Detecting Growth Phase Shifts Based on Leaf Trait Variation of a Canopy Dipterocarp Tree Species (Parashorea chinensis)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
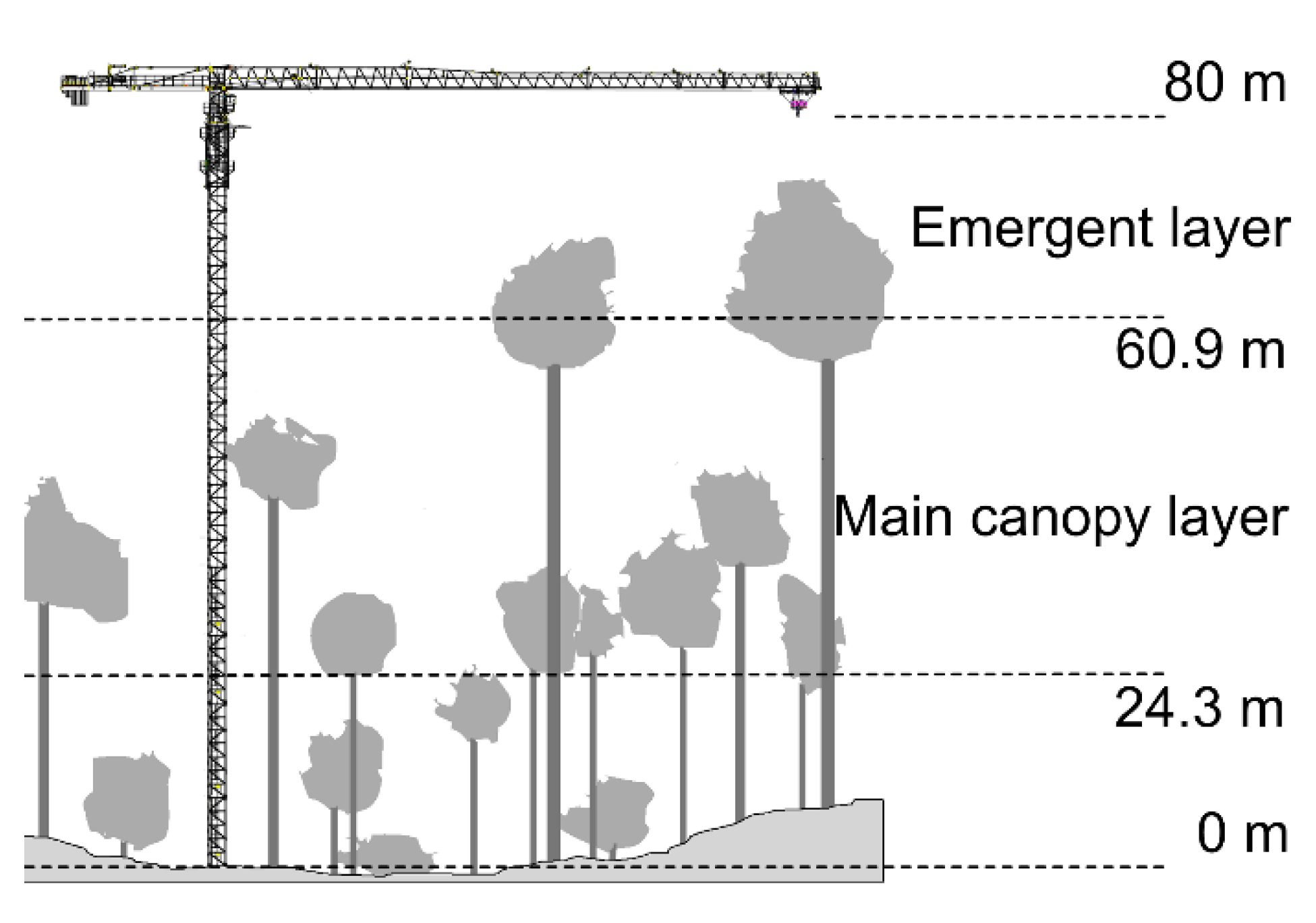
2.1. Study Area
2.2. Light Conditions
2.3. Leaf Dark Respiration Rate (Rd)
2.4. Light-Response Curve
2.5. Leaf Mass per Area (LMA), Leaf Nitrogen, and Carbon
2.6. Statistical Analysis
3. Results
3.1. Tree Size and Light Availability
3.2. Growth Phases
3.3. Leaf Morphological Trait Transition Point of P. chinensis
3.4. Physiological Trait Transition Point of P. chinensis
4. Discussions
4.1. Relationships between Variations in Leaf Traits
4.2. Growth Phases and Adaptive Strategies of P. chinensis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dayrell, R.L.C.; Arruda, A.J.; Pierce, S.; Negreiros, D.; Meyer, P.B.; Lambers, H.; Silveira, F.A.O. Ontogenetic shifts in plant ecological strategies. Funct. Ecol. 2018, 32, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Sack, L.; Grubb, P.J. Why do species of woody seedlings change rank in relative growth rate between low and high irradiance? Funct. Ecol. 2001, 15, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F. Light heterogeneity and plants: From ecophysiology to species coexistence and biodiversity. In Progress in Botany; Esser, K., Lüttge, U., Beyschlag, W., Hellwig, F., Eds.; Springer: Berlin, Germany, 2003; pp. 439–471. [Google Scholar]
- Coopman, R.E.; Briceno, V.F.; Corcuera, L.J.; Reyes-Diaz, M.; Alvarez, D.; Saez, K.; Garcia-Plazaola, J.I.; Alberdi, M.; Bravo, L.A. Tree size and light availability increase photochemical instead of non-photochemical capacities of Nothofagus nitida trees growing in an evergreen temperate rain forest. Tree Physiol. 2011, 31, 1128–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavaleri, M.A.; Oberbauer, S.F.; Clark, D.B.; Clark, D.A.; Ryan, M.G. Height is more important than light in determining leaf morphology in a tropical forest. Ecology 2010, 91, 1730–1739. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Grubb, P.J. The maintenance of species-richness in plant communities: The importance of the regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Pierce, S.; Negreiros, D.; Cerabolini, B.E.L.; Kattge, J.; Díaz, S.; Kleyer, M.; Shipley, B.; Wright, S.J.; Soudzilovskaia, N.A.; Onipchenko, V.G.; et al. A global method for calculating plant CSR ecological strategies applied across biomes world-wide. Funct. Ecol. 2017, 31, 444–457. [Google Scholar] [CrossRef]
- Lloyd, J.; Patiño, S.; Paiva, R.Q.; Nardoto, G.B.; Quesada, C.A.; Santos, A.J.B.; Baker, T.R.; Brand, W.A.; Hilke, I.; Gielmann, H.; et al. Optimisation of photosynthetic carbon gain and within-canopy gradients of associated foliar traits for Amazon forest trees. Biogeosciences 2010, 7, 1833–1859. [Google Scholar] [CrossRef] [Green Version]
- Pierce, S.; Brusa, G.; Vagge, I.; Cerabolini, B.E.L. Allocating CSR plant functional types: The use of leaf economics and size traits to classify woody and herbaceous vascular plants. Funct. Ecol. 2013, 27, 1002–1010. [Google Scholar] [CrossRef] [Green Version]
