Modeling Biomass and Nutrients in a Eucalyptus Stand in the Cerrado

 ,
, 
 ,
,  , , ,
, , ,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
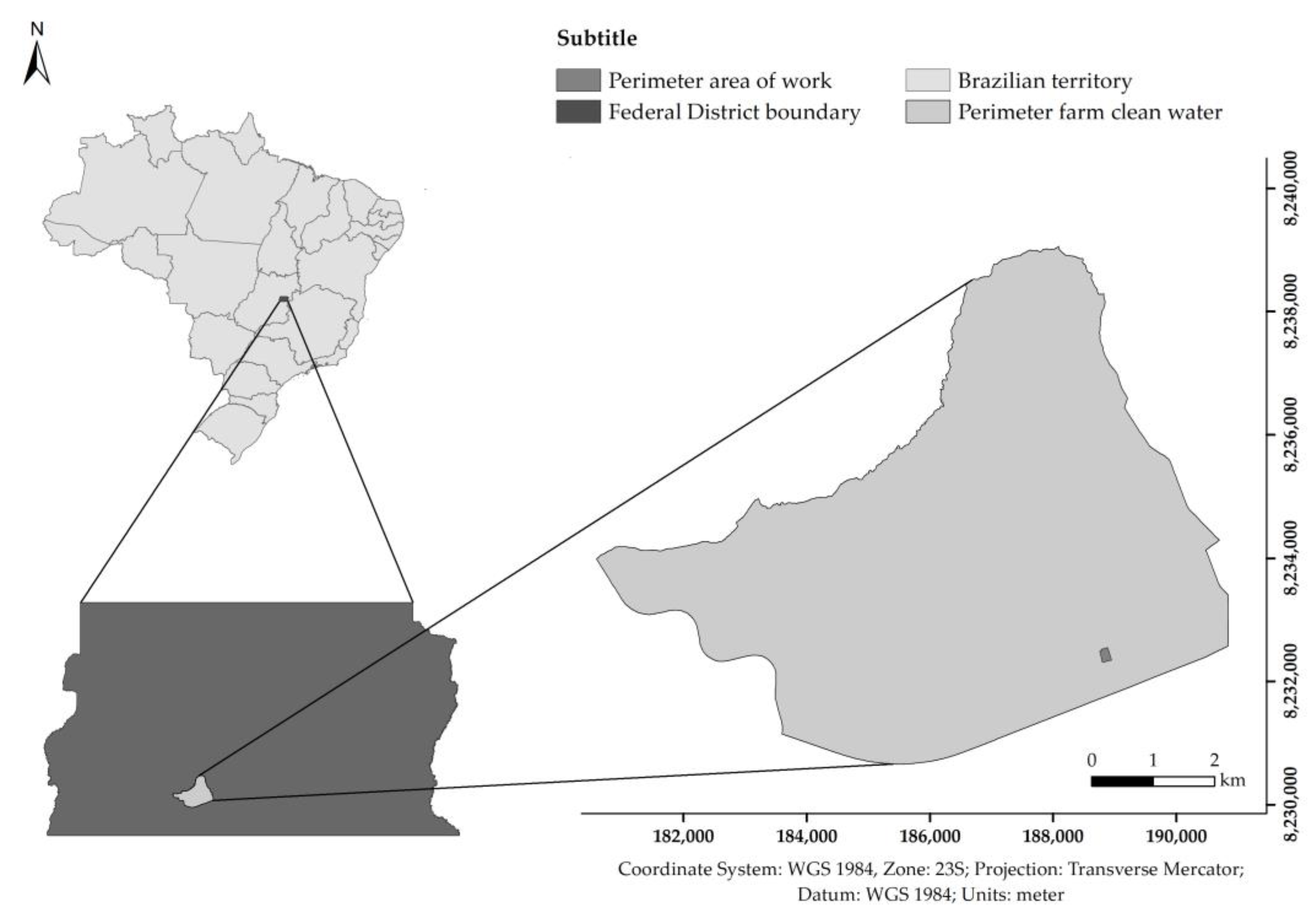
2.1. Study Area
