Development of Fine Root Biomass of Two Contrasting Urban Tree Cultivars in Response to Drought Stress


Abstract
:1. Introduction
2. Materials and Methods
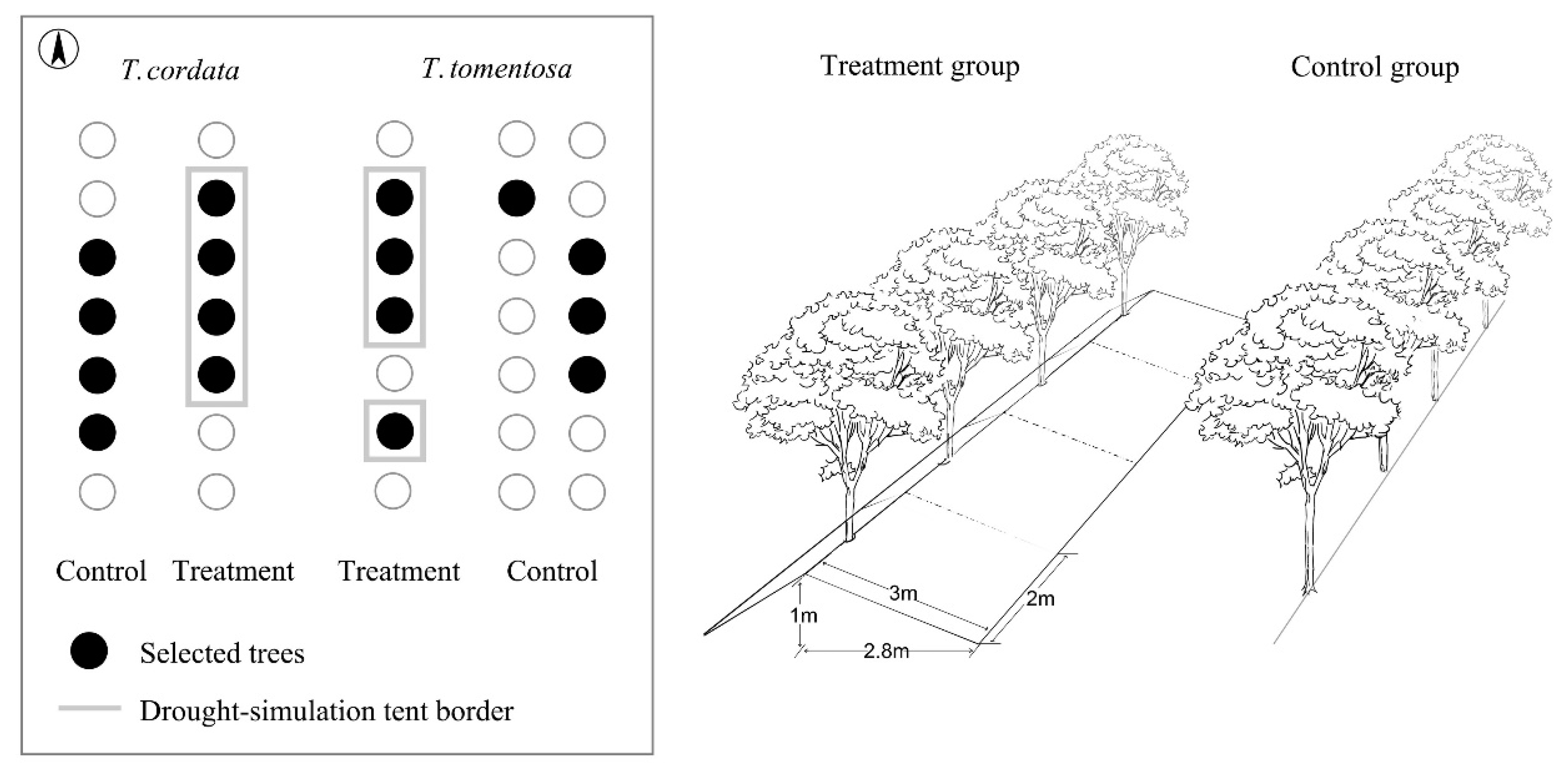
2.1. Site Description and Cultivar Selection
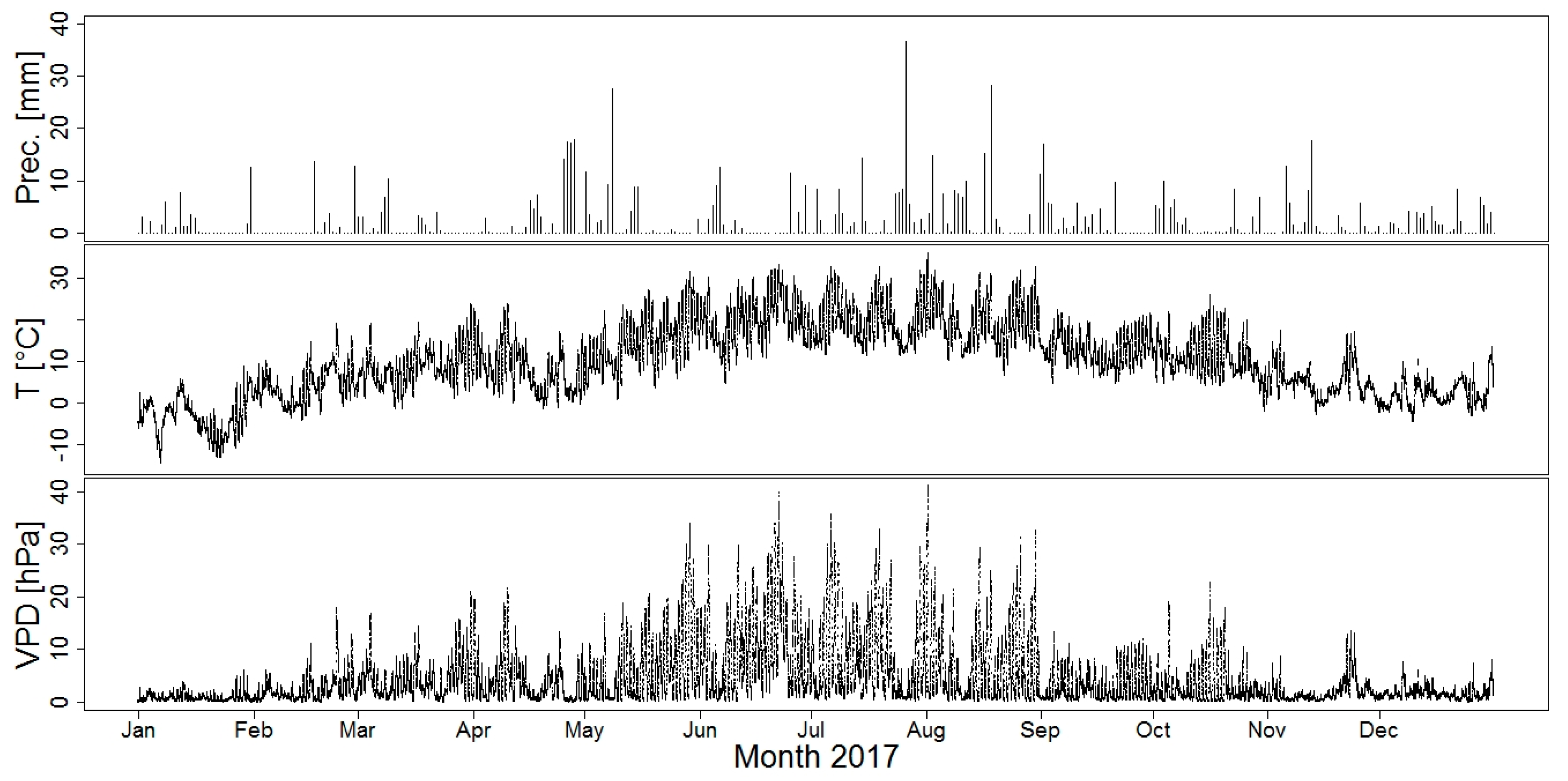
2.2. Climate and Soil Moisture Measurements, Soil Samples
2.3. Drought Experiment in 2017
2.4. Stem Growth and Leaf Area Index (LAI)
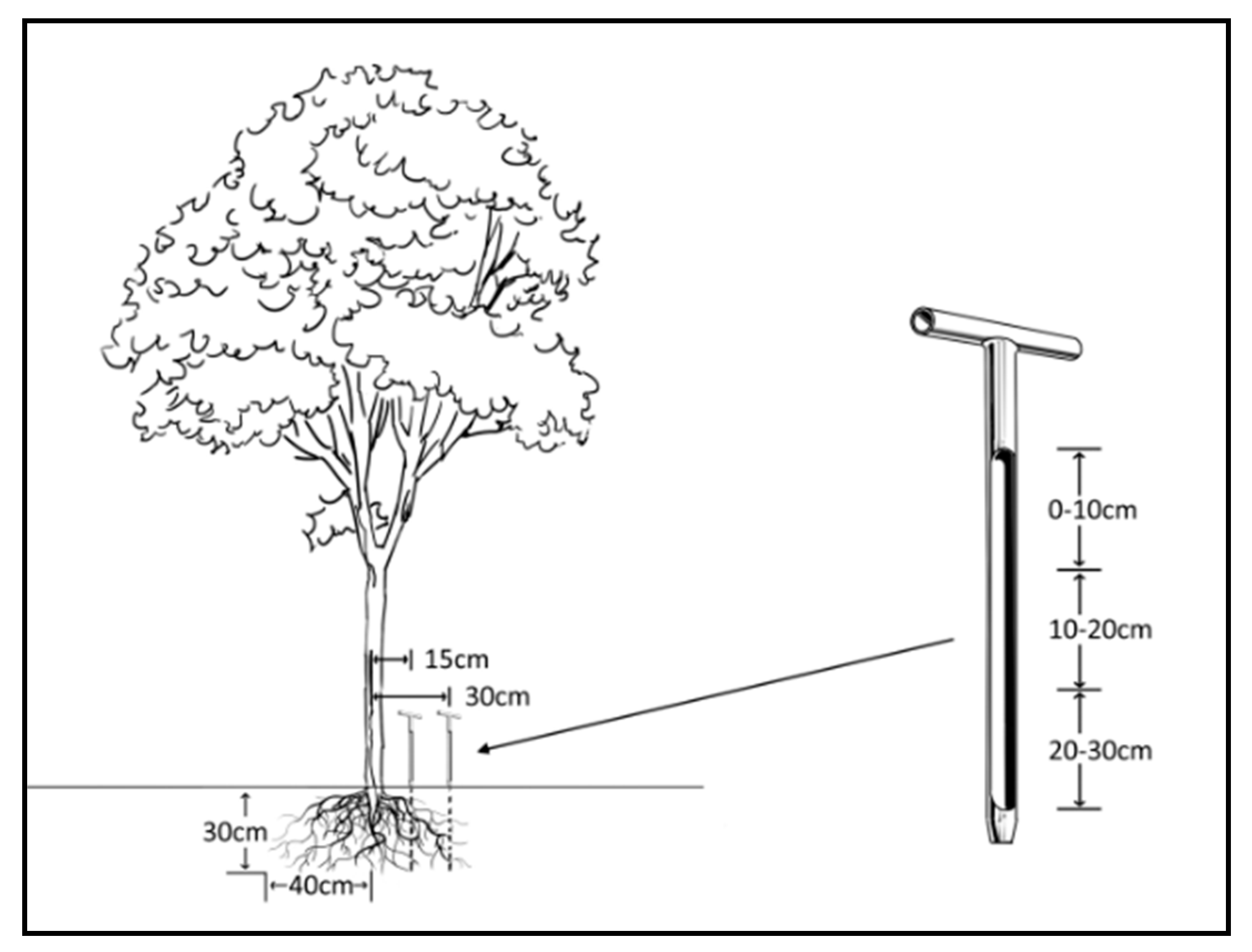
2.5. Coring for Fine Root Biomass
2.6. Statistical Analysis
3. Results
3.1. Weather and Soil Conditions
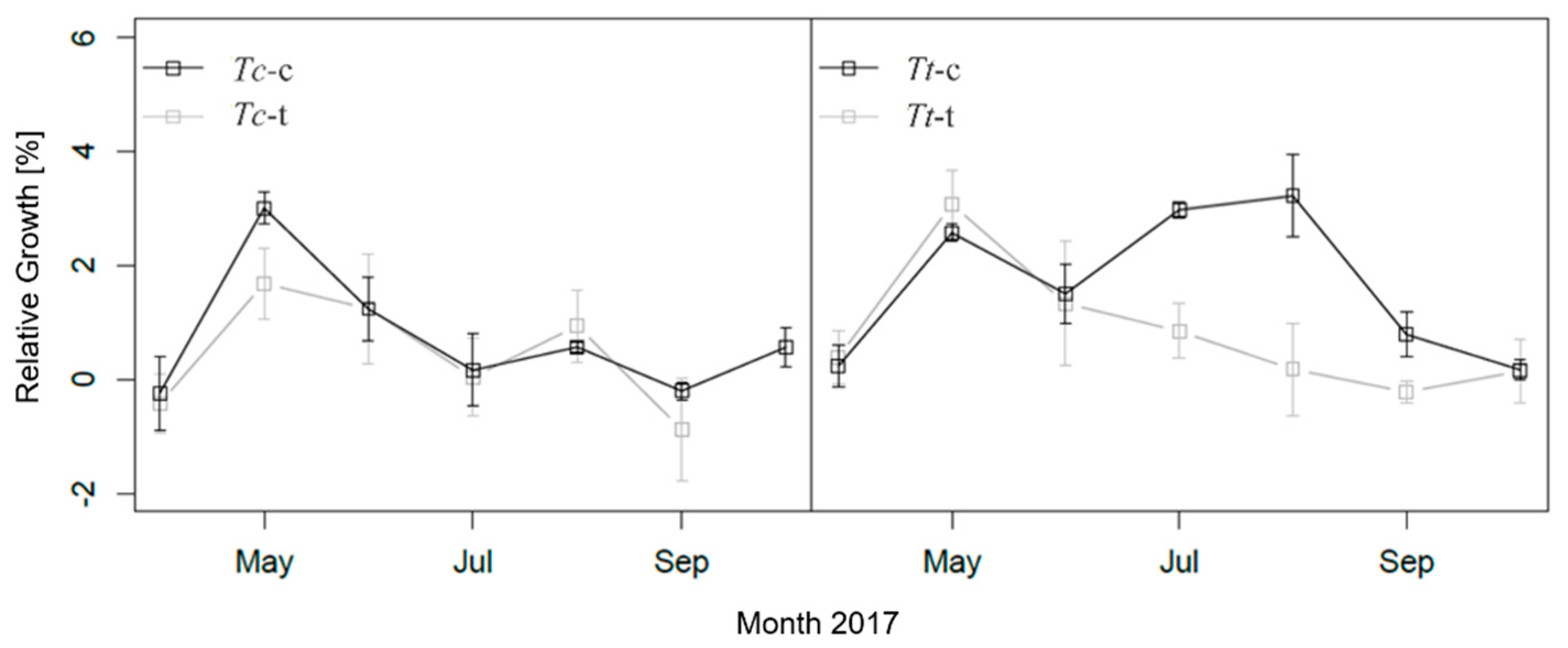
3.2. Relative Stem Growth and LAI
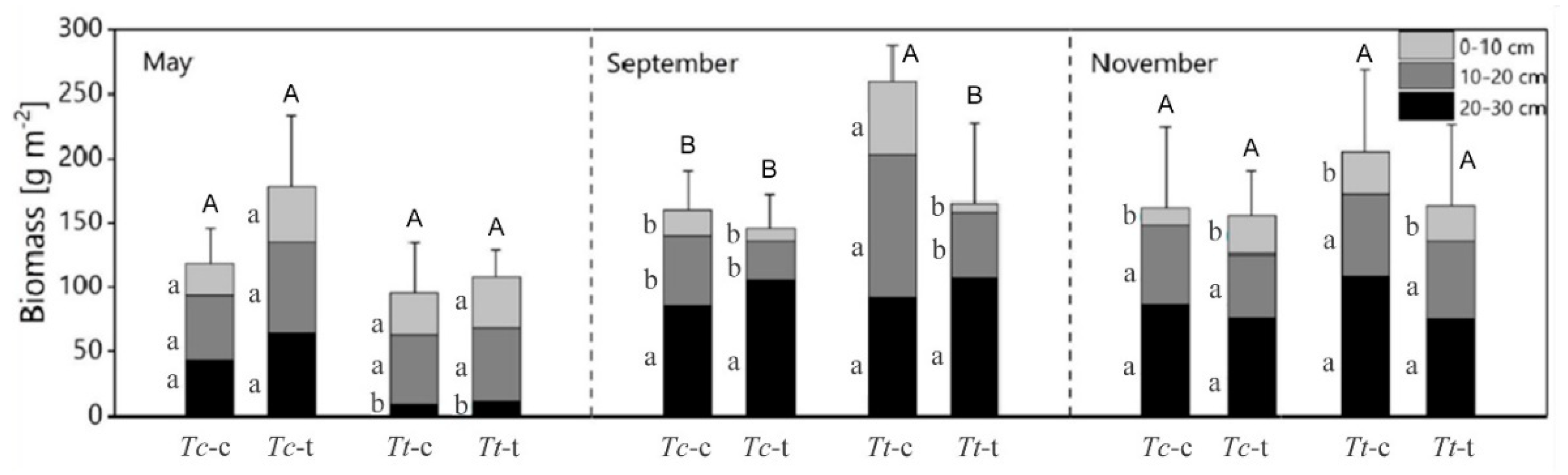
