile organic carbon fractions in a Tibetan alpine meadow. J. Soils Sediments 2016, 17, 326–339. [Google Scholar]
- Bengtsson, G.; BergWall, C. Fate of 15N labelled nitrate and ammonium in a fertilized forest soil. Soil Boil. Biochem. 2000, 32, 545–557. [Google Scholar] [CrossRef]
- Bowman, W.D.; Cleveland, C.C.; Halada, L.; Hreško, J.; Baron, J.S. Negative impact of nitrogen deposition on soil buffering capacity. Nat. Geosci. 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Boil. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Li, Y.; Sun, J.; Tian, D.; Wang, J.; Ha, D.; Qu, Y.; Jing, G.; Niu, S. Soil acid cations induced reduction in soil respiration under nitrogen enrichment and soil acidification. Sci. Total. Environ. 2018, 615, 1535–1546. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, Y.; He, T. Aboveground and belowground litter have equal contributions to soil CO2 emission: An evidence from a 4-year measurement in a subtropical forest. Plant Soil 2017, 421, 7–17. [Google Scholar] [CrossRef]
- Cusack, D.F.; Silver, W.L.; Torn, M.S.; McDowell, W.H. Effects of nitrogen additions on above-and below-ground carbon dynamics in two tropical forests. Biogeochemistry 2011, 104, 203–225. [Google Scholar] [CrossRef]
- Phillips, R.P.; Fahey, T.J. Fertilization effects on fineroot biomass, rhizosphere microbes and respiratory fluxes in hardwood forest soils. New Phytol. 2007, 176, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Wang, W. Combination of nitrogen and phosphorus fertilization enhance ecosystem carbon sequestration in a nitrogen-limited temperate plantation of Northern China. Ecol. Manag. 2015, 341, 59–66. [Google Scholar] [CrossRef]
- Lebauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef]
- Rothstein, D.E.; Cregg, B.M. Effects of nitrogen form on nutrient uptake and physiology of Fraser fir (Abies fraseri). Ecol. Manag. 2005, 219, 69–80. [Google Scholar] [CrossRef]
- Smirnoff, N.; Stewart, G.R. Nitrate assimilation and translocation by higher plants:Comparative physiology and ecological consequences. Physiol. Plant. 1985, 64, 133–140. [Google Scholar] [CrossRef]
- Brzostek, E.; Phillips, R.P.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon–nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar]
- Midgley, M.G.; Phillips, R.P. Resource stoichiometry and the biogeochemical consequences of nitrogen deposition in a mixed deciduous forest. Ecology 2016, 97, 3369–3378. [Google Scholar] [CrossRef]
- Lu, M.; Yang, Y.H.; Luo, Y.; Fang, C.M.; Zhou, X.H.; Chen, J.K.; Yang, X.; Li, B. Responses of ecosystem nitrogen cycle to nitrogen addition: A meta-analysis. New Phytol. 2011, 189, 1040–1050. [Google Scholar] [CrossRef]
- Nadelhoffer, K.J. The potential effects of nitrogen deposition on fine-root production in forest ecosystems. New Phytol. 2000, 147, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Knorr, M.; Frey, S.D.; Curtis, P.S. Nitrogen additions and litter decomposition: A meta-analysis. Ecology 2008, 86, 3252–3257. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.; Garten, C.; Andrews, J. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Eberwein, J.; Oikawa, P.; Allsman, L.; Jenerette, G.; Allsman, L. Carbon availability regulates soil respiration response to nitrogen and temperature. Soil Boil. Biochem. 2015, 88, 158–164. [Google Scholar] [CrossRef]
