Carbon and Nutrient Inputs by Litterfall in Evergreen and Deciduous Forests in Korea

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Stand Descriptions
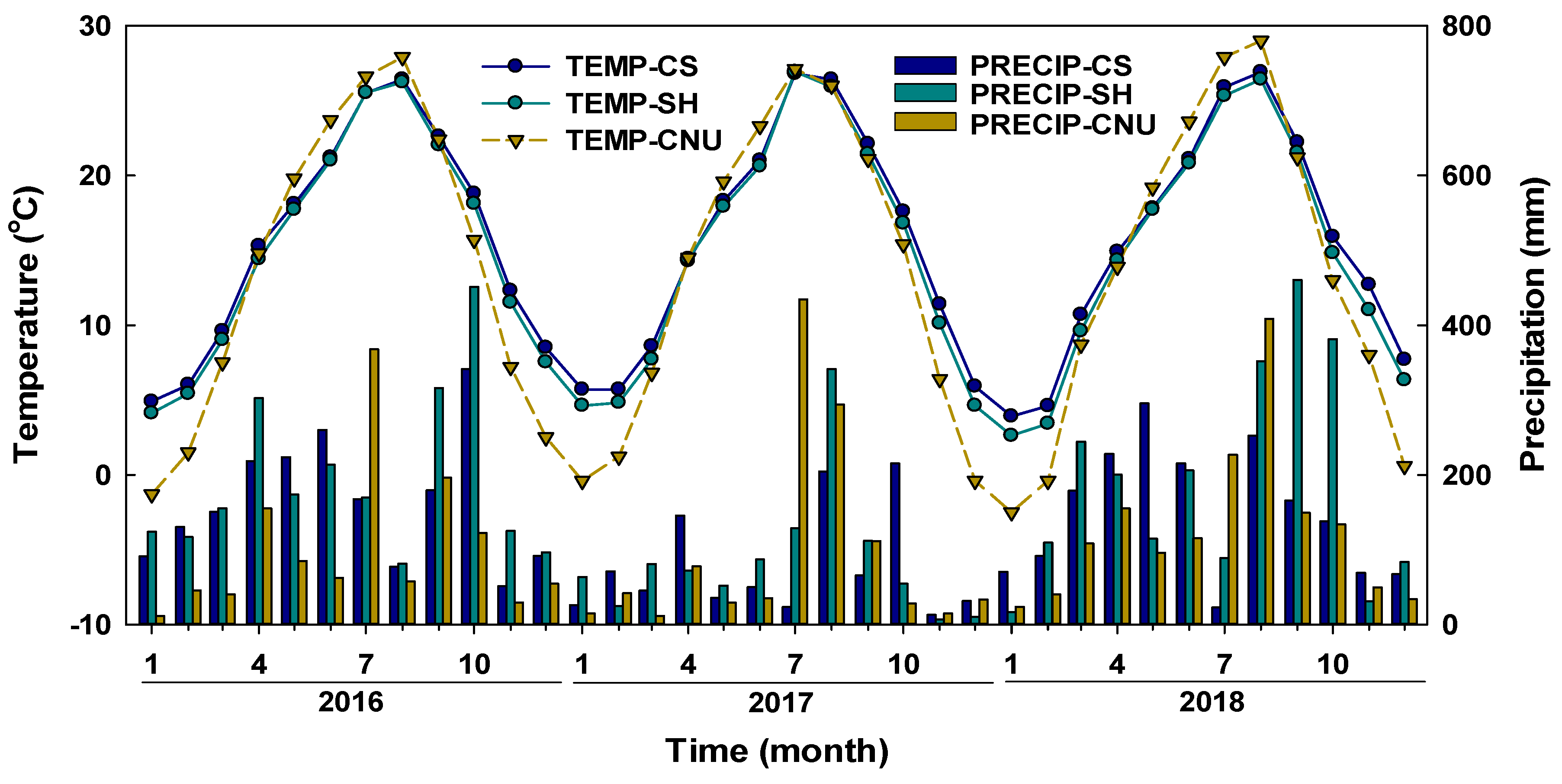
2.2. Climate Conditions
2.3. Litterfall Collection
2.4. Nutrient Analysis
2.5. Statistical Analysis
3. Results
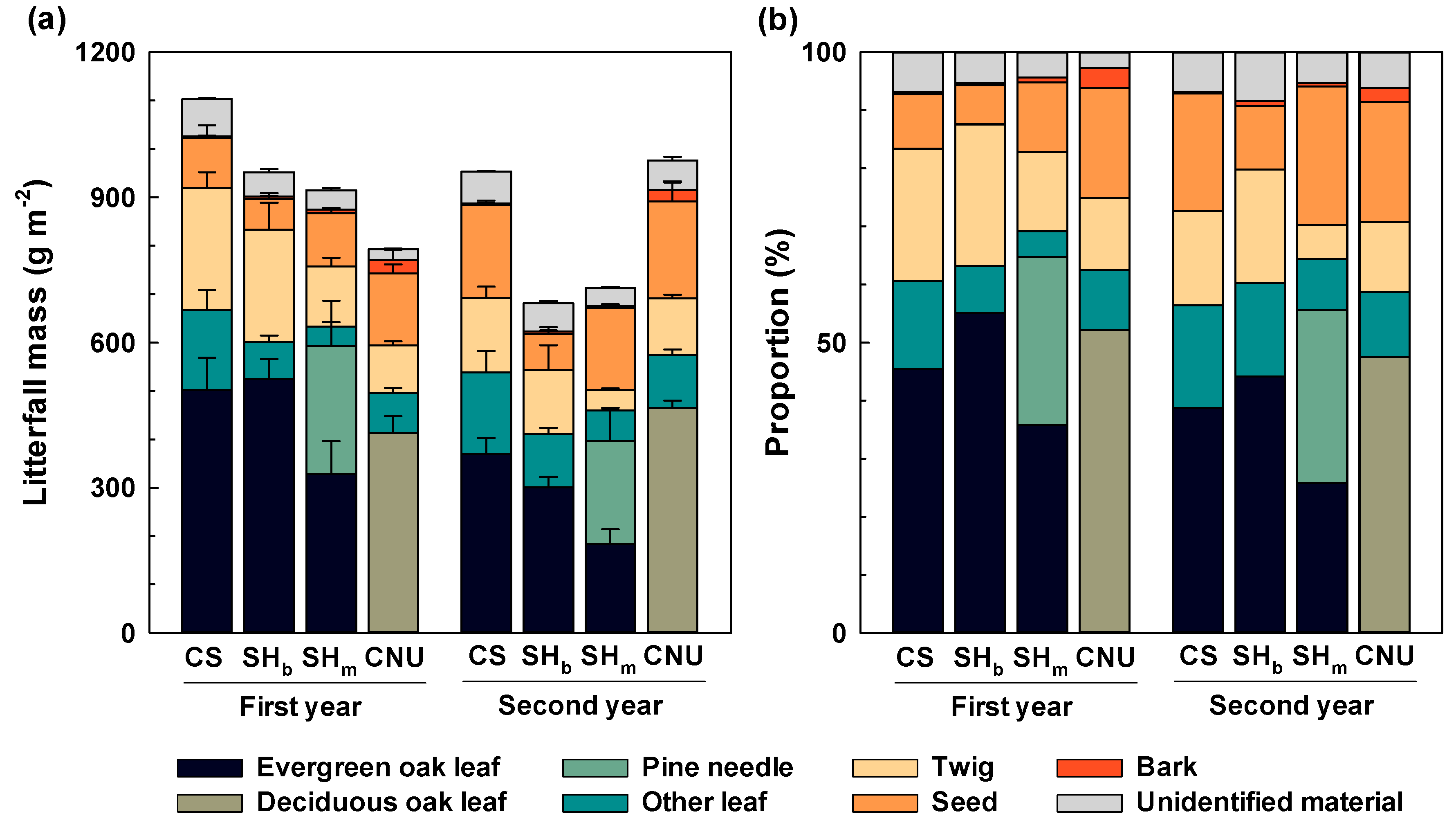
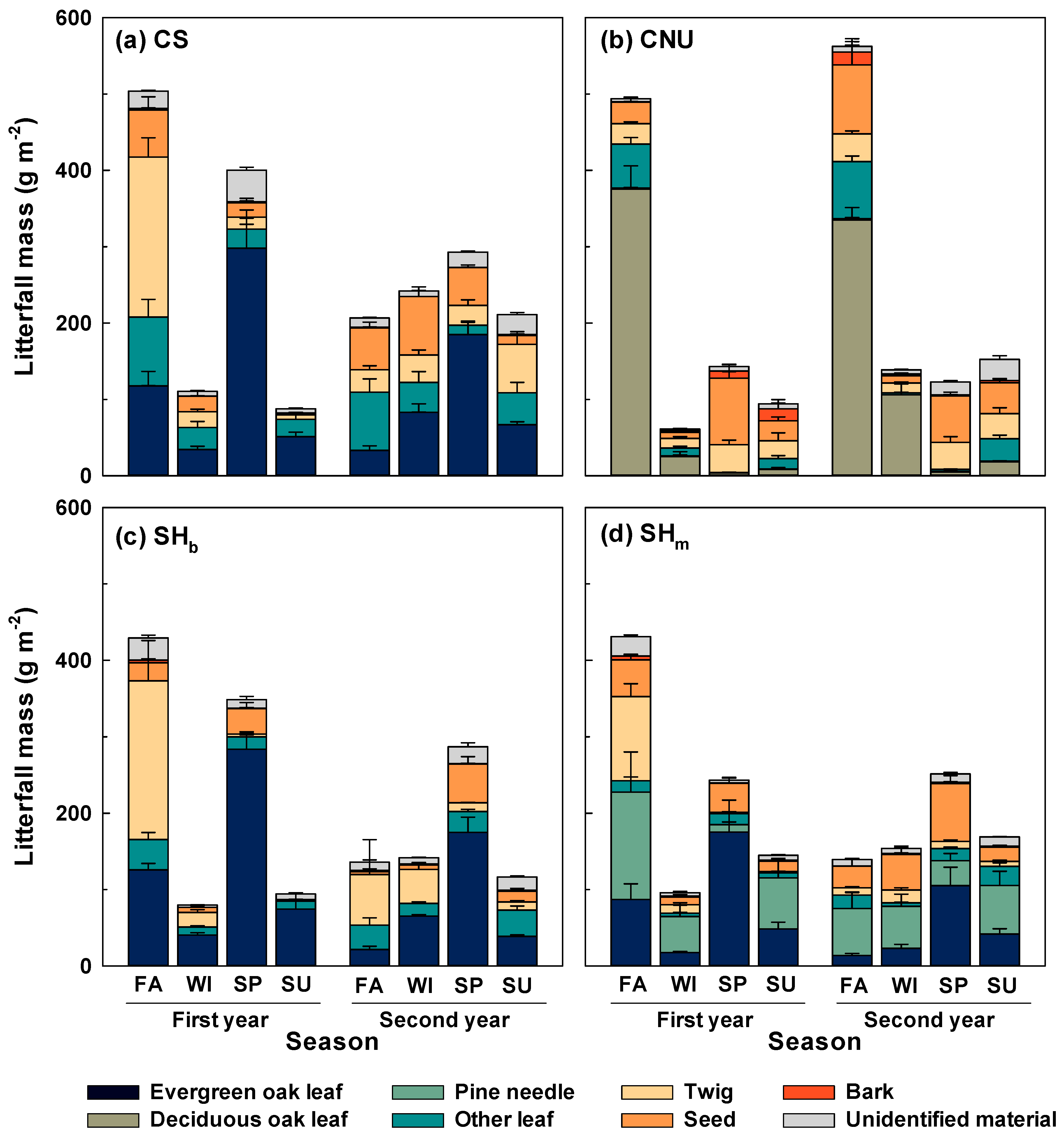
3.1. Litterfall Variation
3.2. Carbon and Nutrient Concentrations of Litterfall
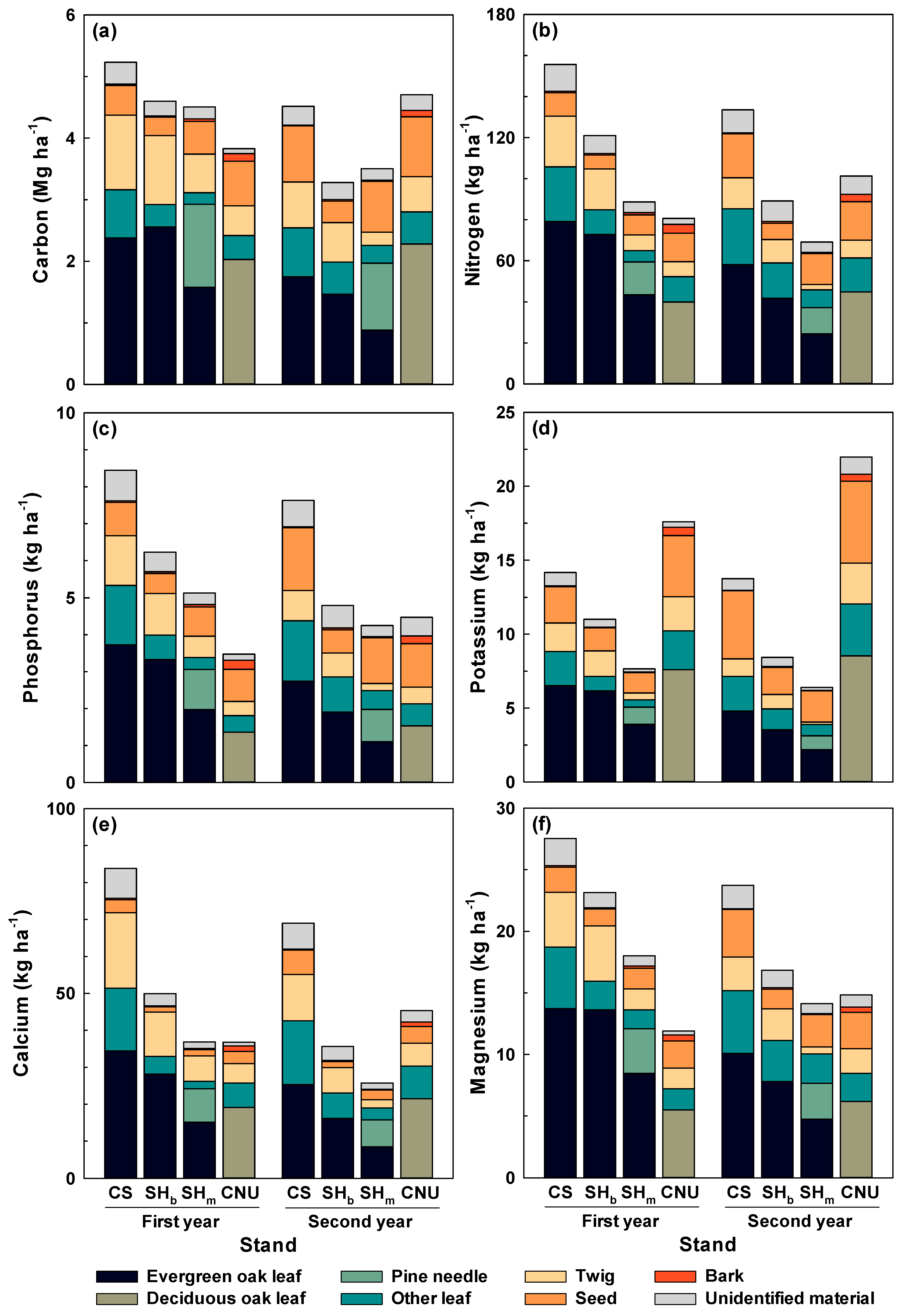
3.3. Carbon and Nutrient Inputs by Litterfall
4. Discussion
4.1. Litterfall Production in Different Forests
4.2. Carbon and Nutrient Fluxes through Litterfall
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kang, K.-N.; Park, G.-S.; Lee, S.-J.; Lee, H.-G.; Kim, J.-S.; Kim, Y.-T. Carbon storages in aboveground and root of Pinus koraiensis and Larix leptolepis stands in Gongju, Chungnam province. Korean J. Agric. Sci. 2010, 37, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, M.; Ohta, S.; Ansori, S.; Arisman, H. Nutrient dynamics via litterfall and litter decomposition on the forest floor of an Acacia mangium Willd. stand in Sumatra. Tropics 2013, 22, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Fioretto, A.; Papa, S.; Fuggi, A. Litter-fall and litter decomposition in a low Mediterranean shrubland. Biol. Fert. Soils 2003, 39, 37–44. [Google Scholar] [CrossRef]
- Onyekwelu, J.C.; Mosandl, R.; Stimm, B. Productivity, site evaluation and state of nutrition of Gmelina arborea plantations in Oluwa and Omo forest reserves, Nigeria. Forest Ecol. Manag. 2006, 229, 214–227. [Google Scholar] [CrossRef]
- Pandey, R.; Sharma, G.; Tripathi, S.; Singh, A. Litterfall, litter decomposition and nutrient dynamics in a subtropical natural oak forest and managed plantation in northeastern India. Forest Ecol. Manag. 2007, 240, 96–104. [Google Scholar] [CrossRef]
- Gairola, S.; Rawal, R.S.; Dhar, U. Patterns of litterfall and return of nutrients across anthropogenic disturbance gradients in three subalpine forests of west Himalaya, India. J. For. Res. 2009, 14, 73–80. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan, W.; Dong, W.; Liu, S. Seasonal patterns of litterfall in forest ecosystem worldwide. Ecol. Complex. 2014, 20, 240–247. [Google Scholar] [CrossRef]
- Malhi, Y.; Doughty, C.; Galbraith, D. The allocation of ecosystem net primary productivity in tropical forests. Philos. Trans. R. Soc. B. 2011, 366, 3225–3245. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [Green Version]
- Neumann, M.; Ukonmaanaho, L.; Johnson, J.; Benham, S.; Vesterdal, L.; Novotný, R.; Verstraeten, A.; Lundin, L.; Thimonier, A.; Michopoulos, P.; et al. Quantifying Carbon and Nutrient Input from Litterfall in European Forests Using Field Observations and Modeling. Global Biogeochem. Cy. 2018, 32, 784–798. [Google Scholar] [CrossRef]