3.3. Total and Vertical Fine Root Biomass
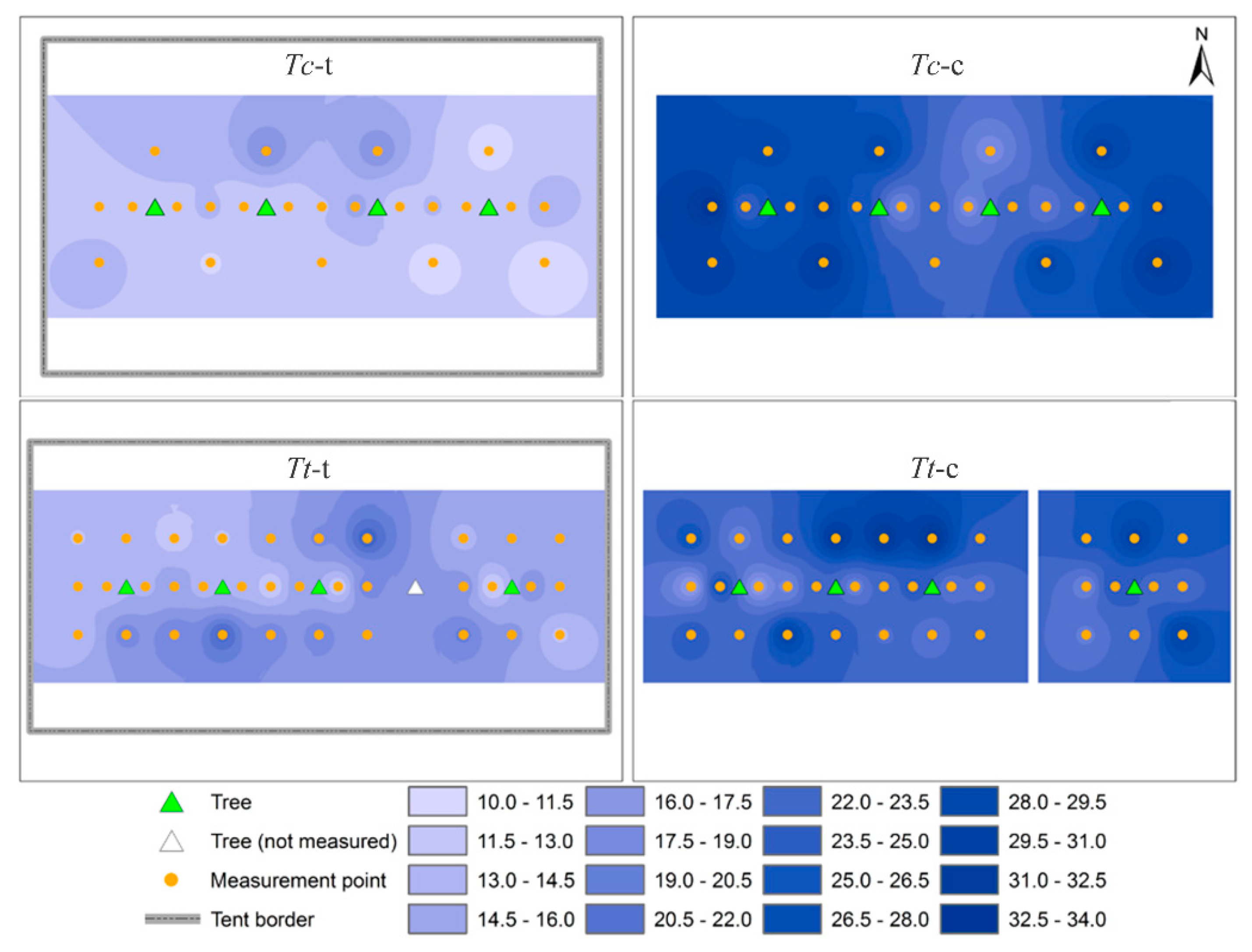
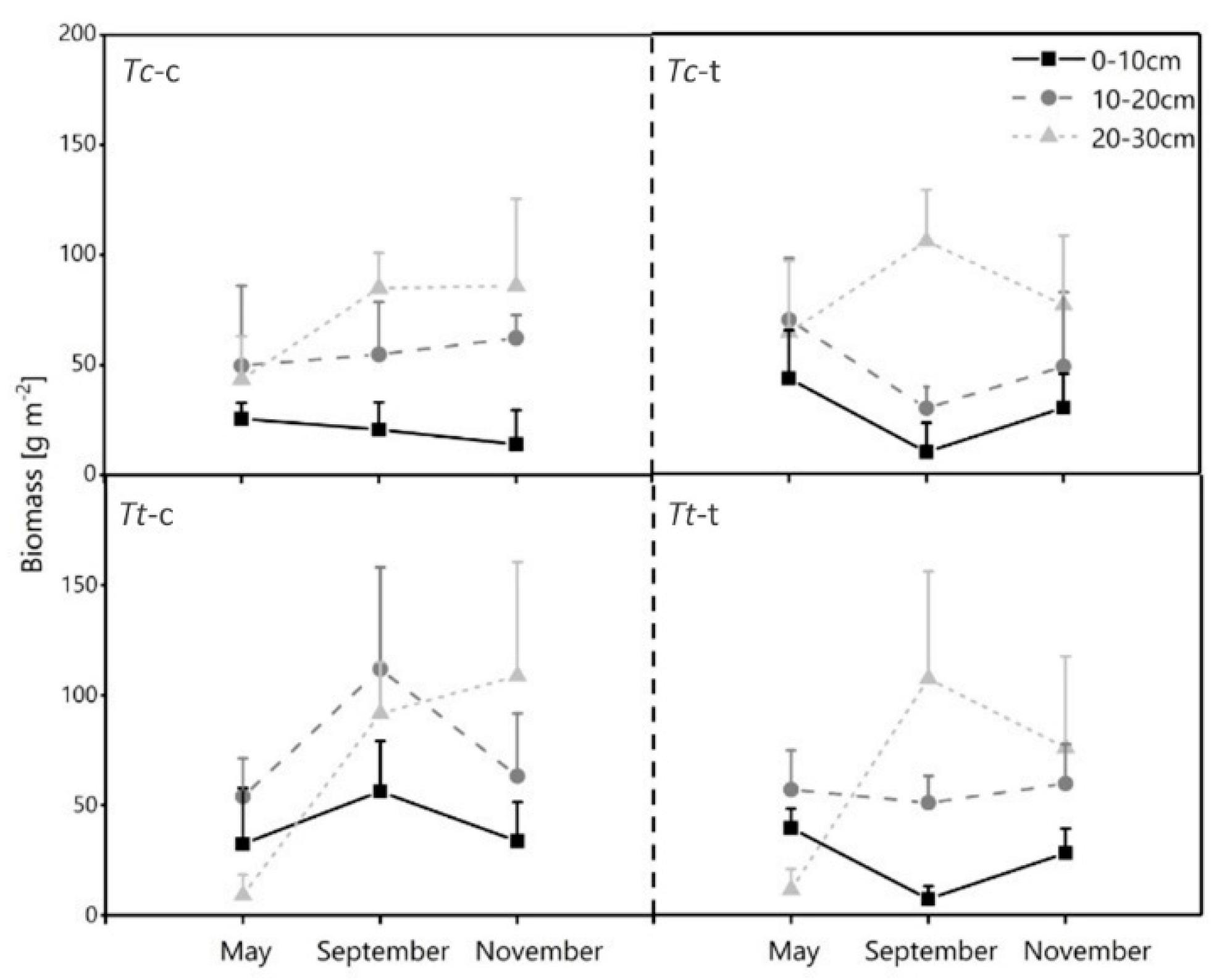
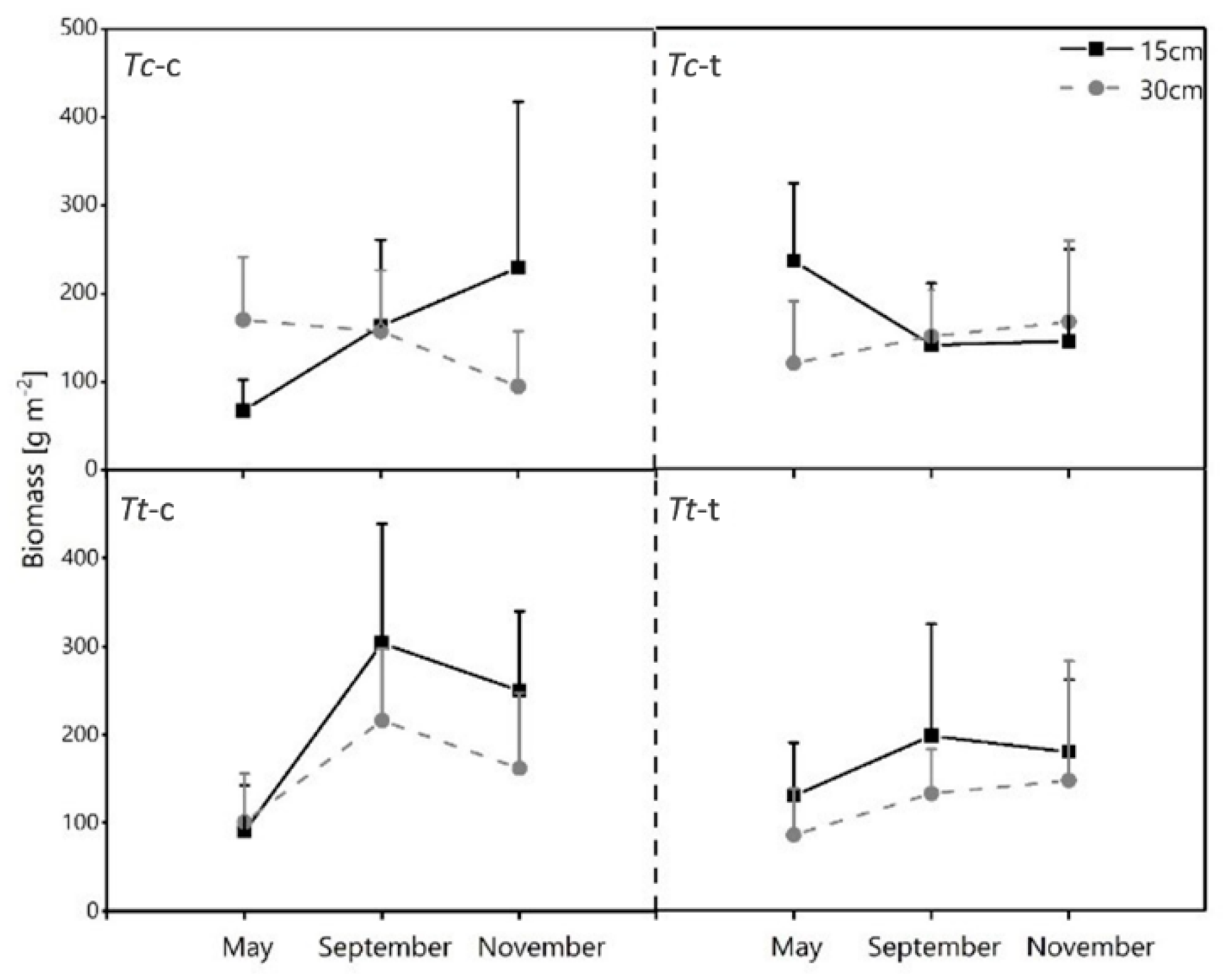
3.4. Horizontal Development of Fine Root Biomass
4. Discussion
4.1. Total Fine Root Biomass
4.2. Vertical Development of Fine Root Biomass
4.3. Horizontal Development of Fine Root Biomass
4.4. Carbon Allocation under Drought Stress
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eisenhauer, N.; Cesarz, S.; Koller, R.; Worm, K.; Reich, P.B. Global change belowground: Impacts of elevated CO2, nitrogen, and summer drought on soil food webs and biodiversity. Glob. Chang. Biol. 2012, 18, 435–447. [Google Scholar] [CrossRef]