- Burton, A.J.; Jarvey, J.C.; Jarvi, M.P.; Zak, D.R.; Pregitzer, K.S. Chronic N deposition alters root respiration-tissue N relationship in northern hardwood forests. Glob. Chang. Biol. 2012, 18, 258–266. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Vicca, S.; Janssens, I.A.; Sardans, J.; Luyssaert, S.; Campioli, M.; Chapin, F.S.; Ciais, P.; Malhi, Y.; Obersteiner, M.; et al. Addendum: Nutrient availability as the key regulator of global forest carbon balance. Nat. Clim. Chang. 2014, 4, 643. [Google Scholar] [CrossRef]
- Yan, D.; Wang, D.; Yang, L. Long-term effect of chemical fertilizer, straw, and manure on labile organic matter fractions in a paddy soil. Boil. Fertil. Soils 2007, 44, 93–101. [Google Scholar] [CrossRef]
- Guigue, J.; Mathieu, O.; Levêque, J.; Mounier, S.J.L.; Laffont, R.; Maron, P.A.; Navarro, N.; Chateau, C.; Amiotte-Suchet, P.; Lucas, Y.; et al. A comparison of extraction procedures for water-extractable organic matter in soils. Eur. J. Soil Sci. 2014, 65, 520–530. [Google Scholar] [CrossRef]
- Gunina, A.; Dippold, M.A.; Glaser, B.; Kuzyakov, Y. Fate of low molecular weight organic substances in an arable soil: From microbial uptake to utilisation and stabilisation. Soil Boil. Biochem. 2014, 77, 304–313. [Google Scholar] [CrossRef]
- Xia, M.; Talhelm, A.F.; Pregitzer, K.S. Chronic nitrogen deposition influences the chemical dynamics of leaf litter and fine roots during decomposition. Soil Boil. Biochem. 2017, 112, 24–34. [Google Scholar] [CrossRef]
- De Vries, F.T.; Caruso, T. Eating from the same plate? Revisiting the role of labile carbon inputs in the soil food web. Soil Boil. Biochem. 2016, 102, 4–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Högberg, M.N.; Bååth, E.; Nordgren, A.; Arnebrant, K.; Högberg, P. Contrasting effects of nitrogen availability on plant carbon supply to mycorrhizal fungi and saprotrophs—A hypothesis based on field observations in boreal forest. New Phytol. 2003, 160, 225–238. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | SOC (g kg−1) | TN (g kg−1) | SBD (g cm−3) | pH | PD (stem ha−1) | DBH (cm) |
|---|---|---|---|---|---|---|
| N0 | 22.53 ± 2.30 a | 1.47 ± 0.20 a | 1.23 ± 0.03 a | 6.01 ± 0.23 a | 1650 ± 119 a | 19.39 ± 0.52 a |
| N5 | 21.19 ± 3.51 a | 1.47 ± 0.16 a | 1.32 ± 0.03 a | 6.39 ± 0.25 a | 2250 ± 348 a | 17.41 ± 0.75 a |
| N10 | 20.10 ± 2.87 a | 1.38 ± 0.12 a | 1.22 ± 0.03 a | 6.14 ± 0.05 a | 1950 ± 284 a | 18.22 ± 1.48 a |
| N20 | 19.68 ± 1.19 a | 1.40 ± 0.10 a | 1.23 ± 0.02 a | 6.38 ± 0.30 a | 2000 ± 245 a | 18.96 ± 1.41 a |
| N40 | 20.75 ± 1.17 a | 1.45 ± 0.24 a | 1.32 ± 0.02 a | 6.93 ± 0.42 a | 2000 ± 108 a | 18.10 ± 0.92 a |
| RA | Annual cumulative soil respiration rate | ROC | Soil readily oxidizable organic carbon |
| RS | Seasonal cumulative soil respiration rate | HEOC | Hot-water-extractable organic carbon |
| Ri | The instantaneous soil respiration rate | MBC | Soil microbial biomass carbon |
| Rai | Annual average instantaneous soil respiration rate | SDOC | Soil dissolved organic carbon |
| R0 | Soil respiration rate at 0 °C | TN | Soil total nitrogen |
| R10 | Basal soil respiration rate at 10 °C | MBN | Soil microbial biomass nitrogen |
