
Dynamic Changes in Plant Resource Use Efficiencies and Their Primary Influence Mechanisms in a Typical Desert Shrub Community

Abstract
:1. Introduction
2. Methods
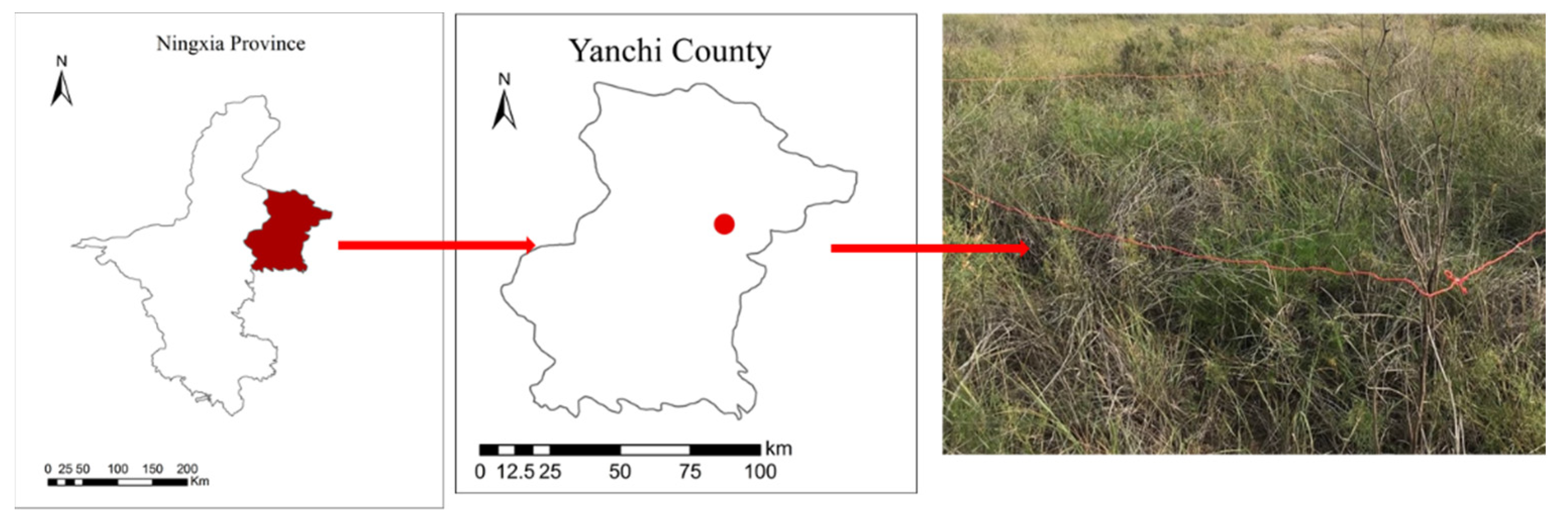
2.1. Site Description
2.2. Photosynthesis Gas-Exchange Measurements
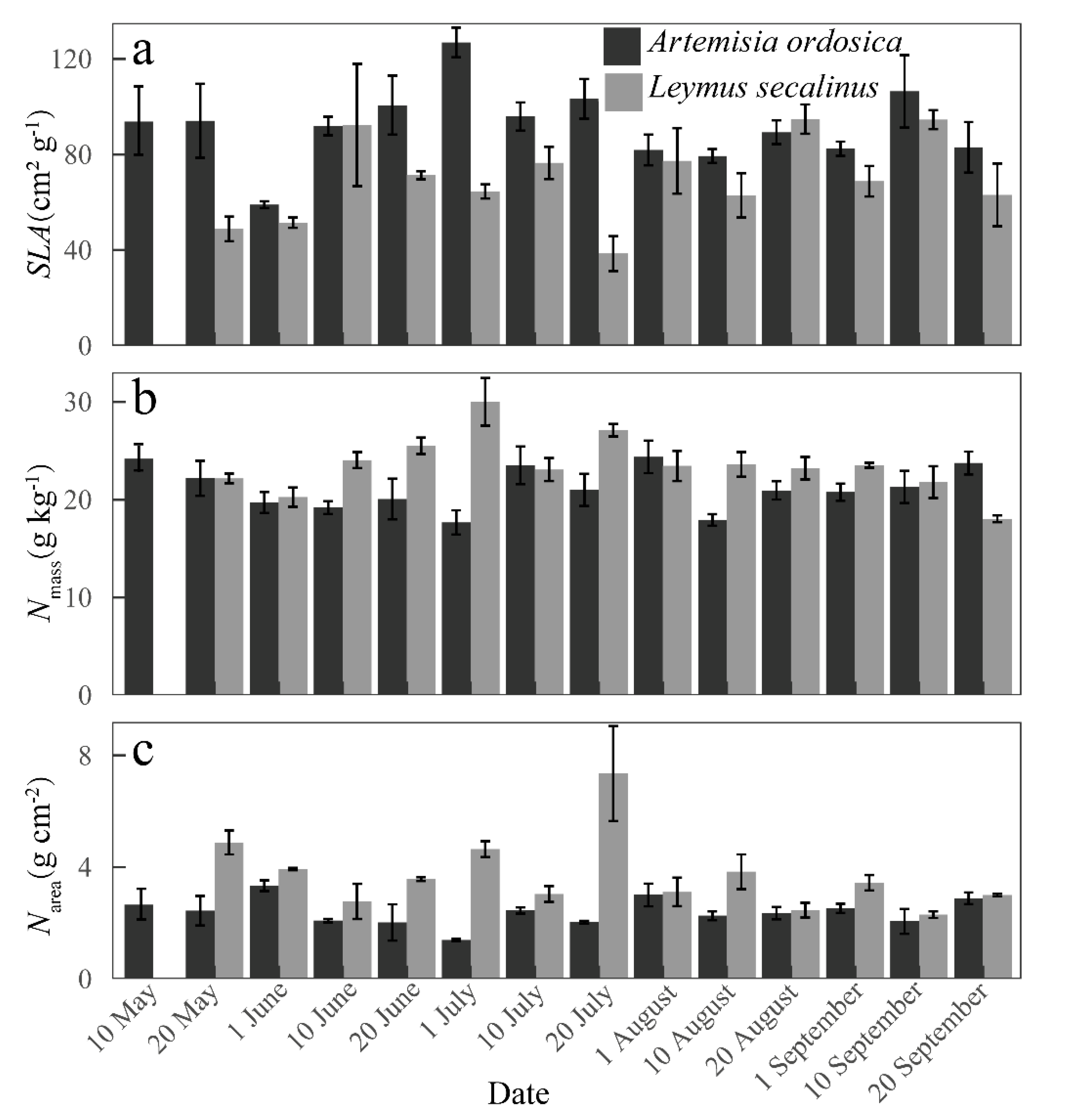