- Solomon, S. Climate Change 2007—The Physical Science Basis: Working Group I Contribution to the Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007; Volume 4. [Google Scholar]
- Angel, S.; Parent, J.; Civco, D.L.; Blei, A.; Potere, D. The dimensions of global urban expansion: Estimates and projections for all countries, 2000–2050. Prog. Plan. 2011, 75, 53–107. [Google Scholar] [CrossRef]
- Seto, K.C.; Fragkias, M.; Güneralp, B.; Reilly, M.K. A meta-analysis of global urban land expansion. PLoS ONE 2011, 6, e23777. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional vegetation die-off in response to global-change-type drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberti, M. The Effects of Urban Patterns on Ecosystem Function. Int. Reg. Sci. Rev. 2016, 28, 168–192. [Google Scholar] [CrossRef]
- Stone, B., Jr. The City and the Coming Climate: Climate Change in the Places We Live; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Oleson, K.W.; Monaghan, A.; Wilhelmi, O.; Barlage, M.; Brunsell, N.; Feddema, J.; Hu, L.; Steinhoff, D.F. Interactions between urbanization, heat stress, and climate change. Clim. Chang. 2013, 129, 525–541. [Google Scholar] [CrossRef]
- Sailor, D.J. A review of methods for estimating anthropogenic heat and moisture emissions in the urban environment. Int. J. Climatol. 2011, 31, 189–199. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Schütze, G.; Perkins, D.; Rötzer, T.; Caldentey, J.; Koike, T.; van Con, T. Climate change accelerates growth of urban trees in metropolises worldwide. Sci. Rep. 2017, 7, 15403. [Google Scholar] [CrossRef]
- Gregg, J.W.; Jones, C.G.; Dawson, T.E. Urbanization effects on tree growth in the vicinity of New York City. Nature 2003, 424, 183–187. [Google Scholar] [CrossRef]
- Roy, S.; Byrne, J.; Pickering, C. A systematic quantitative review of urban tree benefits, costs, and assessment methods across cities in different climatic zones. Urban For. Urban Green. 2012, 11, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Alberti, M.; Marzluff, J.M. Ecological resilience in urban ecosystems: Linking urban patterns to human and ecological functions. Urban Ecosyst. 2004, 7, 241–265. [Google Scholar] [CrossRef]
- Arthur, T.E.; Martin, S.D. Street Tree Directory; National Academy of Sciences: Washington, DC, USA, 1981. [Google Scholar]
- Miller, R.W.; Hauer, R.J.; Werner, L.P. Urban Forestry: Planning and Managing Urban Greenspaces; Waveland Press: Waveland, MS, USA, 2015. [Google Scholar]
- Low, N. The Green City: Sustainable Homes, Sustainable Suburbs; UNSW Press: Randwick, Australia, 2005. [Google Scholar]
- Burden, D. 22 Benefits of Urban Street Trees. 2008. Available online: https://www.vibrantcitieslab.com/resources/22-benefits-of-street-trees/ (accessed on 15 January 2020).