- Pierce, S.; Brusa, G.; Sartori, M.; Cerabolini, B.E.L. Combined use of leaf size and economics traits allows direct comparison of hydrophyte and terrestrial herbaceous adaptive strategies. Ann. Bot. 2012, 109, 1047–1053. [Google Scholar] [CrossRef] [Green Version]
- Pacala, S.W.; Canham, C.D.; Saponara, J.; Silander, J.A.; Kobe, R.K.; Ribbens, E. Forest models defined by field measurements: Estimation, error analysis and dynamics. Ecol. Monogr. 1996, 66, 1–43. [Google Scholar] [CrossRef]
- Kenzo, T.; Inoue, Y.; Yoshimura, M.; Yamashita, M.; Tanaka-Oda, A.; Ichie, T. Height-related changes in leaf photosynthetic traits in diverse Bornean tropical rain forest trees. Oecologia 2015, 177, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.W.; Sillett, S.C.; Jennings, G.M.; Davis, S.D. The limits to tree height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef]
- Thomas, S.C. Photosynthetic capacity peaks at intermediate size in temperate deciduous trees. Tree Physiol. 2010, 30, 555–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bond, B.J. Age-related changes in photosynthesis of woody plants. Trends Plant Sci. 2000, 5, 349–353. [Google Scholar] [CrossRef]
- Niinemets, U. Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris. Tree Physiol. 2002, 22, 515–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.C.; Ickes, K. Ontogenetic changes in leaf size in Malaysian rain forest trees. Biotropica 1995, 27, 427–434. [Google Scholar] [CrossRef]
- Cao, M.; Zhang, J. Tree species diversity of tropical forest vegetation in Xishuangbanna, SW China. Biodivers. Conserv. 1997, 6, 995–1006. [Google Scholar] [CrossRef]
- Cao, M.; Zhu, H.; Wang, H.; Lan, G.; Hu, Y.; Deng, S.Z.X.; Cui, J. Xishuangbanna Tropical Seasonal Rainforest Dynamics Plot: Tree Distribution Maps, Diameter Tables and Species Documentation; Yunnan Science and Technology Press: Kunming, China, 2008. [Google Scholar]
- Van der Velden, N.; Ferry Slik, J.W.; Hu, Y.H.; Lan, G.; Lin, L.; Deng, X.; Poorter, L. Monodominance of Parashorea chinensis on fertile soils in a Chinese tropical rain forest. J. Trop. Ecol. 2014, 30, 311–322. [Google Scholar] [CrossRef]
- Ying, S.; Shuai, J. Study on fruiting behavior, seedling establishment and population age classes of Parashorea chinensis. Acta Bot. Yunnanica 1990, 12, 415–420. [Google Scholar]
- Tang, J.W.; Shi, J.P.; Zhang, G.M.; Bai, K.J. Density, structure and biomass of Papashorea chinensis population in different patches in Xishuangbanna, SW. J. Plant Ecol. 2018, 32, 40–54. [Google Scholar]
- Yan, X.F.; Cao, M. Effects of light intensity on seed germination and seedling early growth of Shorea wantianshuea. Chin. J. Appl. Ecol. 2007, 18, 23–29. [Google Scholar]
- He, C.X.; Li, J.Y.; Zhou, P.; Guo, M.; Zheng, Q.S. Changes of leaf morphological, anatomical structure and carbon isotope ratio with the height of the Wangtian tree (Parashorea chinensis) in Xishuangbanna, China. J. Integr. Plant Biol. 2008, 50, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, B.M.J.; Herz, H.M. Evaluation of different methods to estimate understorey light conditions in tropical forests. J. Trop. Ecol. 2001, 17, 207–224. [Google Scholar] [CrossRef] [Green Version]