2.3. Measurement of Biotic Factors
2.4. Measurement of Abiotic Factors
2.5. Data Processing and Analysis
2.6. Statistical Analysis
3. Results
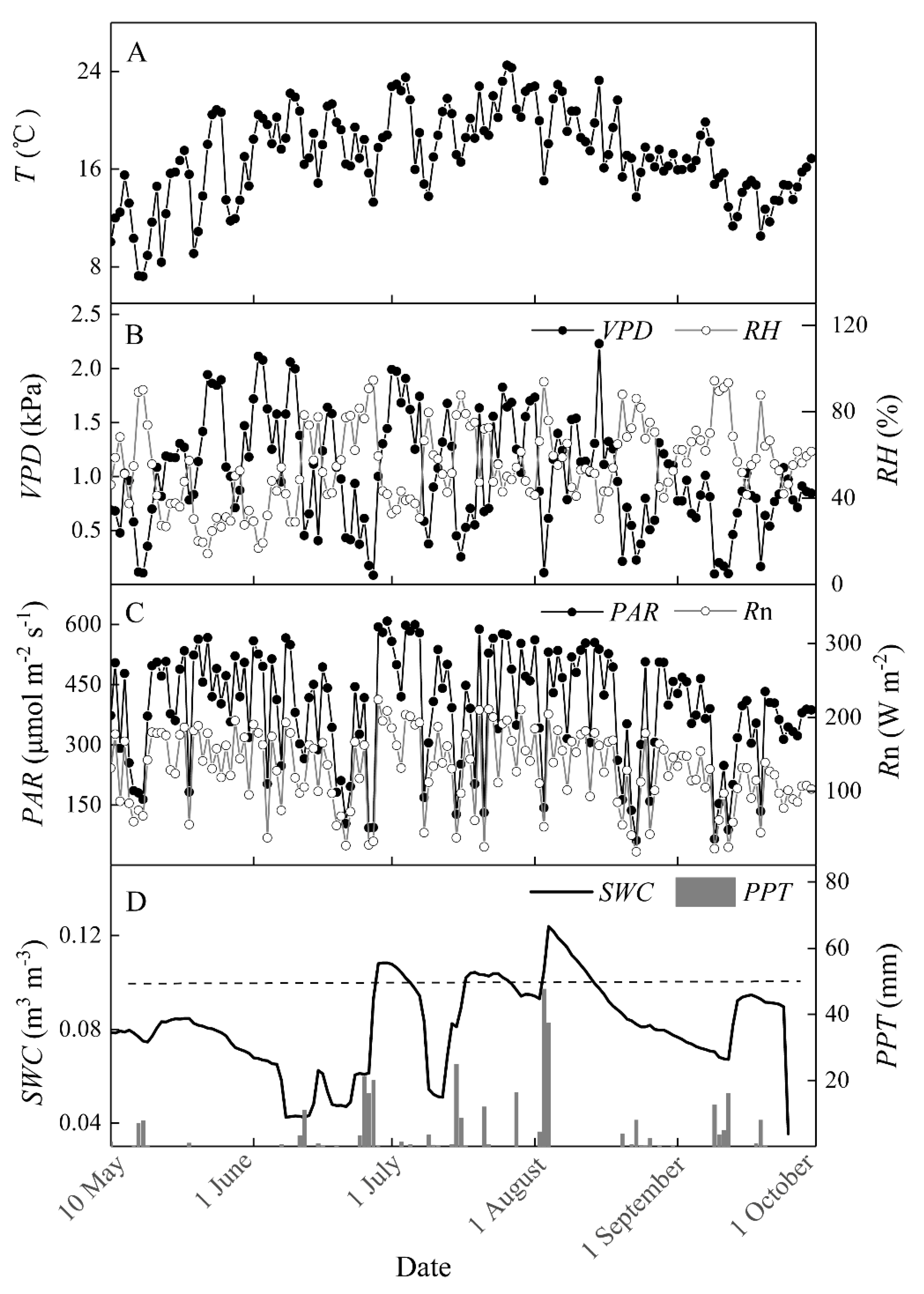