2.2. Data Collection
2.3. Volume Estimate, Biomass, and Nutrient Values
2.4. Modeling of Biomass and Nutrients
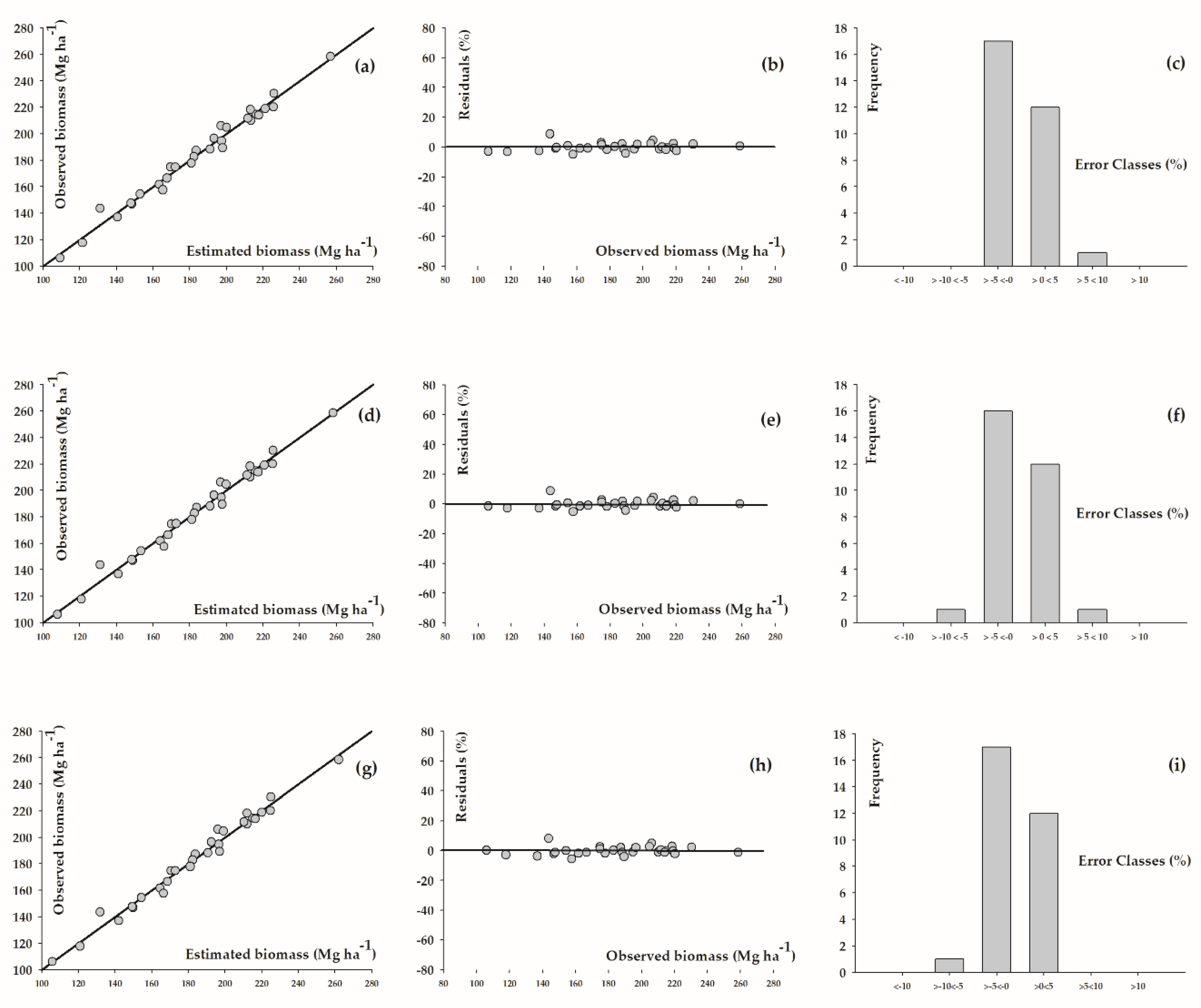
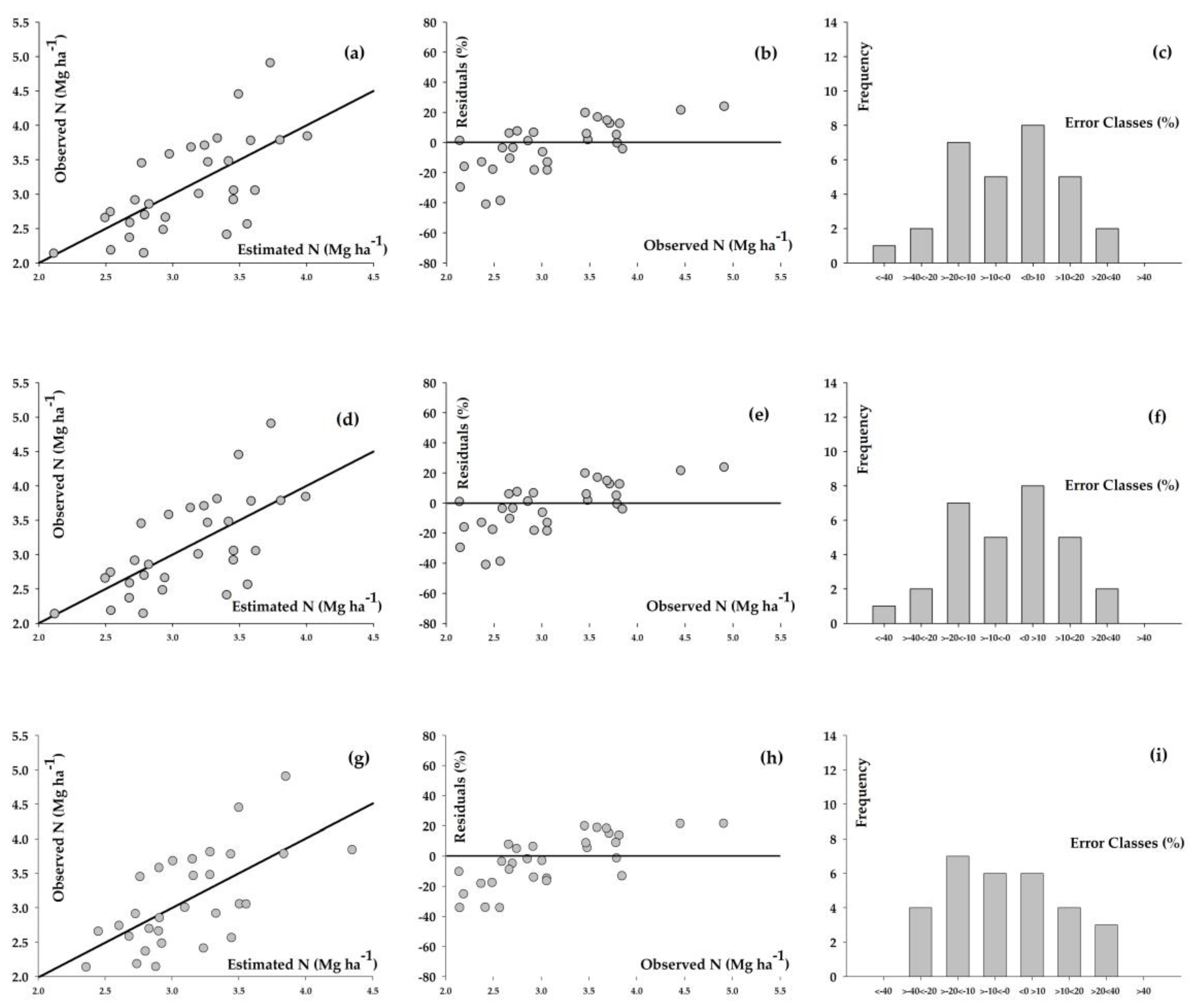