- Atkin, O.K.; Evans, J.R.; Siebke, K. Relationship between the inhibition of leaf respiration by light and enhancement of leaf dark respiration following light treatment. Funct. Plant Biol. 1998, 25, 437–443. [Google Scholar] [CrossRef]
- Florez-Sarasa, I.; Araújo, W.L.; Wallström, S.V.; Rasmusson, A.G.; Fernie, A.R.; Ribas-Carbo, M. Light-responsive metabolite and transcript levels are maintained following a dark-adaptation period in leaves of Arabidopsis Thaliana. New Phytol. 2012, 195, 136–148. [Google Scholar] [CrossRef]
- Meng, L.Z.; Zhang, J.L.; Cao, K.F.; Xu, Z.F. Diurnal changes of photosynthetic characteristics and chlorophyll fluorescence in canopy leaves of four diptocarp species under ex-situ conservation. Acta Phytoecol. Sin. 2005, 29, 976–984. [Google Scholar] [CrossRef] [Green Version]
- Woodruff, D.R.; Meinzer, F.C.; Lachenbruch, B.; Johnson, D.M. Coordination of leaf structure and gas exchange along a height gradient in a tall conifer. Tree Physiol. 2009, 29, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.L.; Meng, L.Z.; Cao, K.F. Sustained diurnal photosynthetic depression in uppermost-canopy leaves of four dipterocarp species in the rainy and dry seasons: Does photorespiration play a role in photoprotection? Tree Physiol. 2009, 29, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Santiago, L.S.; Mulkey, S.S. A test of gas exchange measurements on excised canopy branches of ten tropical tree species. Photosynthetica 2003, 41, 343–347. [Google Scholar] [CrossRef]
- Bassman, J.H.; Zwier, J.C. Gas exchange characteristics of Populus trichocarpa, Populus deltoides and Populus trichocarpa x P. deltoides clones. Tree Physiol. 1991, 8, 145–159. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Cornelissen, J.H.C.; Falster, D.S.; Garnier, E.; Hikosaka, K.; Lamont, B.B.; Lee, W.; Oleksyn, J.; Osada, N.; et al. Assessing the generality of global leaf trait relationships. New Phytol. 2005, 166, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Coble, A.P.; Cavaleri, M.A. Light acclimation optimizes leaf functional traits despite height-related constraints in a canopy shading experiment. Oecologia 2015, 177, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- De’ath, G. Multivariate regression trees: A new technique for modeling species–environment relationships. Ecology 2002, 83, 1105–1117. [Google Scholar] [CrossRef]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.I. Classification and Regression Trees; Wadsworth International Group: Belmont, CA, USA, 1984. [Google Scholar]
- Ishii, H.T.; Jennings, G.M.; Sillett, S.C.; Koch, G.W. Hydrostatic constraints on morphological exploitation of light in tall Sequoia sempervirens trees. Oecologia 2008, 156, 751–763. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Givnish, T.J. Adaptation to sun and shade: A whole-plant perspective. Funct. Plant Biol. 1988, 15, 63–92. [Google Scholar] [CrossRef] [Green Version]
- Craine, J.M.; Reich, P.B. Leaf-level light compensation points in shade-tolerant woody seedlings. New Phytol. 2005, 166, 710–713. [Google Scholar] [CrossRef]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Amthor, J.S. Respiration and Crop Productivity; Springer: New York, NY, USA, 1989. [Google Scholar]