- Xu, X.; Hirata, E.; Shibata, H. Effect of typhoon disturbance on fine litterfall and related nutrient input in a subtropical forest on Okinawa Island, Japan. Basic Appl. Ecol. 2004, 5, 271–282. [Google Scholar] [CrossRef]
- Werneck, M.S.; Pedralli, G.; Gieseke, L.F. Litterfall in three sites of semideciduous forest with different disturbance degree in the Tripui Ecological Station, Ouro Preto, MG. Rev. Bras. Bot. 2001, 24, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Pires, L.A.; Britez, R.M.; Martel, G.; Pagano, S.N. Litterfall, accumulation and decomposition in a restinga at Ilha do Mel, Paranagua, Parana, Brazil. Acta Bot. Bras. 2006, 20, 173–184. [Google Scholar] [CrossRef]
- Lopes, M.C.A.; Araújo, V.F.P.; Vasconcellos, A. The effects of rainfall and vegetation on litterfall production in the semiarid region of northeastern Brazil. Braz. J. Biol. 2015, 75, 703–708. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Westman, C.J.; Berg, B.; Kutsch, W.; Wang, G.Z.; Man, R.; Ilvesniemi, H. Variation in litterfall-climate relationships between coniferous and broadleaf forests in Eurasia. Global Ecol. Biogeogr. 2004, 13, 105–114. [Google Scholar] [CrossRef]
- Scheer, M.B.; Gatti, G.; Wisniewski, C.; Mocochinski, A.Y.; Cavassani, A.T.; Lorenzetto, A.; Putini, F. Patterns of litter production in a secondary alluvial Atlantic Rain Forest in southern Brazil. Revista Brasileira De Botânica 2009, 32, 805–817. [Google Scholar] [CrossRef] [Green Version]
- Kouki, J.; Hokkanen, T. Long-term needle litterfall of a Scots pine Pinus sylvestris stand: Relation to temperature factors. Oecologia 1992, 89, 176–181. [Google Scholar] [CrossRef]
- Liu, C.; Berg, B.; Kutsch, W.; Westman, C.J.; Ilvesniemi, H.; Shen, X.; Shen, G.; Chen, X. Leaf litter nitrogen concentration as related to climatic factors in Eurasian forests. Global Ecol. Biogeogr. 2006, 15, 438–444. [Google Scholar] [CrossRef]
- Bigelow, S.W.; Canham, C.D. Litterfall as a niche construction process in a northern hardwood forest. Ecosphere 2015, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kamruzzaman, M.; Basak, K.; Paul, S.K.; Ahmed, S.; Osawa, A. Litterfall production, decomposition and nutrient accumulation in Sundarbans mangrove forests, Bangladesh. For. Sci. Technol. 2019, 15, 24–32. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Morse, C.W.; Buxton, R.P.; Bonner, K.I.; Mason, N.W.H.; Wardle, D.A. Litterfall, nutrient concentrations and decomposability of litter in a New Zealand temperate montane rain forest. N. Z. J. Ecol. 2013, 37, 162–171. [Google Scholar]
- Meentemeyer, V.; Box, E.O.; Thompson, R. World Patterns and Amounts of Terrestrial Plant Litter Production. BioScience 1982, 32, 125–128. [Google Scholar] [CrossRef]