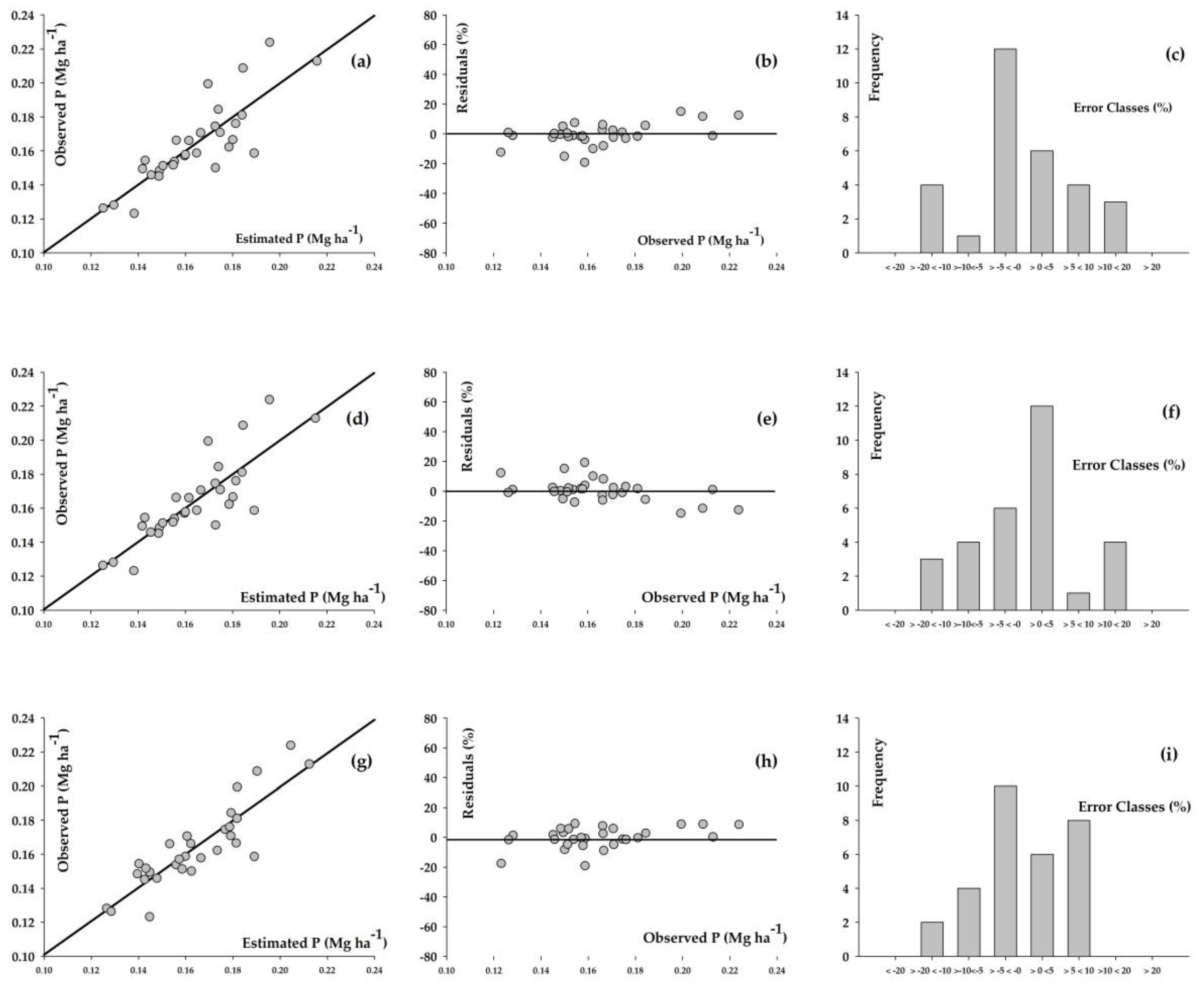
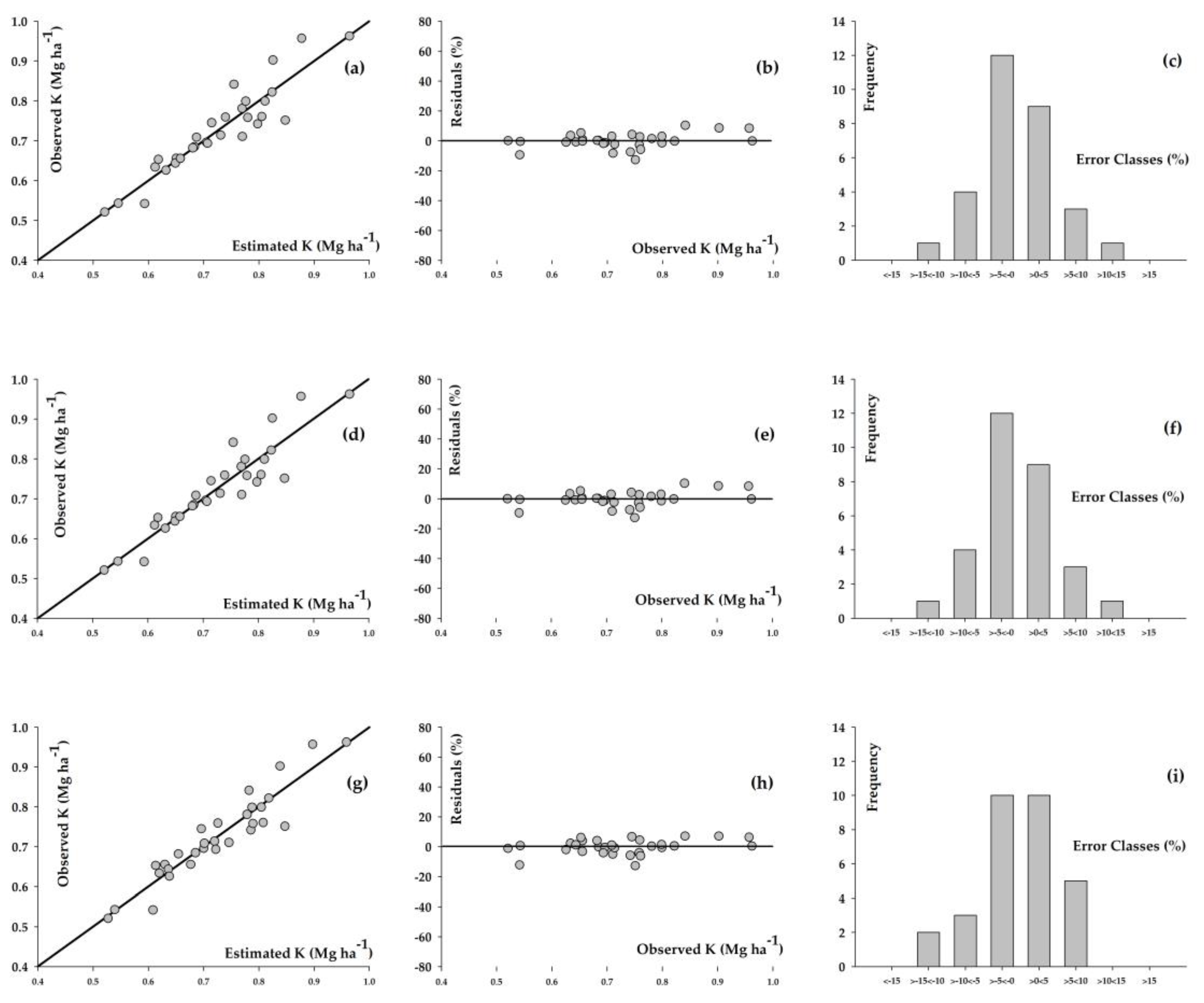
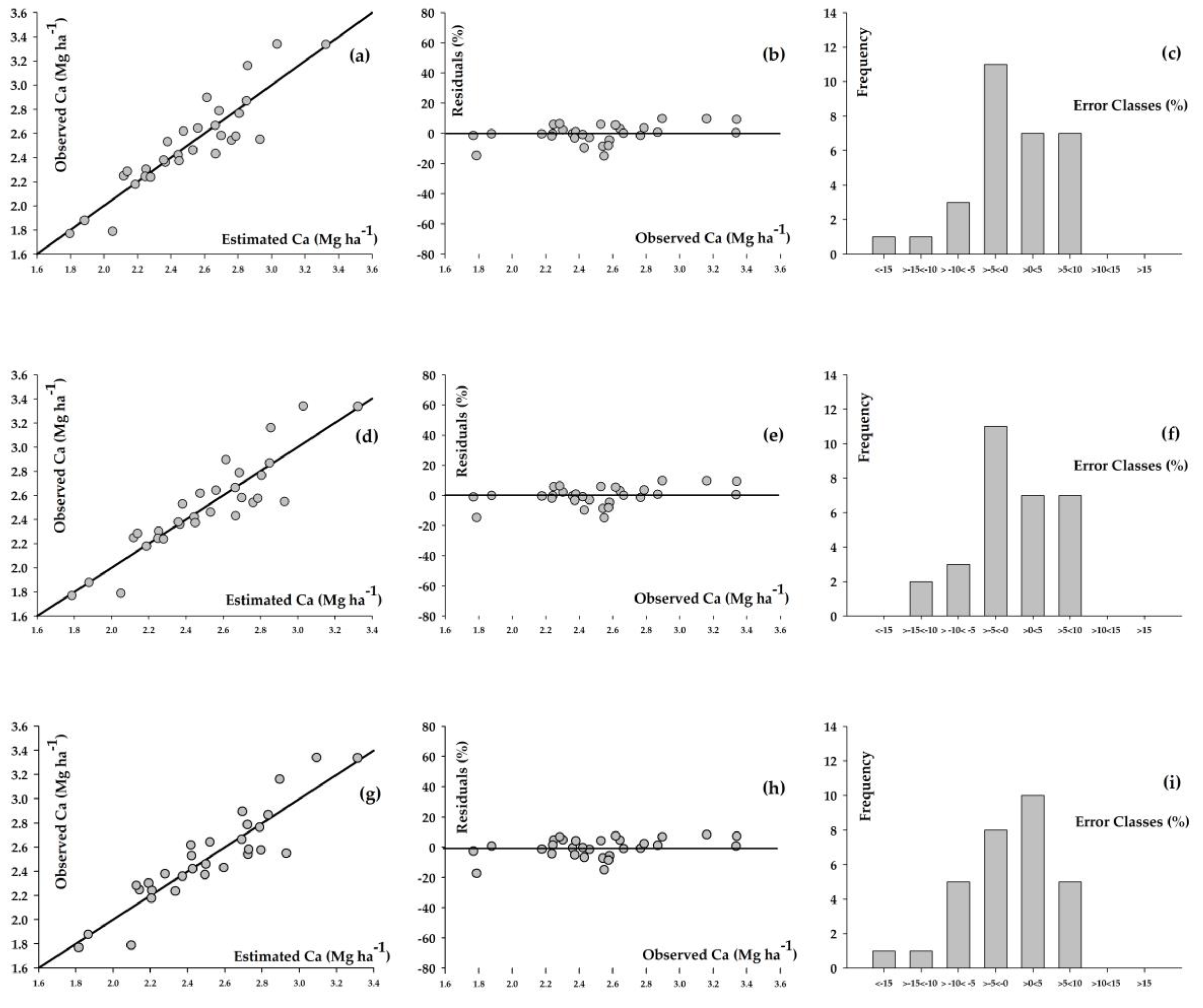
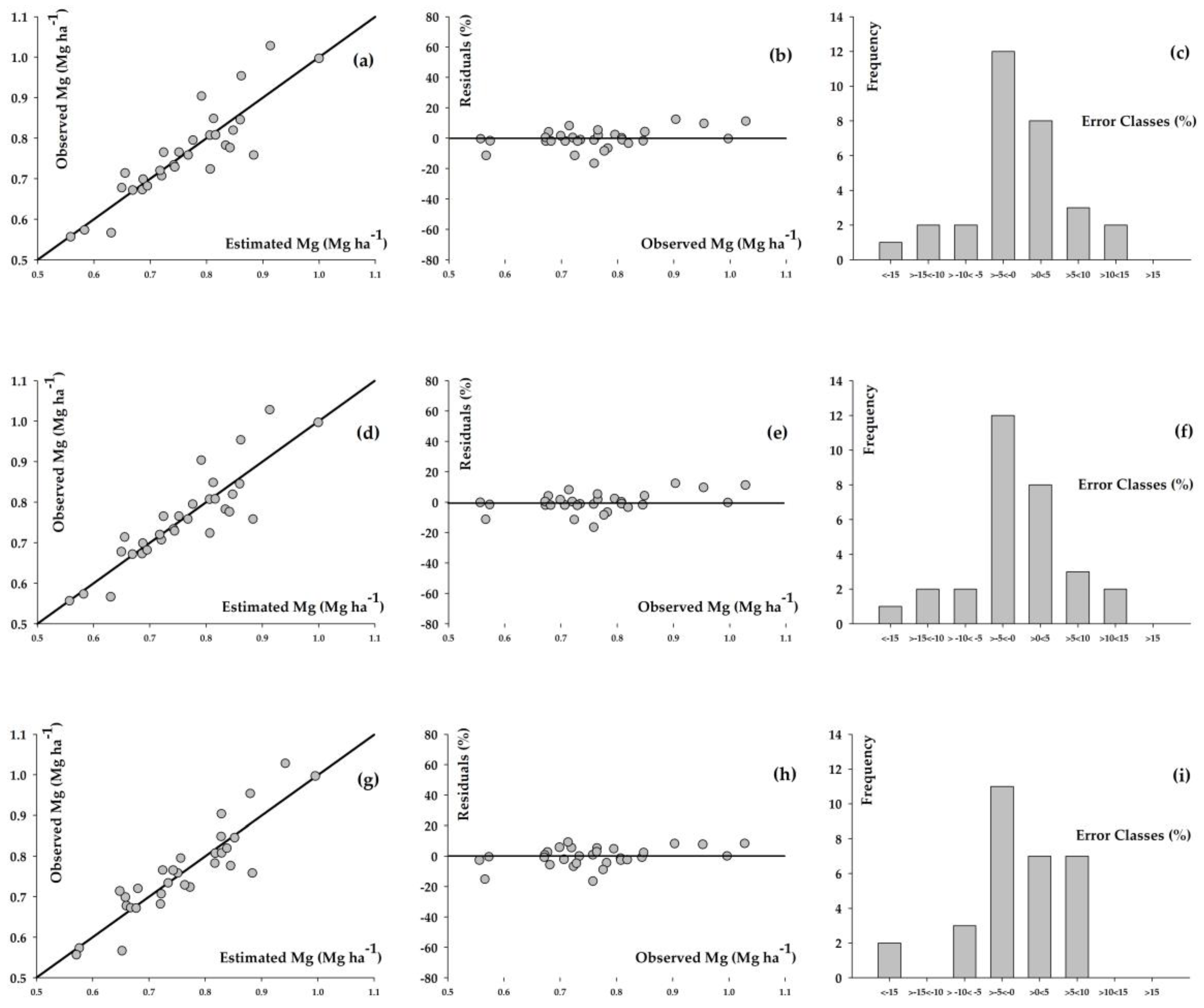
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IBÁ-Industria Brasileira de Árvores. Relatório 2019. Available online: https://iba.org/datafiles/publicacoes/relatorios/iba-relatorioanual2019.pdf (accessed on 25 March 2020).