| Q10 | Temperature sensitivity of soil respiration | SDON | Soil dissolved nitrogen |
| SOC | Soil organic carbon | HEON | Hot-water-extractable nitrogen |
| Treatment | RA (g C m−2 year−1) | R0 (μmol CO2 m−2 s−1) | R10 (μmol CO2 m−2 s−1) | Rai (μmol CO2 m−2 s−1) | Q10 | T °C |
|---|---|---|---|---|---|---|
| CK | 790.25 ± 30.85 a | 0.41 ± 0.02 a | 2.24 ± 0.08 a | 2.19 ± 0.09 a | 5.41 ± 0.12 a | 7.41 ± 1.59a |
| N5 | 644.64 ± 28.29 ab | 0.39 ± 0.01 a | 2.03 ± 0.09 ab | 1.99 ± 0.07 ab | 5.26 ± 0.12 a | 7.41 ± 1.63a |
| N10 | 628.45 ± 44.48 b | 0.34 ± 0.03 a | 1.86 ± 0.14 ab | 1.78 ± 0.07 ab | 5.55 ± 0.14 a | 7.56 ± 1.66a |
| N20 | 594.73 ± 39.45 b | 0.39 ± 0.02 a | 1.89 ± 0.11 ab | 1.80 ± 0.11 ab | 4.84 ± 0.33 a | 7.49 ± 1.63a |
| N40 | 542.78 ± 38.80 b | 0.36 ± 0.03 a | 1.71 ± 0.07 b | 1.74 ± 0.13 b | 4.81 ± 0.29 a | 7.48 ± 1.66a |
| Variables | Df | Sum Sq | Mean Sq | F | P |
|---|---|---|---|---|---|
| T | 4 | 32743 | 8186 | 11.596 | 4.86 × 10−7 |
| S | 3 | 1342413 | 447471 | 633.862 | <2 × 10−16 |
| T × S | 12 | 44167 | 3681 | 5.214 | 6.53 × 10−6 |
| Soil Variable | CK | N5 | N10 | N20 | N40 |
|---|---|---|---|---|---|
| SOC (g kg−1) | 19.18 ± 3.11 a | 18.20 ± 2.10 a | 18.76 ± 2.27 a | 16.70 ± 1.04 a | 16.65 ± 0.58 a |
| ROC(g kg−1) | 4.94 ± 0.94 a | 4.19 ± 0.50 a | 5.10 ± 0.42 a | 4.29 ± 0.30 a | 4.20 ± 0.35 a |
| MBC(g kg−1) * | 342.39 ± 31.16 a | 251.58 ± 10.64 b | 275.23 ± 12.46 ab | 273.07 ± 16.98 ab | 247.90 ± 19.07 b |
| SDOC(mg kg−1) * | 159.43 ± 6.45 a | 126.43 ± 8.57 ab | 122.54 ± 9.49 b | 120.43 ± 8.70 b | 124.19 ± 6.26 b |
| HEOC(mg kg−1) | 741.95 ± 64.17 a | 765.79 ± 25.86 a | 816.39 ± 95.62 a | 761.86 ± 49.36 a | 693.87 ± 46.89 a |
| TN (g kg−1) | 1.35 ± 0.20 a | 1.36 ± 0.13 a | 1.44 ± 0.14 a | 1.27 ± 0.08 a | 1.37 ± 0.03 a |
| MBN(mg kg−1) | 57.88 ± 8.50 a | 45.52 ± 4.82 a | 45.48 ± 1.76 a | 50.03 ± 4.16 a | 45.02 ± 2.05 a |
| SDON(mg kg−1) | 26.44 ± 0.64 a | 29.94 ± 0.78 a | 29.97 ± 1.77 a | 28.51 ± 1.41 a | 27.95 ± 0.60 a |
| HEON (mg kg−1) | 67.46 ± 6.20 a | 65.59 ± 2.56 a | 69.35 ± 3.37 a | 66.32 ± 2.98 a | 66.59 ± 2.45 a |
| pH | 6.55 ± 0.23 a | 6.60 ± 0.21 a | 6.51 ± 0.18 a | 6.48 ± 0.28 a | 6.53 ± 0.25 a |
| SOC/TN * | 14.11 ± 0.32 a | 13.30 ± 0.41 ab | 12.90 ± 0.30 ab | 13.13 ± 0.27 ab | 12.16 ± 0.30 b |
| SDOC/SDON ** | 6.03 ± 0.18 a | 4.23 ± 0.31 b | 4.12 ± 0.35 b | 4.23 ± 0.29 b | 4.46 ± 0.31 b |
| HEOC/HEON | 11.02 ± 0.08 a | 11.68 ± 0.13 a | 11.65 ± 0.91 a | 11.46 ± 0.33 a | 10.39 ± 0.33 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Liu, Y.; Zhou, Z.; Zhang, Y. Inorganic Nitrogen Addition Affects Soil Respiration and Belowground Organic Carbon Fraction for a Pinus tabuliformis Forest. Forests 2019, 10, 369. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050369
Zhang H, Liu Y, Zhou Z, Zhang Y. Inorganic Nitrogen Addition Affects Soil Respiration and Belowground Organic Carbon Fraction for a Pinus tabuliformis Forest. Forests. 2019; 10(5):369. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050369
Chicago/Turabian StyleZhang, Huan, Yanhong Liu, Zhiyong Zhou, and Yueying Zhang. 2019. "Inorganic Nitrogen Addition Affects Soil Respiration and Belowground Organic Carbon Fraction for a Pinus tabuliformis Forest" Forests 10, no. 5: 369. https://0-doi-org.brum.beds.ac.uk/10.3390/f10050369