- Park, J.B. Characterization of lava-formed petrology and petrochemistry; Institute of Environmental Resource Research: Jeju Special Self-Governing Province, Korea, 2010. [Google Scholar]
- Choi, H.-M.; Lee, J.-Y. Changes of groundwater conditions on Jeju volcanic island, Korea: Implications for sustainable agriculture. Afr. J. Agric. Res. 2012, 7, 647–661. [Google Scholar] [CrossRef]
- Cho, I.-Y.; Kang, D.-W.; Kang, J.; Hwang, H.; Won, J.-H.; Paek, W.K.; Seo, S.-Y. A study on the biodiversity of benthic invertebrates in the waters of Seogwipo, Jeju Island, Korea. J. Asia Pac. Biodivers. 2014, 7, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.Y.; Nason, J.D.; Sun, B.Y.; Moon, M.-O.; Chung, J.M.; Park, C.-W.; Chung, M.G. Extremely low levels of genetic variation in the Critically Endangered monotypic fern genus Mankyua chejuense (Ophioglossaceae) from Korea: Implications for conservation. Biochem. Syst Ecol. 2010, 38, 888–896. [Google Scholar] [CrossRef]
- Shin, J.S.; George, S. Composition and genesis of volcanic ash soils in Jeju Island, I. Physico-chemical and macro-micromorphological properties. J. Miner. Soc. Korea 1988, 1, 32–39. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kim, D.-S.; Lee, K.C.; Lee, J.-S.; King, G.M.; Kang, S. Microbial community structure and functional potential of lava-formed Gotjawal soils in Jeju, Korea. PLoS ONE 2018, 13, e0204761. [Google Scholar] [CrossRef]
- Jeong, K.J. A study on perception and use of Gotjawal in Jeju Island. J. Photo-Geogr. 2012, 22, 11–28. [Google Scholar]
- Kang, H.-G.; Kim, C.-S.; Kim, E.-S. Human influence, regeneration, and conservation of the Gotjawal forests in Jeju Island, Korea. J. Mar. Isl. Cult. 2013, 2, 85–92. [Google Scholar] [CrossRef] [Green Version]
- An, J.Y.; Han, S.H.; Youn, W.B.; Lee, S.I.; Rahman, A.; Dao, H.T.T.; Seo, J.M.; Aung, A.; Choi, H.S.; Park, B.B. Comparison of litterfall production in three forest types in Jeju Island, South Korea. J. Forestry Res. 2019. [Google Scholar] [CrossRef]
- Rahman, A. Carbon and nutrient cycling by litterfall and litter decomposition at three different forests in South Korea. Master’s Thesis, Chungnam National University, Daejeon, Korea, August 2019. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; p. 256. [Google Scholar]
- Peet, R.K. The Measurement of Species Diversity, Annual Review of Ecology and Systematics. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- Watanabe, F.S.; Olsen, S.R. Test of an Ascorbic Acid Method for Determining Phosphorus in Water and NaHCO3 Extracts from Soil1. Soil Sci. Soc. Am. J. 1965, 29, 677. [Google Scholar] [CrossRef]