- Gonçalves, J.L.M.; Alvares, C.A.; Higa, A.R.; Silva, E.V.; Alfenas, A.C.; Stahl, J.; Ferraz, S.F.B.; Lima, W.P.; Brancalion, P.H.S.; Hubner, A.; et al. Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalypt plantations. Ecol. Manag. 2013, 301, 6–27. [Google Scholar] [CrossRef]
- Colodette, J.L.; Gomes, C.M.; Gomes, F.J.; Cabral, C.P. The Brazilian wood biomass supply and utilization focusing on eucalypt. Chem. Biol. Technol. Ag. 2014, 25, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, Y.; Xu, D.; Leininger, T.D.; Zhang, N. A system dynamic model to estimate hydrological processes and water use in a eucalypt plantation. Ecol. Eng. 2016, 86, 290–299. [Google Scholar] [CrossRef]
- Versini, A.; Mareschal, L.; Matsoumbou, T.; Zeller, B.; Ranger, J.; Laclau, J.P. Effects of litter manipulation in a tropical Eucalyptus plantation on leaching of mineral nutrients, dissolved organic nitrogen and dissolved organic carbon. Geoderma 2014, 232–234, 426–436. [Google Scholar] [CrossRef]
- Gatto, A.; Bussinguer, A.P.; Ribeiro, F.C.; Azevedo, G.B.; Bueno, C.B.; Monteiro, M.M.; Souza, P.F. Ciclagem e balanço de nutrientes no sistema solo-planta em um plantio de Eucalyptus sp., no Distrito Federal. R. Bras. Ciênc. Solo 2014, 38, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, G.W.D.; Soares, E.M.B.; Oliveira, F.C.C.; Silva, I.R.; Dungait, J.A.J.; Souza, I.F.; Vergütz, L. Nutrient release from decomposing Eucalyptus harvest residues following simulated management practices in multiple sites in Brazil. Ecol. Manag. 2016, 370, 1–11. [Google Scholar] [CrossRef]
- Richards, A.E.; Forrester, D.I.; Bauhus, J.; Scherer-Lorenzen, M. The influence of mixed tree plantations on the nutrition of individual species: A review. Tree Physiol. 2010, 30, 1192–1208. [Google Scholar] [CrossRef] [Green Version]
- Skorupa, A.L.A.; Barros, N.F.; Neves, J.L.C. Forest litter decomposition as affected by eucalyptus stand age and topography in south-eastern Brazil. Rev. Árvore 2015, 39, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, T.L.; Mello, A.A.; Ferreira, R.A.; Sanquetta, C.R.; Nakajima, N.Y. Equações hipsométricas e volumétricas para um povoamento de Eucalyptus sp. localizado na FLONA do Ibura, Sergipe. Rev. Bras. Ciênc. Agrár. 2011, 6, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Persson, H.J.; Ståhl, G. Characterizing uncertainty in forest remote sensing studies. Remote Sens. 2020, 12, 505. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Feng, Z.; Chen, P.; Chen, X. Development of a Tree Growth Difference Equation and Its Application in Forecasting the Biomass Carbon Stocks of Chinese Forests in 2050. Forests 2019, 10, 582. [Google Scholar] [CrossRef] [Green Version]
- Schikowski, A.B.; Corte, A.P.D.; Sanquetta, C.R. Modelagem do crescimento e de biomassa individual de Pinus. Pesq. Flor. Bras. 2013, 33, 269–278. [Google Scholar] [CrossRef]
- Sedmák, R.; Scheer, L. Modelling of tree diameter growth using growth functions parameterised by least squares and Bayesian methods. J. Sci. 2012, 58, 245–252. [Google Scholar]
- Martins, R.M.; Leite, M.V.S.; Cabacinha, C.D.; Assis, A.L. Teste de identidade de modelos volumétricos para povoamentos de Eucalyptus sp. em sete municípios de Minas Gerais. Enciclopédia Biosf. 2015, 11, 18–33. [Google Scholar]
- Nunes, J.S.; Soares, T.S. Estimativas volumétricas para um povoamento adensado de Eucalyptus sp. em regime de curta rotação. J. Neotrop. Agric. 2017, 4, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Ramos, J.H.; Santos-Posadas, H.M.; Lazalde, J.R.V.; Urias, J.C.T.; Pérez, G.Á.; Hernández-Ramos, H.; Méndez-Lopez, B.; Peduzzi, A. Estimación del volumen comercial en plantaciones de Eucalyptus urophylla con modelos de volumen total y de razón. Agrociencia 2017, 51, 561–580. [Google Scholar]
- Sanquetta, C.R.; Wojciechowski, J.; Corte, D.A.P.; Behling, A.; Netto, S.P.; Rodrigues, A.L.; Sanquetta, M.N.I. Comparison of data mining and allometric model in estimation of tree biomass. Bioinformatics 2015, 16, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, M.C.D.; Barreto-Garcia, P.A.B.; Sanquetta, C.R.; Novaes, A.B.; Melo, L.C. Biomass and carbon stock from Pinus caribaea var. hondurensis under homogenous stands in southwest Bahia, Brazil. Cienc. Rural 2016, 46, 957–962. [Google Scholar] [CrossRef] [Green Version]
- Mendes, T.R.S.; Miguel, E.P.; Vasconcelos, P.G.A.; Valadão, M.B.X.; Rezende, A.V.; Matricardi, E.A.T.; Angelo, H.; Gatto, A.; Nappo, M.E. Use of aerial image in the estimation of volume and biomass of Eucalyptus sp. forest stand. Aust. J. Crop. Sci. 2020, 14, 286–294. [Google Scholar] [CrossRef]
- Vanclay, J.K. Modeling Forest Growth and Yield: Applications to Mixed Tropical Forests, 1st ed.; CABI: Wallingford, UK, 1994; pp. 154–196. [Google Scholar]
- Pereira, A.R.S.; Cordeiro, M.A.; Abreu, J.C.; Santos, R.O.; Natalino, J.; Silva, M. Modelagem volumétrica para Eucalyptus urograndis no Município de Porto Grande, Amapá, Brasil. Biota Amaz. 2016, 6, 10–14. [Google Scholar] [CrossRef]
- Machado, S.A.; Figura, M.A.; Silva, L.C.R.; Téo, S.J.; Stolle, L.; Urbano, E. Modelagem volumétrica para bracatinga (Mimosa scabrella) em bracatingais nativos. Pesq. Flor. Bras. 2008, 56, 17–29. [Google Scholar]
- Assis, C.O.; Trugilho, P.F.; Goulart, S.L.; Assis, M.R.; Bianchi, M.L. Efeito da aplicação de nitrogênio na produção e qualidade da madeira e carvão vegetal de um híbrido de Eucalyptus grandis × Eucalyptus urophylla. Floresta Ambient. 2018, 25, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Pertille, C.T.; Sanquetta, C.R.; Nicoletti, M.F.; Topanotti, L.R. Volume prediction through form factor and regression models by age class for Pinus taeda L. Adv. Sci. 2018, 5, 431–436. [Google Scholar]