- Kenzo, T.; Yoneda, R.; Sano, M.; Araki, M.; Shimizu, A.; Tanaka-Oda, A.; Chann, S. Variations in leaf photosynthetic and morphological traits with tree height in various tree species in a Cambodian tropical dry evergreen forest. JARQ 2012, 46, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Oguchi, R.; Hikosaka, K.; Hirose, T. Leaf anatomy as a constraint for photosynthetic acclimation: Differential responses in leaf anatomy to increasing growth irradiance among three deciduous trees. Plant Cell Environ. 2005, 28, 916–927. [Google Scholar] [CrossRef]
- Baltzer, J.L.; Thomas, S.C. Determinants of whole-plant light requirements in Bornean rain forest tree saplings. J. Ecol. 2007, 95, 1208–1221. [Google Scholar] [CrossRef]
- Yan, X.F.; Cao, M. The endangered causes and protective strategies for Shorea wantianshuea, a tropical rain forest tree species in Xishuangbanna. J. Fujian Sci. Technol. 2008, 35, 187–191. [Google Scholar]
- Field, C. Allocating leaf nitrogen for the maximization of carbon gain: Leaf age as a control on the allocation program. Oecologia 1983, 56, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Kogami, H.; Hanba, Y.T.; Kibe, T.; Terashima, I.; Masuzawa, T. CO2 transfer conductance, leaf structure and carbon isotope composition of Polygonum cuspidatum leaves from low and high altitudes. Plant Cell Environ. 2001, 24, 529–538. [Google Scholar] [CrossRef]
- Ishida, A.; Yazaki, K.; Hoe, A.L. Ontogenetic transition of leaf physiology and anatomy from seedlings to mature trees of a rain forest pioneer tree, Macaranga gigantea. Tree Physiol. 2005, 25, 513–522. [Google Scholar] [CrossRef]
- Yan, X.F.; Cao, M. Seeding growth and survival of the endangered tree species Shorea wantianshuea after a mast-fruiting event. J. Plant Ecol. 2008, 32, 55–64. [Google Scholar]
- Clark, D.A.; Clark, D.B. Life history diversity of canopy and emergent trees in a neotropical rain forest. Ecol. Monogr. 1992, 62, 315–344. [Google Scholar] [CrossRef]
- Connell, J.H.; Green, P.T. Seedling dynamics over thirty-two years in a tropical rain forest tree. Ecology 2000, 81, 568–584. [Google Scholar] [CrossRef]
- Still, M.J. Rates of mortality and growth in three groups of dipterocarp seedlings in Sabah, Malaysia. In The Ecology of Tropical Forest Tree Seedlings (Man & The Biosphere Series); Swaine, M.D., Ed.; UNESCO: Paris, France, 1996; Volume 18, pp. 315–332. [Google Scholar]
- Su, W.H. Preliminary study on the dynamics of Pometia tomentosa population in the tropical seasonal rain forest of Xishuangbanna. Acta Bot. Yunnanica 1997, 9, 92–96. [Google Scholar]
- Silvertown, J. Introduction to Plant Population Ecology; Longman Scientific & Technical: New York, NY, USA, 1987. [Google Scholar]
- Kitajima, K. Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees. Oecologia 1994, 98, 419–428. [Google Scholar] [CrossRef]
- Kitahashi, Y.; Ichie, T.; Maruyama, Y.; Kenzo, T.; Kitaoka, S.; Matsuki, S.; Chong, L.; Nakashizuka, T.; Koike, T. Photosynthetic water use efficiency in tree crowns of Shorea beccariana and Dryobalanops aromatica in a tropical rain forest in Sarawak, East Malaysia. Photosynthetica 2008, 46, 151–155. [Google Scholar] [CrossRef]