- Oh, J.G.; Lee, S.Y.; Kim, D.C.; Baek, S.Y.; Lee, K.J. A study on the litter decomposition and distribution of related to location condition, Jeju Island. Proc. Korean Soc. Environ. Ecol. Con. 2018, 49–50. [Google Scholar]
- Shen, G.; Chen, D.; Wu, Y.; Liu, L.; Liu, C. Spatial patterns and estimates of global forest litterfall. Ecosphere 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Fernando, E.S.; Balatibat, J.B.; Peras, J.R.; Jumawid, R.J.J.; Benavente, A.B., Jr.; Bautista, R.R.; dela Cruz, S. Resource inventory and assessment of biodiversity in the Subic Bay Metropolitan Authority (SBMA). Termin. Rep. 1998, 1–65. [Google Scholar]
- Pedersen, L.B.; Bille-Hansen, J. A comparison of litterfall and element fluxes in even aged Norway spruce, sitka spruce and beech stands in Denmark. Forest Ecol. Manag. 1999, 114, 55–70. [Google Scholar] [CrossRef]
- Pretzsch, H.; Block, J.; Dieler, J.; Dong, P.H.; Kohnle, U.; Nagel, J.; Spellmann, H.; Zingg, A. Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann. For. Sci. 2010, 67, 712. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Ma, Y.; Zhao, K.; Niklaus, P.A.; Schmid, B.; He, J.S. Positive effects of tree species diversity on litterfall quantity and quality along a secondary successional chronosequence in a subtropical forest. J. Plant. Ecol. 2017, 10, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Zhou, X.; Wen, Y.; Zhu, H.; You, Y.; Qin, Z.; Li, Y.; Huang, X.; Yan, L.; Li, H.; et al. Coniferous-Broadleaf Mixture Increases Soil Microbial Biomass and Functions Accompanied by Improved Stand Biomass and Litter Production in Subtropical China. Forests 2019, 10, 879. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, S.; Huang, Y. Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. Forest Ecol. Manag. 2008, 255, 1210–1218. [Google Scholar] [CrossRef]
- Bray, J.R.; Gorham, E. Litter Production in Forests of the World. Adv. Ecol. Res. 1964, 2, 101–157. [Google Scholar] [CrossRef]
- Albrektson, A. Needle litterfall in stands of Pinus sylvestris L. in Sweden, in relation to site quality, stand age and latitude. Scand. J. For. Res. 1998, 3, 333–342. [Google Scholar] [CrossRef]
- Zhou, G.; Guan, L.; Wei, X.; Zhang, D.; Zhang, Q.; Yan, J.; Wen, D.; Liu, J.; Liu, S.; Huang, Z.; et al. Litterfall Production Along Successional and Altitudinal Gradients of Subtropical Monsoon Evergreen Broadleaved Forests in Guangdong, China. Plant Ecol. 2006, 188, 77–89. [Google Scholar] [CrossRef]
- Kelty, M.J. The role of species mixtures in plantation forestry. For. Ecol. Manag. 2006, 233, 195–204. [Google Scholar] [CrossRef]
- Lu, S.W.; Liu, C.P. Patterns of litterfall and nutrient return at different altitudes in evergreen hardwood forests of Central Taiwan. Ann. For. Sci. 2012, 69, 877–886. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Yang, W.; Tan, B.; Xu, Z.; Zhang, Y.; Yang, J.; Ni, X.; Wu, F. Seasonal Dynamics of Litterfall in a Sub-Alpine Spruce-Fir Forest on the Eastern Tibetan Plateau: Allometric Scaling Relationships Based on One Year of Observations. Forests 2017, 8, 314. [Google Scholar] [CrossRef] [Green Version]