- Leal, F.A.; Cabacinha, C.D.; Castro, R.V.O.; Matricardi, E.A.T. Amostragem de árvores de Eucalyptus na cubagem rigorosa para estimativa de modelos volumétricos. Rev. Bras. Biom. 2015, 33, 91–103. [Google Scholar]
- Castro, R.V.O.; Araújo, R.A.A.; Leite, H.G.; Neves, A.F.; Castro, M.; Silva, A.; Pereira, R.S.; Leal, F.A. Modelagem do crescimento e da produção de povoamentos de Eucalyptus em nível de distribuição diamétrica utilizando índice de local. Rev. Árvore 2016, 40, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Verão, D.S.; Bleich, M.E.; Martins, N.P.; Bassotto, J.M.; Mortat, A.F.; Santos, A.F.A. Concentração de nutrientes em Eucalyptus urograndis (Eucalyptus grandis W. Hill ex Maiden × Eucalyptus urophylla S. T Blake) com sete anos de idade na borda sul da Amazônia. Biodiversidade 2016, 15, 35–44. [Google Scholar]
- Barros, F.N.; Neves, J.C.L.; Novais, R.F. Nutrição e adubação mineral do eucalipto. In Eucaliptocultura No Brasil: Silvicultura, Manejo e Ambiência, 1st ed.; Vale, A.B., Machado, C.C., Pires, J.M.M., Vilar, M.B., Costa, C.B., Nacif, P.A., Eds.; SIF: Piracicaba, Brazil, 2014; pp. 187–208. [Google Scholar]
- Viera, M.; Schumacher, M.V.; Bonacina, D.M.; Ramos, L.O.O.; Rodrígues-Soalleiro, R. Biomass and nutrient allocation to aboveground components in fertilized Eucalyptus saligna and E. urograndis plantations. New Forests 2017, 48, 445–462. [Google Scholar] [CrossRef]
- Lattimore, B.; Smith, C.T.; Titus, B.D.; Stupak, I.; Ednell, G. Environmental factors in woodfuel production: Opportunities, risks, and criteria and indicators for sustainable practices. Biomass Bioenergy 2009, 33, 1321–1342. [Google Scholar] [CrossRef]
- Viera, M.; Schumacher, M.V.; Trüby, P.; Araújo, E.F. Implicações nutricionais com base em diferentes intensidades de colheita da biomassa de Eucalyptus urophylla × Eucalyptus globulus. Cienc. Rural 2015, 45, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.C.; Graley, A.M. Gains and losses in soil nutrients associated with harvesting and burning eucalypt rainforest. Plant. Soil 1983, 74, 437–450. [Google Scholar] [CrossRef]
- Mareschal, L.; Laclau, J.P.; Nzila, D.D.; Versini, A.; Koutika, L.S.; Mazoumbou, J.C.; Deleporte, P.; Bouillet, J.P.; Ranger, J. Nutrient leaching and deep drainage under Eucalyptus plantations managed in short rotations after afforestation of an African savanna: Two 7-year time series. Ecol. Manag. 2013, 307, 242–254. [Google Scholar] [CrossRef]
- Laclau, J.P.; Ranger, J.; Gonçalves, J.L.M.; Maquère, V.; Krusche, A.V.; M’Bou, A.T.; Nouvellon, Y.; Saint-André, L.; Bouillet, J.P.; Piccolo, M.C.; et al. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations: Main features shown by intensive monitoring in Congo and Brazil. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Valadares, S.V.; Neves, J.C.L.; Leite, H.G.; Barros, N.F.; Cropper Jr, W.P.; Gerber, S. Predicting phosphorus use efficiency and allocation in eucalypt plantations. Ecol. Manag. 2020, 460, 1–5. [Google Scholar] [CrossRef]
- Jones, H.E.; Madeira, M.; Herraez, L.; Dighton, J.; Fabiâo, A.; González-Rio, F.; Fernandez, M.; Gomez, C.; Tomé, M.; Feith, H.; et al. The effect of organic-matter management on the productivity of Eucalyptus globulus stands in Spain and Portugal: Tree growth and harvest residue decomposition in relation to site and treatment. Ecol. Manag. 1999, 122, 73–86. [Google Scholar] [CrossRef]
- Husch, B.; Miller, C.I.; Beers, T.W. Forest Mensuration, 2nd ed.; Jonh Willey & Sons: New York, NY, USA, 1972; p. 367. [Google Scholar]
- Silva, F.C. Análise química de tecido vegetal. In Manual de Análises Químicas de Solo, Plantas e Fertilizantes, 2nd ed.; Embrapa Informação Tecnológica: Brasília, Brazil, 2009; pp. 193–204. [Google Scholar]
- Schumacher, F.X.; Hall, F.S. Logarithmic expression of timber-tree volume. J. Agri. Res. 1933, 47, 719–734. [Google Scholar]
- Richards, F.J. A flexible growth function for empirical use. J. Exp. Bot. 1959, 10, 290–300. [Google Scholar] [CrossRef]
- Weibull, W. A statistical distribution function of wide applicability. J. Appl. Mech. 1951, 18, 293–299. [Google Scholar]
- Subasinghe, S.M.C.U.P. Construction of allometric relationships to predict growth parameters, stem biomass and carbon of Eucalyptus grandis growing in Sri Lanka. J. Trop. Sci. 2015, 5, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Assis, M.R.; Trugilho, P.F.; Rosado, S.C.S.; Protasio, T.P.; Goulart, S.L. Modelagem da biomassa e do estoque de carbono em plantas jovens de Eucalyptus. Science 2015, 43, 225–233. [Google Scholar]
- Ribeiro, M.D.S.B.; Jorge, L.A.B.; Mischan, M.M.; Santos, A.L.; Ballarin, A.W. Avaliação da produção de biomassa do fuste de um clone híbrido de eucalipto sob diferentes espaçamentos. Cienc. Florest 2017, 27, 31–45. [Google Scholar]
- Wernsdörfer, H.; Jonard, M.; Genet, A.; Legoutf, A.; Nys, C.; Saint-André, L.; Ponette, Q. Modelling of nutrient concentrations in roundwood based on diameterand tissue proportion: Evidence for an additional site-age effect in the case of Fagus sylvatica. Ecol. Manag. 2014, 330, 192–204. [Google Scholar] [CrossRef]
- Rance, S.J.; Mendham, D.S.; Cameron, D.M.; Grove, T.S. An evaluation of the conical approximation as a generic model for estimating stem volume, biomass and nutrient content in young Eucalyptus plantations. New Forests 2012, 43, 109–128. [Google Scholar] [CrossRef]
- Dick, G.; Schumacher, M.V.; Momolli, D.R.; Santos, J.C.; Ludvichak, A.A.; Guimarães, C.C.; Souza, H.P. Quantificação da biomassa e nutrientes em um povoamento de Eucalyptus dunnii Maiden estabelecido no Bioma Pampa. Ecol. E Nutr. Florest. 2016, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Miguel, E.P.; Rezende, A.V.; Pereira, R.S.; Azevedo, G.B.; Mota, F.C.M.; Souza, A.N.; Joaquim, M.S. Modeling and prediction of volume and aereal biomass of the tree vegetation in a Cerradão area of central Brazil. Interciencia 2017, 42, 21–27. [Google Scholar]