- Sutton, M.; Dragosits, U.; Theobald, M.; McDonald, A.; Nemitz, E.; Blyth, J.; Sneath, R.; Williams, A.; Hall, J.; Bealey, W. The role of trees in landscape planning to reduce the impacts of atmospheric ammonia deposition. In Landscape Ecology of Trees and Forests; IALE: Nottingham, UK, 2004; pp. 143–150. [Google Scholar]
- Nowak, D.J.; Greenfield, E.J.; Hoehn, R.E.; Lapoint, E. Carbon storage and sequestration by trees in urban and community areas of the United States. Environ. Pollut. 2013, 178, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rötzer, T.; Seifert, T.; Gayler, S.; Priesack, E.; Pretzsch, H. Effects of stress and defence allocation on tree growth: Simulation results at the individual and stand level. In Growth and Defence in Plants; Springer: Berlin/Heidelberg, Germany, 2012; pp. 401–432. [Google Scholar]
- Moser, A.; Rötzer, T.; Pauleit, S.; Pretzsch, H. The urban environment can modify drought stress of small-leaved lime (Tilia cordata Mill.) and black locust (Robinia pseudoacacia L.). Forests 2016, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Neill, C. Comparison of soil coring and ingrowth methods for measuring belowground production. Ecology 1992, 73, 1918–1921. [Google Scholar] [CrossRef]
- Hendrick, R.L.; Pregitzer, K.S. The dynamics of fine root length, biomass, and nitrogen content in two northern hardwood ecosystems. Can. J. For. Res. 1993, 23, 2507–2520. [Google Scholar] [CrossRef]
- Vogt, K.A.; Vogt, D.J.; Palmiotto, P.A.; Boon, P.; O’Hara, J.; Asbjornsen, H. Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species. Plant Soil 1995, 187, 159–219. [Google Scholar] [CrossRef]
- Curtis, P.S.; Zak, D.R.; Pregitzer, K.S.; Lussenhop, J.; Teeri, J.A. Linking above-and belowground responses to rising CO 2 in northern deciduous forest species. In Carbon Dioxide and Terrestrial Ecosystems; Elsevier: Amsterdam, The Netherlands, 1996; pp. 41–51. [Google Scholar]
- Thornley, J. A balanced quantitative model for root: Shoot ratios in vegetative plants. Ann. Bot. 1972, 36, 431–441. [Google Scholar] [CrossRef]
- Zhang, C.; Stratopoulos, L.M.F.; Pretzsch, H.; Rötzer, T. How Do Tilia cordata Greenspire Trees Cope with Drought Stress Regarding Their Biomass Allocation and Ecosystem Services? Forests 2019, 10, 676. [Google Scholar] [CrossRef] [Green Version]
- Lyr, H.; Hoffmann, G. Growth rates and growth periodicity of tree roots. In International Review of Forestry Research; Elsevier: Amsterdam, The Netherlands, 1967; Volume 2, pp. 181–236. [Google Scholar]
- Borchert, R. Soil and stem water storage determine phenology and distribution of tropical dry forest trees. Ecology 1994, 75, 1437–1449. [Google Scholar] [CrossRef]
- Green, J.; Dawson, L.; Proctor, J.; Duff, E.; Elston, D. Fine root dynamics in a tropical rain forest is influenced by rainfall. Plant Soil 2005, 276, 23–32. [Google Scholar] [CrossRef]
- Stone, E.L.; Kalisz, P.J. On the maximum extent of tree roots. For. Ecol. Manag. 1991, 46, 59–102. [Google Scholar] [CrossRef]
- Zwieniecki, M.A.; Newton, M. Roots growing in rock fissures: Their morphological adaptation. Plant Soil 1995, 172, 181–187. [Google Scholar] [CrossRef]
- De Sousa, C.A. Turning brownfields into green space in the City of Toronto. Landsc. Urban Plan. 2003, 62, 181–198. [Google Scholar] [CrossRef]
- Asch, F.; Dingkuhn, M.; Sow, A.; Audebert, A. Drought-induced changes in rooting patterns and assimilate partitioning between root and shoot in upland rice. Field Crops Res. 2005, 93, 223–236. [Google Scholar] [CrossRef]
- Stratópoulos, L.M.F.; Zhang, C.; Häberle, K.H.; Pauleit, S.; Duthweiler, S.; Pretzsch, H.; Rötzer, T. Effects of Drought on the Phenology, Growth, and Morphological Development of Three Urban Tree Species and Cultivars. Sustainability 2019, 11, 5117. [Google Scholar] [CrossRef] [Green Version]
- Girardet, H. The Gaia Atlas of Cities: New Directions for Sustainable Urban Living; UN-HABITAT: Nairobi, Kenya, 1996. [Google Scholar]
- Hough, M. Cities and Natural Process; Routledge: Abington, UK, 2004. [Google Scholar]
- Register, R. Ecocities: Rebuilding Cities in Balance with Nature; New Society Publishers: Gabriolav, BC, Canada, 2006. [Google Scholar]
- Newman, P.; Jennings, I. Cities as Sustainable Ecosystems: Principles and Practices; Island Press: Washington, DC, USA, 2012. [Google Scholar]
- Meier, I.C.; Leuschner, C. Belowground drought response of European beech: Fine root biomass and carbon partitioning in 14 mature stands across a precipitation gradient. Glob. Chang. Biol. 2008, 14, 2081–2095. [Google Scholar] [CrossRef]
- Rowell, D.P.; Jones, R.G. Causes and uncertainty of future summer drying over Europe. Clim. Dyn. 2006, 27, 281–299. [Google Scholar] [CrossRef]
- Moser, A.; Rahman, M.A.; Pretzsch, H.; Pauleit, S.; Rötzer, T. Inter-and intraannual growth patterns of urban small-leaved lime (Tilia cordata mill.) at two public squares with contrasting microclimatic conditions. Int. J. Biometeorol. 2017, 61, 1095–1107. [Google Scholar] [CrossRef]
- Solomon, S. Climate Change 2007: The Physical Science Basis; Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2007; Volume 996. [Google Scholar]
- Pauleit, S.; Jones, N.; Nyhuus, S.; Pirnat, J.; Salbitano, F. Urban forest resources in European cities. In Urban Forests and Trees; Springer: Berlin/Heidelberg, Germany, 2005; pp. 49–80. [Google Scholar]