- Vyse, K.; Pagter, M.; Zuther, E.; Hincha, D.K. Deacclimation after cold acclimation—a crucial, but widely neglected part of plant winter survival. J. Exp. Bot. 2019, 70, 4595–4604. [Google Scholar] [CrossRef] [Green Version]
- Aguilos, M.M.; Takagi, K.; Takahashi, H.; Hasegawa, J.; Ashiya, D.; Kotsuka, C.; Naniwa, A.; Sakai, R.; Ito, K.; Miyoshi, C.; et al. Enhanced annual litterfall production due to spring solar radiation in cool-temperate mixed forests of northern Hokkaido, Japan. J. Agric. Meteorol. 2012, 68, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Goulden, M.L.; Munger, J.W.; Fan, S.M.; Daube, B.C.; Wofsy, S.C. Exchange of carbon dioxide by a deciduous forest: Response to interannual climate variability. Science 1996, 271, 1576–1578. [Google Scholar] [CrossRef] [Green Version]
- Reiners, W.A.; Lang, G.E. Changes in Litterfall along a Gradient in Altitude. J. Ecol. 1987, 75, 629–638. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, J.F.; Chen, G.S.; Xie, J.S.; Gao, R.; Li, Z.; Jin, Z. Litter production, seasonal pattern and nutrient return in seven natural forests compared with a plantation in southern China. Forestry 2005, 78, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.H.; Mc Graw, R.L.; George, M.F.; Garrett, H.E. Shade effects on forage crops with potential in temperate agroforestry practices. Agroforest. Syst. 1999, 44, 109–119. [Google Scholar] [CrossRef]
- Liang, H.W. Studies on the litterfall of two forest types in mid-altitude of Laoshan mountain in Tianlin Country. Chin. J. Ecol. 1994, 13, 21–26. [Google Scholar]
- Lin, K.C.; Hamburg, S.P.; Tang, S.L.; Hsia, Y.J.; Lin, T.C. Typhoon effects on litterfall in a subtropical forest. Can. J. For. Res. 2003, 2184–2192. [Google Scholar] [CrossRef]
- Baldocchi, D.D.; Ma, S.; Rambal, S.; Misson, L.; Ourcival, J.M.; Limousin, J.M.; Pereira, J.; Papale, D. On the differential advantages of evergreenness and deciduousness in mediterranean oak woodlands: A flux perspective. Ecol. Appl. 2010, 20, 1583–1597. [Google Scholar] [CrossRef]
- Royer, D.L.; Osborne, C.P.; Beerling, D.J. Contrasting seasonal patterns of carbon gain in evergreen and deciduous trees of ancient polar forests. Paleobiology 2005, 31, 141–150. [Google Scholar] [CrossRef]
- Tomlinson, K.W.; Poorter, L.; Sterck, F.J.; Borghetti, F.; Ward, D.; Bie, S.D.; Langevelde, F.V. Leaf adaptations of evergreen and deciduous trees of semi-arid and humid savannas on three continents. J. Ecol. 2013, 101, 430–440. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Thompson, K. Functional leaf attributes predict litter decomposition rate in herbaceous plants. New Phytol. 1997, 135, 109–114. [Google Scholar] [CrossRef]