- Piñeiro, G.; Perelman, S.; Guerschman, J.; Paruelo, J.M. How to evaluate models: Observed vs. predicted or predicted vs. observed? Ecol. Model. 2008, 216, 316–322. [Google Scholar] [CrossRef]
- Ré, D.S.; Engel, V.L.; Ota, L.M.S.; Jorge, L.A.B. Equações alométricas em plantios mistos visando à restauração da floresta estacional semidecidual. Cerne 2015, 21, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Lanzarin, K.; Fleig, F.D.; Meyer, E.A.; Pereira, L.D.; Schröder, T.; Lisboa, G.S. Crescimento e biomassa de indivíduos jovens de Cabralea canjerana (Vell.) Mart. Biofix 2018, 3, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Fleming, R.A. The Weibull model and an ecological application: Describing the dynamics of foliage biomass on Scots pine. Ecol. Model. 2001, 138, 309–319. [Google Scholar] [CrossRef]
- Anthony, J.; Baccam, J.; Bier, I.; Gregg, E.; Halverson, L.; Mulcahy, R.; Okanla, E.; Osman, S.A.; Pancoast, A.R.; Schultz, K.C.; et al. Modeling mayfly nymph length distribution and population dynamics across a gradient of stream temperatures and stream types. SPORA J. Biomath. 2018, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Teleken, J.T.; Galvão, A.C.; Robazza, W.S. Use of modified Richards model to predict isothermal and non-isothermal microbial growth. Braz. J. Microbiol. 2018, 49, 614–620. [Google Scholar] [CrossRef]
- Campos, J.C.C.; Leite, H.G. Mensuração Florestal: Perguntas e Respostas, 5th ed.; UFV: Viçosa, Brazil, 2017; p. 636. [Google Scholar]
- Souza, A.F.; Junior, E.O.R.; Laura, V.A. Desenvolvimento inicial e eficiência de uso de água e nitrogênio por mudas de Calophyllum brasiliense, Eucalyptus urograndis, Tabebuia impetiginosa e Toona ciliata. Cienc. Florest 2018, 284, 1465–1477. [Google Scholar] [CrossRef] [Green Version]
- Viera, M.; Fernández, F.R.; Rodríguez-Soalleiro, R. Nutritional prescriptions for Eucalyptus plantations: Lessons learned from Spain. Forests 2016, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Resquin, F.; Navarro-Cerrillo, R.M.; Carrasco-Letelier, L.; Casnati, C.R.; Bentancor, L. Evaluation of the nutrient content in biomass of Eucalyptus species from short rotation plantations in Uruguay. Biomass Bioenergy 2020, 134, 1–14. [Google Scholar] [CrossRef]
- Valadão, M.B.X.; Carneiro, K.M.S.; Inkotte, J.; Ribeiro, F.P.; Miguel, E.P.; Gatto, A. Litterfall, litter layer and leaf decomposition in Eucalyptus stands on Cerrado soils. Sci. Forests 2019, 47, 256–264. [Google Scholar] [CrossRef]
- Carvalho, W.; Canilha, L.; Ferraz, A.; Milagres, A.M.F. Uma visão sobre a estrutura, composição e biodegradação da madeira. Quim. Nova 2009, 32, 2191–2195. [Google Scholar] [CrossRef] [Green Version]
- Viera, M.; Schumacher, M.V.; Araújo, E.F. Disponibilização de nutrientes via decomposição da serapilheira foliar em um plantio de Eucalyptus urophylla × Eucalyptus globulus. Floresta Ambient. 2015, 21, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Lima, R.P.; Fernandes, M.M.; Fernandes, M.R.M.; Matricardi, E.A.T. Aporte e Decomposição da Serapilheira na Caatinga no Sul do Piauí. Floresta Ambient. 2015, 22, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Bachega, L.R.; Bouillet, J.P.; Piccolo, M.C.; Saint-André, L.; Bouvete, J.M.; Nouvellon, Y.; Gonçalves, J.L.M.; Robin, A.; Laclau, J.P. Decomposition of Eucalyptus grandis and Acacia mangium leaves and fine roots in tropical conditions did not meet the Home Field Advantage hypothesis. Ecol. Manag. 2016, 359, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Vieira, M.; Schumacher, M.V.; Trüby, P.; Araújo, E.F. Biomassa e nutrientes em um povoamento de Eucalyptus urophylla × Eucalyptus globulus, em Eldorado do Sul-RS. Ecol. E Nutr. Florest. 2013, 1, 1–13. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Moller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier: Oxford, UK, 2012; Volume 1, pp. 135–189. [Google Scholar]
- Saidelles, F.L.F.; Caldeira, M.V.W.; Schumacher, M.V.; Balbinot, R.; Schirmer, W.N. Determinação do ponto de amostragem para a quantificação de macronutrientes em Acacia mearnsii de Wild. Floresta 2010, 40, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Téo, S.J.; Machado, S.A.; Figueiredo Filho, A.; Reissmann, C.B. Modelagem de micronutrientes para Mimosa scabrella na região metropolitana de Curitiba, estado do Paraná. Pesq. Flor. Bras. 2009, 59, 15–26. [Google Scholar] [CrossRef]
- Wink, C.; Reinert, D.J.; Tornquist, C.G.; Silva, I.R. Dinâmica do carbono e nitrogênio em plantações de eucalipto no Rio Grande do Sul. R. Bras. Ciênc. Solo 2015, 39, 1623–1632. [Google Scholar] [CrossRef] [Green Version]
- Cantarella, H. Nitrogênio. In Fertilidade do Solo, 1st ed.; Novais, R.F., Alvarez, V.H., Barros, N.F., Fontes, R.L.F., Cantarutti, R.B., Neves, J.C.L., Eds.; SBCS: Viçosa, Brazil, 2007; Volume 1, pp. 375–470. [Google Scholar]
- Barbosa, V.; Barreto-Garcia, P.; Gama-Rodrigues, E.; Paula, A. Biomassa, carbono e nitrogênio na serapilheira acumulada de florestas plantadas e nativa. Floresta Ambient. 2017, 24, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Barreto, P.A.B.; Gama-Rodrigues, E.F.; Gama-Rodrigues, A.C.; Barros, N.F.; Alves, B.J.; Fonseca, S. Mineralização de nitrogênio e carbono em solos sob plantações de eucalipto, em uma sequência de idades. R. Bras. Ciênc. Solo 2010, 34, 735–745. [Google Scholar] [CrossRef]
- Zanchi, G.; Belyazid, S.; Akselsson, C.; Yu, L. Modelling the effects of management intensification on multiple forest services: A Swedish case study. Ecol. Model. 2014, 284, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Zanchi, G.; Akselsson, C.; Wallander, H.; Belyazid, S. Modeling the forest phosphorus nutrition in a southwestern Swedish forest site. Ecol. Model. 2018, 369, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Gielen, S.; Batllea, J.V.; Vincke, C.; Van Hees, M.; Vandenhovea, H. Concentrations and distributions of Al, Ca, Cl, K, Mg and Mn in a Scots pine forest in Belgium. Ecol. Model. 2016, 324, 1–10. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Volume | Unit | Biomass | Unit |