- Bühler, O.; Balder, H.; Kristoffersen, F. Establishment of urban trees. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2009, 4, 1–11. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Radoglou, K.; Dobrowolska, D.; Spyroglou, G.; Nicolescu, V.N. A review on the ecology and silviculture of limes (Tilia cordata Mill., Tilia platyphyllos Scop. and Tilia tomentosa Moench.) in Europe. Romania 2008, 15, 16. [Google Scholar]
- Stratópoulos, L.M.F.; Zhang, C.; Duthweiler, S.; Häberle, K.H.; Rötzer, T.; Xu, C.; Pauleit, S. Tree species from two contrasting habitats for use in harsh urban environments respond differently to extreme drought. Int. J. Biometeorol. 2019, 63, 197–208. [Google Scholar] [CrossRef]
- Goisser, M.; Geppert, U.; Rötzer, T.; Paya, A.; Huber, A.; Kerner, R.; Bauerle, T.; Pretzsch, H.; Pritsch, K.; Häberle, K. Does belowground interaction with Fagus sylvatica increase drought susceptibility of photosynthesis and stem growth in Picea abies? For. Ecol. Manag. 2016, 375, 268–278. [Google Scholar] [CrossRef]
- Leuschner, C.; Hertel, D.; Schmid, I.; Koch, O.; Muhs, A.; Hölscher, D. Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility. Plant Soil 2004, 258, 43–56. [Google Scholar] [CrossRef]
- Gillner, S.; Bräuning, A.; Roloff, A. Dendrochronological analysis of urban trees: Climatic response and impact of drought on frequently used tree species. Trees 2014, 28, 1079–1093. [Google Scholar] [CrossRef]
- Gewin, V. Food: An underground revolution. Nat. News 2010, 466, 552–553. [Google Scholar] [CrossRef]
- Pierret, A.; Maeght, J.L.; Clément, C.; Montoroi, J.P.; Hartmann, C.; Gonkhamdee, S. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef] [Green Version]
- Schenk, H.J. The Shallowest Possible Water Extraction Profile: A Null Model for Global Root Distributions. Vadose Zone J. 2008, 7, 1119–1124. [Google Scholar] [CrossRef]
- Schenk, H.J. Root competition: Beyond resource depletion. J. Ecol. 2006, 94, 725–739. [Google Scholar] [CrossRef]
- Joslin, J.; Gaudinski, J.B.; Torn, M.S.; Riley, W.; Hanson, P.J. Fine-root turnover patterns and their relationship to root diameter and soil depth in a 14C-labeled hardwood forest. New Phytol. 2006, 172, 523–535. [Google Scholar] [CrossRef]
- Germon, A.; Cardinael, R.; Prieto, I.; Mao, Z.; Kim, J.; Stokes, A.; Dupraz, C.; Laclau, J.P.; Jourdan, C. Unexpected phenology and lifespan of shallow and deep fine roots of walnut trees grown in a silvoarable Mediterranean agroforestry system. Plant Soil 2016, 401, 409–426. [Google Scholar] [CrossRef]
- Abramoff, R.Z.; Finzi, A.C. Are above-and below-ground phenology in sync? New Phytol. 2015, 205, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- McCormack, M.L.; Guo, D. Impacts of environmental factors on fine root lifespan. Front. Plant Sci. 2014, 5, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Moser, A.; Rötzer, T.; Pauleit, S. Microclimatic differences and their influence on transpirational cooling of Tilia cordata in two contrasting street canyons in Munich, Germany. Agric. For. Meteorol. 2017, 232, 443–456. [Google Scholar] [CrossRef]
- Schipka, F. Blattwasserzustand und Wasserumsatz von Vier Buchenwäldern Entlang Eines Niederschlagsgradienten in Mitteldeutschland; Niedersächsische Staats-und Universitätsbibliothek Göttingen: Göttingen, Germany, 2003. [Google Scholar]
- Rötzer, T.; Seifert, T.; Pretzsch, H. Modelling above and below ground carbon dynamics in a mixed beech and spruce stand influenced by climate. Eur. J. For. Res. 2009, 128, 171–182. [Google Scholar] [CrossRef]
- Klein, T. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Funct. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef]
- Rötzer, T.; Biber, P.; Moser, A.; Schäfer, C.; Pretzsch, H. Stem and root diameter growth of European beech and Norway spruce under extreme drought. For. Ecol. Manag. 2017, 406, 184–195. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Fine Sand % | Fine Silt % | Grit % | Coarse Grit % | Medium Sand % | Medium Silt % |
|---|---|---|---|---|---|---|---|
| Tc-c | 4 | 15.8 ± 3.6 | 6.5 ± 1.1 | 9.0 ± 2.2 | 16.0 ± 1.2 | 27.3 ± 4.9 | 8.7 ± 3.3 |
| Tc-t | 4 | 22.6 ± 5.9 | 3.8 ± 1.3 | 7.7 ± 4.3 | 12.2 ± 2.7 | 33.4 ± 4.7 | 8.1 ± 1.3 |
| Tt-c | 4 | 14.2 ± 0.4 | 5.7 ± 2.5 | 11.2 ± 0.7 | 10.8 ± 2.3 | 23.7 ± 1.8 | 14.0 ± 4.3 |
| Tt-t | 4 | 14.0 ± 0.7 | 7.3 ± 0.9 | 11.6 ± 0.5 | 12.7 ± 2.3 | 23.1 ± 2.0 | 11.2 ± 1.0 |
| Cultivar | Group | n | LAI (m2 m−2 ± SD) |
|---|---|---|---|
| T. cordata | Control | 4 | 2.53 ± 0.49 |
| Treatment | 4 | 2.13 ± 0.48 | |
| T. tomentosa | Control | 4 | 2.92 ± 0.21 |
| Treatment | 4 | 3.11 ± 0.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Stratópoulos, L.M.F.; Xu, C.; Pretzsch, H.; Rötzer, T. Development of Fine Root Biomass of Two Contrasting Urban Tree Cultivars in Response to Drought Stress. Forests 2020, 11, 108. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010108
Zhang C, Stratópoulos LMF, Xu C, Pretzsch H, Rötzer T. Development of Fine Root Biomass of Two Contrasting Urban Tree Cultivars in Response to Drought Stress. Forests. 2020; 11(1):108. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010108
Chicago/Turabian StyleZhang, Chi, Laura Myrtiá Faní Stratópoulos, Chao Xu, Hans Pretzsch, and Thomas Rötzer. 2020. "Development of Fine Root Biomass of Two Contrasting Urban Tree Cultivars in Response to Drought Stress" Forests 11, no. 1: 108. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010108