- Kikuzawa, K. A Cost-Benefit Analysis of Leaf Habit and Leaf Longevity of Trees and Their Geographical Pattern. Am. Nat. 1991, 138, 1250–1263. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [Green Version]
- Givnish, T. Adaptive significance of evergreen vs. deciduous leaves: Solving the triple paradox. Silva. Fenn. 2002, 36, 703–743. [Google Scholar] [CrossRef]
- Hollinger, D.Y. Leaf and Simulated Whole-Canopy Photosynthesis in Two Co-Occurring Tree Species. Ecology 1992, 73, 1–14. [Google Scholar] [CrossRef]
- Mooney, H.A.; Dunn, E.L. Photosynthetic Systems of Mediterranean-Climate Shrubs and Trees of California and Chile. Am. Nat. 1970, 104, 447–453. [Google Scholar] [CrossRef]
- Park, W.-P.; Song, K.-C.; Koo, B.-J.; Hyun, H.-N. Distribution of Available Silicon of Volcanic Ash Soils in Jeju Island. App. Environm. Soil Sci. 2019, 1–10. [Google Scholar] [CrossRef]
- Takahashi, T.; Dahlgren, R.A. Nature, properties and function of aluminum–humus complexes in volcanic soils. Geoderma 2016, 263, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Moon, K.-H.; Lim, H.-C.; Hyun, H.-N. Distribution of organic matter and Alo +1/2Feo contents in soils using principal component and multiple regression analysis in Jeju Island. Korean J. Soil Sci. Fert. 2010, 43, 748–754. [Google Scholar]
- Nanzyo, M.; Shibata, Y.; Wada, N. Complete contact of Brassica roots without phosphates in a phosphorus-deficient. Soil Sci. Plant Nutr. 2002, 48, 847–853. [Google Scholar] [CrossRef]
- Greger, M.; Bertell, G. Effects of Ca2+ and Cd2+ on the carbohydrate metabolism in sugar beet (Beta vulgaris). J. Exp. Bot. 1992, 43, 167–173. [Google Scholar] [CrossRef]
- Pontigo, S.; Ribera, A.; Gianfreda, L.; Mora, M.D.L.L.; Nikolic, M.; Cartes, P. Silicon in vascular plants: Uptake, transport and its influence on mineral stress under acidic conditions. Planta 2015, 242, 23–37. [Google Scholar] [CrossRef]
- Nascimento, C.W.A.D.; Nunes, G.H.D.S.; Preston, H.A.F.; Silva, F.B.V.D.; Preston, W.; Loureiro, F.L.C. Influence of silicon fertilization on nutrient accumulation, yield and fruit quality of melon grown in Northeastern Brazil. Silicon 2019. [Google Scholar] [CrossRef]
- Schaller, J.; Faucherre, S.; Joss, H.; Obst, M.; Goeckede, M.; Planer-Friedrich, B.; Peiffer, S.; Gilfedder, B.; Elberling, B. Silicon increases the phosphorus availability of Arctic soils. Sci. Rep.-UK 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, E.R.M.; Tomlinson, K.W.; Carvalheiro, L.G.; Kirkman, K.; Bie, S.D.; Prins, H.H.T.; Langevelde, F.V. Short-term effect of nutrient availability and rainfall distribution on biomass production and leaf nutrient content of savanna tree species. PLoS ONE 2014, 9, 1–9. [Google Scholar] [CrossRef]