|---|---|---|---|---|
| Mean | 328.88 | m3 ha−1 | 181.405 | Mg ha−1 |
| Variance | 3385.654 | (m3 ha−1)2 | 1061.586 | (Mg ha−1)2 |
| Standard deviation | 58.186 | m3 ha−1 | 32.582 | Mg ha−1 |
| Coefficient of variation (%) | 17.692 | % | 17.961 | % |
| Variance of the mean | 74.351 | (m3 ha−1)2 | 23.310 | (Mg ha−1)2 |
| Standard error of mean | 8.623 | m3 ha−1 | 4.828 | Mg ha−1 |
| Absolute sample error | 17.441 | m3 ha−1 | 9.766 | Mg ha−1 |
| Sample relative error | 5.303 | % | 5.384 | % |
| Variables | G I | H II | DBH III | Biomass IV | N V | P VI | K VII | Ca VIII | Mg IX |
|---|---|---|---|---|---|---|---|---|---|
| G I | 1.00 | 0.45 ** | 0.47 ** | 0.97 ** | 0.26 ns | 0.84 ** | 0.93 ** | 0.90 ** | 0.88 ** |
| H II | 0.45 ** | 1.00 | 0.82 ** | 0.59 ** | −0.55 ** | 0.20 ns | 0.34 * | 0.35 * | 0.28 ns |
| DBH III | 0.47 ** | 0.82 ** | 1.00 | 0.61 ** | −0.61 ** | 0.01 ns | 0.20 ns | 0.19 ns | 0.10 ns |
| Biomass IV | 0.97 ** | 0.59 ** | 0.61 ** | 1.00 | 0.14 ns | 0.73 ** | 0.86 ** | 0.82 ** | 0.79 ** |
| N V | 0.26 ns | −0.55 ** | −0.61 ** | 0.14 ns | 1.00 | 0.49 ** | 0.40 ** | 0.35 * | 0.43 ** |
| P VI | 0.84 ** | 0.20 ns | 0.01 ns | 0.73 ** | 0.49 ** | 1.00 | 0.98 ** | 0.98 ** | 0.99 ** |
| K VII | 0.93 ** | 0.34 * | 0.20 ns | 0.86 ** | 0.40 ** | 0.98 ** | 1.00 | 0.99 ** | 0.99 ** |
| Ca VIII | 0.90 ** | 0.35 * | 0.19 ns | 0.82 ** | 0.35 * | 0.98 ** | 0.99 ** | 1.00 | 0.99 ** |
| Mg IX | 0.88 ** | 0.28 ns | 0.10 ns | 0.79 ** | 0.43 ** | 0.99 ** | 0.99 ** | 0.99 ** | 1.00 |
| Model | Equation |
|---|---|
| Richards (Richards 1959) | * |
| Weibull (Weibull and Sweden 1951) | ** |
| Schumacher-Hall (Schumacher and Hall 1933) | *** |
| Compartments | Mean Biomass | N | P | K | Ca | Mg |
|---|---|---|---|---|---|---|
| (Mg) | (%) | |||||
| Leaves | 0.011 (4.36%) | 1.375 | 0.077 | 0.433 | 0.573 | 0.363 |
| Branches | 0.010 (4.26%) | 0.460 | 0.039 | 0.318 | 0.322 | 0.169 |
| Bark | 0.029 (11.99%) | 0.255 | 0.020 | 0.260 | 1.198 | 0.984 |
| Crown wood | 0.014 (5.58%) | 0.091 | 0.008 | 0.103 | 0.099 | 0.019 |
| Stem wood | 0.177 (72.15%) | 0.084 | 0.002 | 0.039 | 0.056 | 0.005 |
| Roots | 0.004 (1.66%) | 0.292 | 0.024 | 0.085 | 0.307 | 0.083 |
| Total | 0.246 (100%) | 2.557 | 0.170 | 1.238 | 2.555 | 1.623 |
| Model | α * | β * | γ * | δ ** | Syx *** (Mg ha−1) | Syx **** (%) | r ***** |
|---|---|---|---|---|---|---|---|
| Richards | 304.492 | −2.984 | 0.000 | 0.034 | 4.966 | 2.701 | 0.991 |
| Weibull | 335.095 | 271.007 | 0.000 | 1.099 | 4.930 | 2.682 | 0.991 |
| Schumacher–Hall | 1.282 | 1.013 | 0.504 | - | 4.934 | 2.684 | 0.991 |
| Model | α * | β * | γ * | δ ** | Syx *** (Mg ha−1) | Syx **** (%) | r ***** |
|---|---|---|---|---|---|---|---|
| Richards | 4.600 | −3.213 | −0.011 | 2.538 | 0.547 | 17.581 | 0.661 |
| Weibull | 4.157 | 6.601 | 12,476.673 | −1.502 | 0.547 | 17.578 | 0.661 |
| Schumacher–Hall | 256.475 | −1.288 | −0.283 | - | 0.546 | 17.525 | 0.648 |
| Model | α * | β * | γ * | δ ** | Syx *** (Mg ha−1) | Syx **** (%) | r ***** |
|---|---|---|---|---|---|---|---|
| Richards | 0.650 | 4.142 | 0.054 | 1.927 | 0.014 | 8.386 | 0.840 |
| Weibull | 1.407 | 1.319 | 0.000 | 1.683 | 0.014 | 8.388 | 0.840 |
| Schumacher-Hall | 0.048 | 0.884. | −0.562 | - | 0.012 | 7.214 | 0.880 |
| Model | α * | β * | γ * | δ ** | Syx *** (Mg ha−1) | Syx **** (%) | r ***** |
|---|---|---|---|---|---|---|---|
| Richards | 2.192 | 0.254 | 0.032 | 0.370 | 0.042 | 5.790 | 0.930 |
| Weibull | 3.664 | 3.397 | 0.001 | 1.444 | 0.042 | 5.789 | 0.930 |
| Schumacher-Hall | 0.085 | 0.906 | −0.289 | - | 0.038 | 5.291 | 0.939 |
| Model | α * | β * | γ * | δ ** | Syx *** (Mg ha−1) | Syx **** (%) | r ***** |
|---|---|---|---|---|---|---|---|
| Richards | 7.439 | −0.557 | 0.030 | 0.199 | 0.171 | 6.827 | 0.906 |
| Weibull | 21.904 | 21.227 | 0.002 | 1.206 | 0.171 | 6.819 | 0.906 |
| Schumacher–Hall | 0.269 | 0.895 | −0.248 | - | 0.162 | 6.445 | 0.913 |
| Model | α * | β * | γ * | δ ** | Syx *** (Mg ha−1) | Syx **** (%) | r ***** |
|---|---|---|---|---|---|---|---|
| Richards | 2.296 | 0.463 | 0.032 | 0.444 | 0.055 | 7.202 | 0.887 |
| Weibull | 4.388 | 4.103 | 0.001 | 1.360 | 0.055 | 7.200 | 0.887 |
| Schumacher–Hall | 0.127 | 0.885 | −0.381 | - | 0.050 | 6.506 | 0.905 |
| Variables (Mg ha−1) | Pcalculated * | Aggregated Difference (%) |
|---|---|---|
| Biomass | 0.89 | 0,43 |
| Nitrogen | 0.77 | −0.57 |
| Phosphorus | 0.14 | −4.09 |
| Potassium | 0.08 | −3.28 |
| Calcium | 0.09 | −3.78 |
| Magnesium | 0.70 | −3.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valadão, M.B.X.; Carneiro, K.M.S.; Ribeiro, F.P.; Inkotte, J.; Rodrigues, M.I.; Mendes, T.R.S.; Vieira, D.A.; Matias, R.A.M.; Lima, M.B.O.; Miguel, E.P.; et al. Modeling Biomass and Nutrients in a Eucalyptus Stand in the Cerrado. Forests 2020, 11, 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/f11101097
Valadão MBX, Carneiro KMS, Ribeiro FP, Inkotte J, Rodrigues MI, Mendes TRS, Vieira DA, Matias RAM, Lima MBO, Miguel EP, et al. Modeling Biomass and Nutrients in a Eucalyptus Stand in the Cerrado. Forests. 2020; 11(10):1097. https://0-doi-org.brum.beds.ac.uk/10.3390/f11101097
Chicago/Turabian StyleValadão, Marco B. X., Karla M. S. Carneiro, Fabiana P. Ribeiro, Jonas Inkotte, Maísa I. Rodrigues, Thallita R. S. Mendes, Daniel A. Vieira, Renan A. M. Matias, Mirella B. O. Lima, Eder P. Miguel, and et al. 2020. "Modeling Biomass and Nutrients in a Eucalyptus Stand in the Cerrado" Forests 11, no. 10: 1097. https://0-doi-org.brum.beds.ac.uk/10.3390/f11101097