- Kenzo, T.; Ichie, T.; Yoneda, R.; Kitahashi, Y.; Watanabe, Y.; Ninomiya, I.; Koike, T. Interspecific variation of photosynthesis and leaf characteristics in canopy trees of five species of Dipterocarpaceae in a tropical rain forest. Tree Physiol. 2004, 24, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Kenzo, T.; Ichie, T.; Watanabe, Y.; Yoneda, R.; Ninomiya, I.; Koike, T. Changes in photosynthesis and leaf characteristics with tree height in five dipterocarp species in a tropical rain forest. Tree Physiol. 2006, 26, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Martin, A.R.; Thomas, S.C. Size-dependent changes in leaf and wood chemical traits in two Caribbean rainforest trees. Tree Physiol. 2013, 33, 1338–1353. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.; Appanah, S. On the statistical analysis of reproductive size thresholds in dipterocarp forests. J. Trop. For. Sci. 1995, 27, 412–418. [Google Scholar]
- Patankar, R.; Thomas, S.C.; Smith, S.M. A gall-inducing arthropod drives declines in canopy tree photosynthesis. Oecologia 2011, 167, 701–709. [Google Scholar] [CrossRef]
- Hossain, S.M.Y.; Caspersen, J.P. In-situ measurement of twig dieback and regrowth in mature Acer saccharum trees. For. Ecol. Manag. 2012, 270, 183–188. [Google Scholar] [CrossRef]
- Sillett, S.C.; Van Pelt, R.; Koch, G.W.; Ambrose, A.R.; Carroll, A.L.; Antoine, M.E.; Mifsud, B.M. Increasing wood production through old age in tall trees. For. Ecol. Manag. 2010, 259, 976–994. [Google Scholar] [CrossRef]
- Parkhurst, D.F.; Loucks, O.L. Optimal leaf size in relation to environment. J. Ecol. 1972, 60, 505–537. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Phase | Transmittance of Diffuse Light | Tree Height | DBH | Strategy | Representative Traits (Variation Tendency) |
|---|---|---|---|---|---|
| Emergent | 100% | 60.9 m | ≥96.8 cm | – | Leaf area and Amax_area (decline) |
| Canopy | 100% | 24.3–60.9 m | 26.7–96.8 cm | C | Leaf area and Amax_area (highest) |
| Growth release | 18.33–100% | 14.9–24.3 m | 14.8–26.7 cm | _ | All traits (rapid change) |
| Suppressed | 3.94–18.33% | 8.3–14.9 m | 6–14.8 cm | S | LCP (low) PNUE and Amax_mass (highest) |
| Vulnerable | 0–3.94% | 0–8.3 m | <6 cm | _ | LCP (low) PNUE and Amax_mass (low) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; Deng, X.; Dong, J.; Zhang, W.; Hu, T.; Nakamura, A.; Song, X.; Fu, P.; Cao, M. Detecting Growth Phase Shifts Based on Leaf Trait Variation of a Canopy Dipterocarp Tree Species (Parashorea chinensis). Forests 2020, 11, 1145. https://0-doi-org.brum.beds.ac.uk/10.3390/f11111145
Deng Y, Deng X, Dong J, Zhang W, Hu T, Nakamura A, Song X, Fu P, Cao M. Detecting Growth Phase Shifts Based on Leaf Trait Variation of a Canopy Dipterocarp Tree Species (Parashorea chinensis). Forests. 2020; 11(11):1145. https://0-doi-org.brum.beds.ac.uk/10.3390/f11111145
Chicago/Turabian StyleDeng, Yun, Xiaobao Deng, Jinlong Dong, Wenfu Zhang, Tao Hu, Akihiro Nakamura, Xiaoyang Song, Peili Fu, and Min Cao. 2020. "Detecting Growth Phase Shifts Based on Leaf Trait Variation of a Canopy Dipterocarp Tree Species (Parashorea chinensis)" Forests 11, no. 11: 1145. https://0-doi-org.brum.beds.ac.uk/10.3390/f11111145