- Jiang, D.; Geng, Q.; Li, Q.; Luo, Y.; Vogel, J.; Shi, Z.; Ruan, H.; Xu, X. Nitrogen and phosphorus resorption in planted forests worldwide. Forests 2019, 10, 201. [Google Scholar] [CrossRef] [Green Version]
- Lodge, D.J.; Mcdowell, W.H. Summary of ecosystem-level effects of Caribbean hurricanes. Biotropica 1991, 23, 373. [Google Scholar] [CrossRef]
- Golley, F.B.; McGimmis, J.T.; Clements, R.G.; Child, G.I.; Duever, M.J. Mineral. Cycling in a Tropical Moist Forest Ecosystem; University of Georgia Press: Athens, GA, USA, 1975; p. 248. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand | CS1 | SHb2 | SHm3 | CNU4 | ||||
|---|---|---|---|---|---|---|---|---|
| Forest Type | Evergreen Broadleaved | Evergreen Broadleaved | Evergreen Mixed | Deciduous Broadleaved | ||||
| All tree species | ||||||||
| DBH (cm) | 12.5 | (0.3) | 19.1 | (0.2) | 21.6 | (0.9) | 20.2 | (1.8) |
| Height (m) | 9.4 | (0.2) | 10.7 | (0.2) | 10.9 | (0.3) | 12.5 | (0.5) |
| BA (m2 ha−1) 5 | 34.0 | (0.4) | 50.7 | (0.6) | 50.9 | (0.4) | 33.2 | (4.4) |
| Density (ha−1) | 2475 | (51) | 1300 | (25) | 1075 | (35) | 900 | (135) |
| Dominant species | ||||||||
| Oak 6 | ||||||||
| DBH (cm) | 12.4 | (0.5) | 27.3 | (2.2) | 20.7 | (1.1) | 21.4 | (1.9) |
| Height (m) | 9.5 | (0.2) | 13.0 | (0.5) | 10.9 | (0.2) | 13.8 | (0.9) |
| BA (m2 ha−1) 5 | 30.3 | (0.2) | 44.4 | (0.8) | 28.5 | (0.3) | 18.5 | (3.2) |
| P. thunbergii | ||||||||
| DBH (cm) | - | - | 31.4 | (1.9) | - | |||
| Height (m) | - | - | 13.2 | (0.9) | - | |||
| BA (m2 ha−1) 5 | - | - | 18.7 | (3.2) | - | |||
| Diversity indices | ||||||||
| Shannon’s diversity index 7 | 1.4 | 2.1 | 1.4 | 1.5 | ||||
| Shannon’s equitability | 0.5 | 0.8 | 0.5 | 0.7 | ||||
| Simpson’s diversity index | 2.2 | 5.9 | 2.5 | 3.2 | ||||
| Simpson’s equitability | 0.1 | 0.4 | 0.2 | 0.4 | ||||
| Stand | Season | Stand×Season | ||
|---|---|---|---|---|
| Litter components | df | 3 | 7 | 21 |
| Leaf litterfall | ||||
| Oak1 | <0.0001 | <0.0001 | <0.0001 | |
| Pine2 | <0.0001 | 0.0263 | 0.0026 | |
| Other species | <0.0001 | <0.0001 | <0.0001 | |
| Total leaf | 0.0476 | <0.0001 | <0.0001 | |
| Other litterfall | ||||
| Twig | <0.0001 | <0.0001 | <0.0001 | |
| Seed | <0.0001 | <0.0001 | <0.0001 | |
| Bark | <0.0001 | 0.0087 | 0.52183 | |
| Unidentified material | <0.0001 | <0.0001 | <0.0001 | |
| Total litterfall | 0.0002 | 0.0002 | <0.0001 | |
| Stand | Litter Component | Carbon (%) | Nitrogen (%) | Phosphorus (%) | Potassium (%) | Calcium (%) | Magnesium (%) |
|---|---|---|---|---|---|---|---|
| CS1 | EO5 leaf | 47.48 (0.21) | 1.58 (0.05) | 0.07 (0.00) | 0.13 (0.01) | 0.69 (0.04) | 0.27 (0.01) |
| Other leaf | 46.98 (0.37) | 1.60 (0.10) | 0.10 (0.00) | 0.14 (0.02) | 1.02 (0.07) | 0.30 (0.04) | |
| Twig | 48.08 (0.19) | 0.98 (0.10) | 0.05 (0.00) | 0.08 (0.01) | 0.81 (0.08) | 0.18 (0.01) | |
| Seed | 47.25 (1.27) | 1.11 (0.19) | 0.09 (0.01) | 0.24 (0.04) | 0.34 (0.05) | 0.20 (0.04) | |
| Bark | 47.11 (0.24) | 1.72 (0.09) | 0.11 (0.01) | 0.12 (0.00) | 1.07 (0.13) | 0.29 (0.02) | |
| SHb2 | EO leaf | 48.76 (0.28) | 1.39(0.05) | 0.06 (0.01) | 0.12 (0.01) | 0.54 (0.03) | 0.26 (0.03) |
| Other leaf | 47.65 (0.29) | 1.57 (0.07) | 0.09 (0.00) | 0.13 (0.01) | 0.63 (0.07) | 0.30 (0.02) | |
| Twig | 48.12 (0.25) | 0.86 (0.05) | 0.05 (0.00) | 0.07 (0.01) | 0.52 (0.03) | 0.19 (0.02) | |
| Seed | 47.19 (0.08) | 1.07 (0.09) | 0.08 (0.01) | 0.25 (0.06) | 0.22 (0.00) | 0.21 (0.04) | |
| Bark | 47.49 (0.22) | 1.73 (0.11) | 0.11 (0.01) | 0.11 (0.01) | 0.65 (0.04) | 0.25 (0.02) | |
| SHm3 | EO leaf | 48.13 (0.11) | 1.32 (0.04) | 0.06 (0.00) | 0.12 (0.01) | 0.46 (0.02) | 0.26 (0.01) |
| Pine6 needle | 51.01 (0.09) | 0.60 (0.03) | 0.04 (0.00) | 0.04 (0.00) | 0.34 (0.02) | 0.14 (0.01) | |
| Other leaf | 46.15 (0.63) | 1.37 (0.11) | 0.08 (0.01) | 0.12 (0.02) | 0.50 (0.07) | 0.38 (0.02) | |
| Twig | 50.30 (0.73) | 0.61 (0.07) | 0.05 (0.00) | 0.04 (0.00) | 0.55 (0.06) | 0.14 (0.02) | |
| Seed | 48.74 (0.89) | 0.89 (0.10) | 0.07 (0.00) | 0.13 (0.01) | 0.15 (0.03) | 0.15 (0.01) | |
| Bark | 48.68 (0.61) | 1.32 (0.25) | 0.08 (0.01) | 0.05 (0.01) | 0.44 (0.05) | 0.21 (0.03) | |
| CNU4 | DO7 leaf | 49.11 (0.46) | 0.97(0.09) | 0.03 (0.00) | 0.18 (0.01) | 0.46 (0.01) | 0.13 (0.01) |
| Other leaf | 47.69 (0.34) | 1.52 (0.06) | 0.05 (0.00) | 0.32 (0.02) | 0.81 (0.14) | 0.21 (0.02) | |
| Twig | 48.69 (0.22) | 0.73 (0.04) | 0.04 (0.00) | 0.23 (0.01) | 0.53 (0.01) | 0.17 (0.02) | |
| Seed | 48.50 (0.19) | 0.93 (0.12) | 0.06 (0.00) | 0.28 (0.05) | 0.22 (0.05) | 0.15 (0.01) | |
| Bark | 44.29 (1.69) | 1.55 (0.19) | 0.09 (0.01) | 0.20 (0.03) | 0.53 (0.14) | 0.17 (0.01) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, B.B.; Rahman, A.; Han, S.H.; Youn, W.B.; Hyun, H.J.; Hernandez, J.; An, J.Y. Carbon and Nutrient Inputs by Litterfall in Evergreen and Deciduous Forests in Korea. Forests 2020, 11, 143. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020143
Park BB, Rahman A, Han SH, Youn WB, Hyun HJ, Hernandez J, An JY. Carbon and Nutrient Inputs by Litterfall in Evergreen and Deciduous Forests in Korea. Forests. 2020; 11(2):143. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020143
Chicago/Turabian StylePark, Byung Bae, Afroja Rahman, Si Ho Han, Woo Bin Youn, Hwa Ja Hyun, Jonathan Hernandez, and Ji Young An. 2020. "Carbon and Nutrient Inputs by Litterfall in Evergreen and Deciduous Forests in Korea" Forests 11, no. 2: 143. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020143