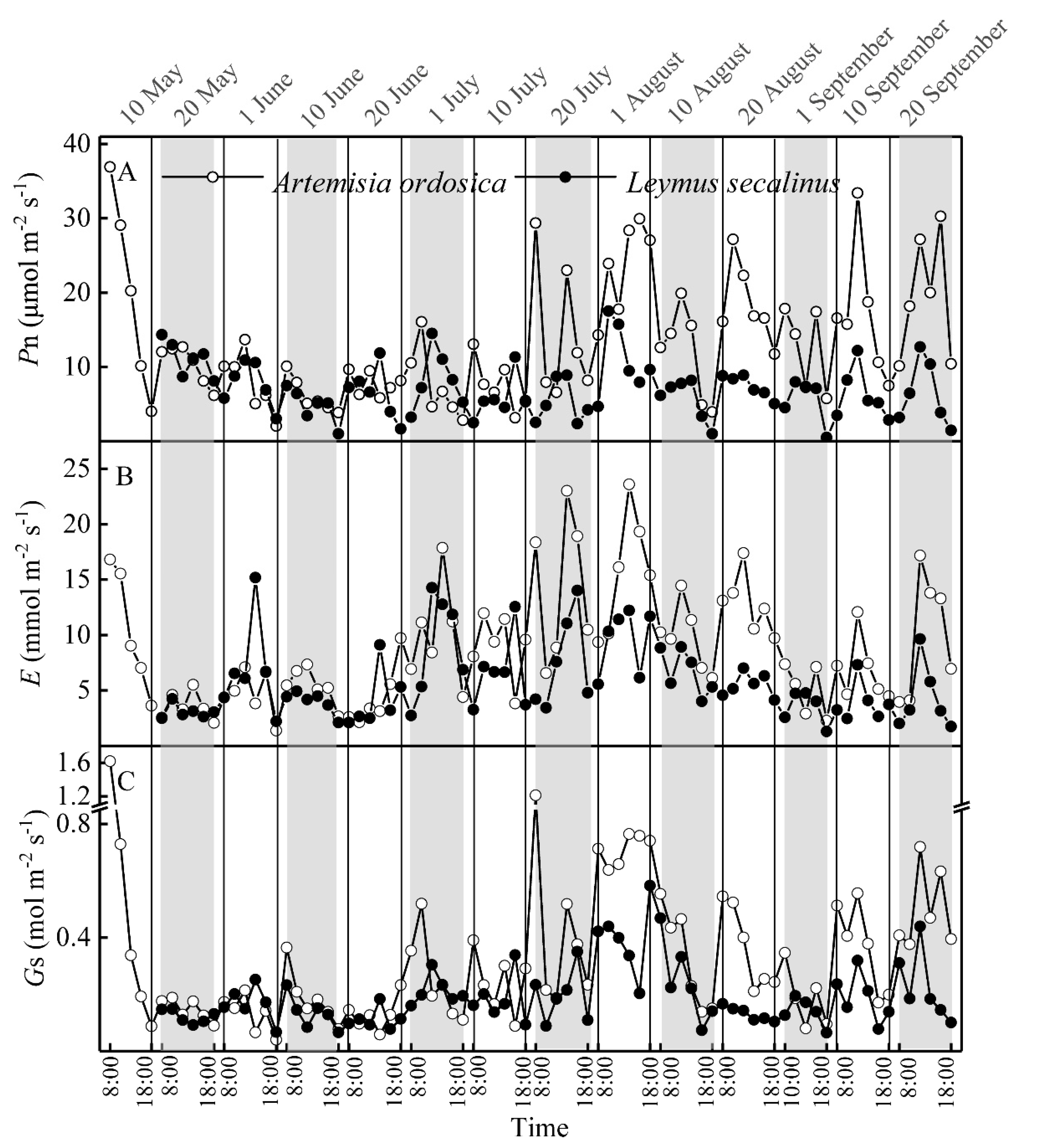
3.1. Variations in Biophysical Factors and Photosynthetic Parameters
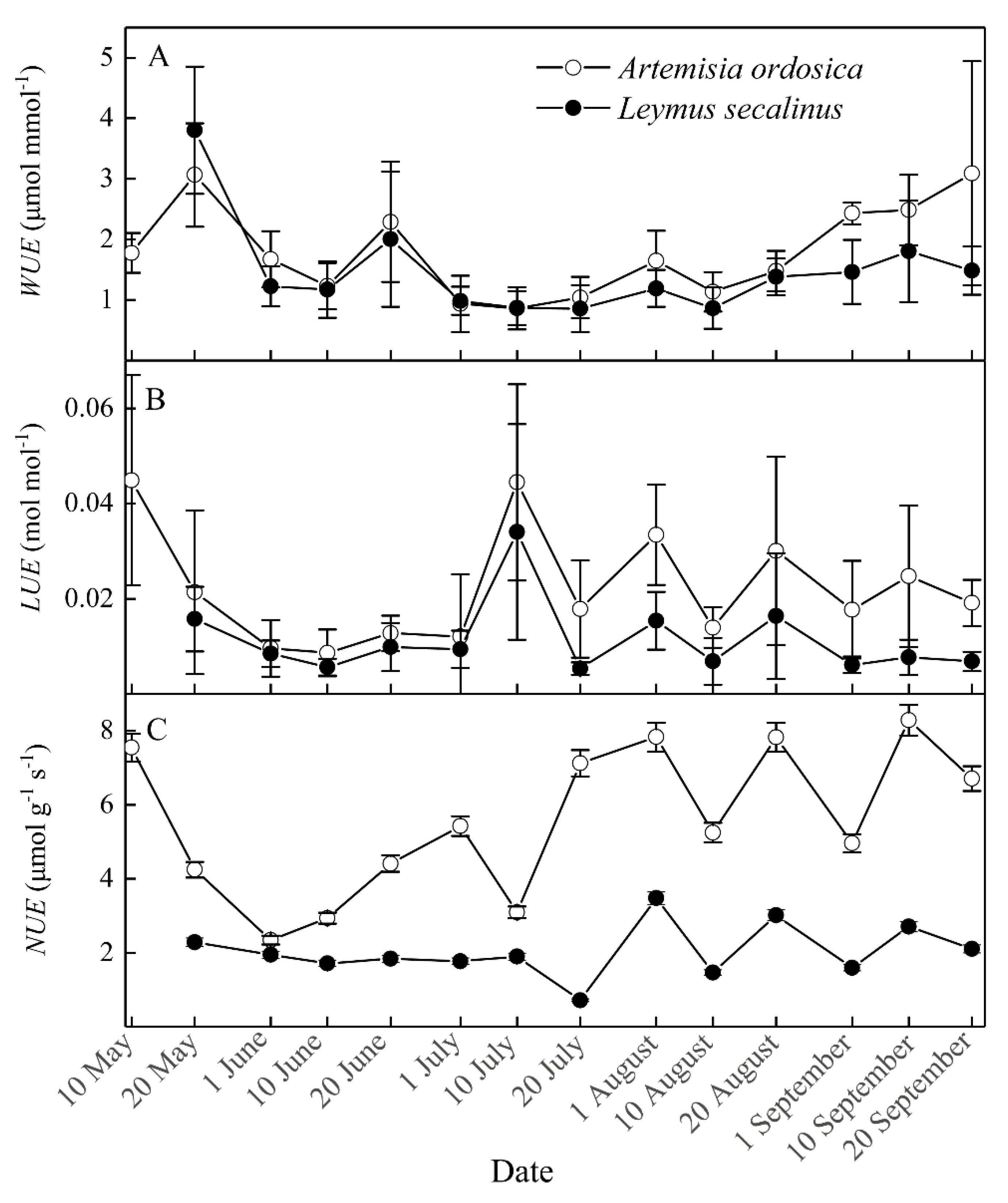
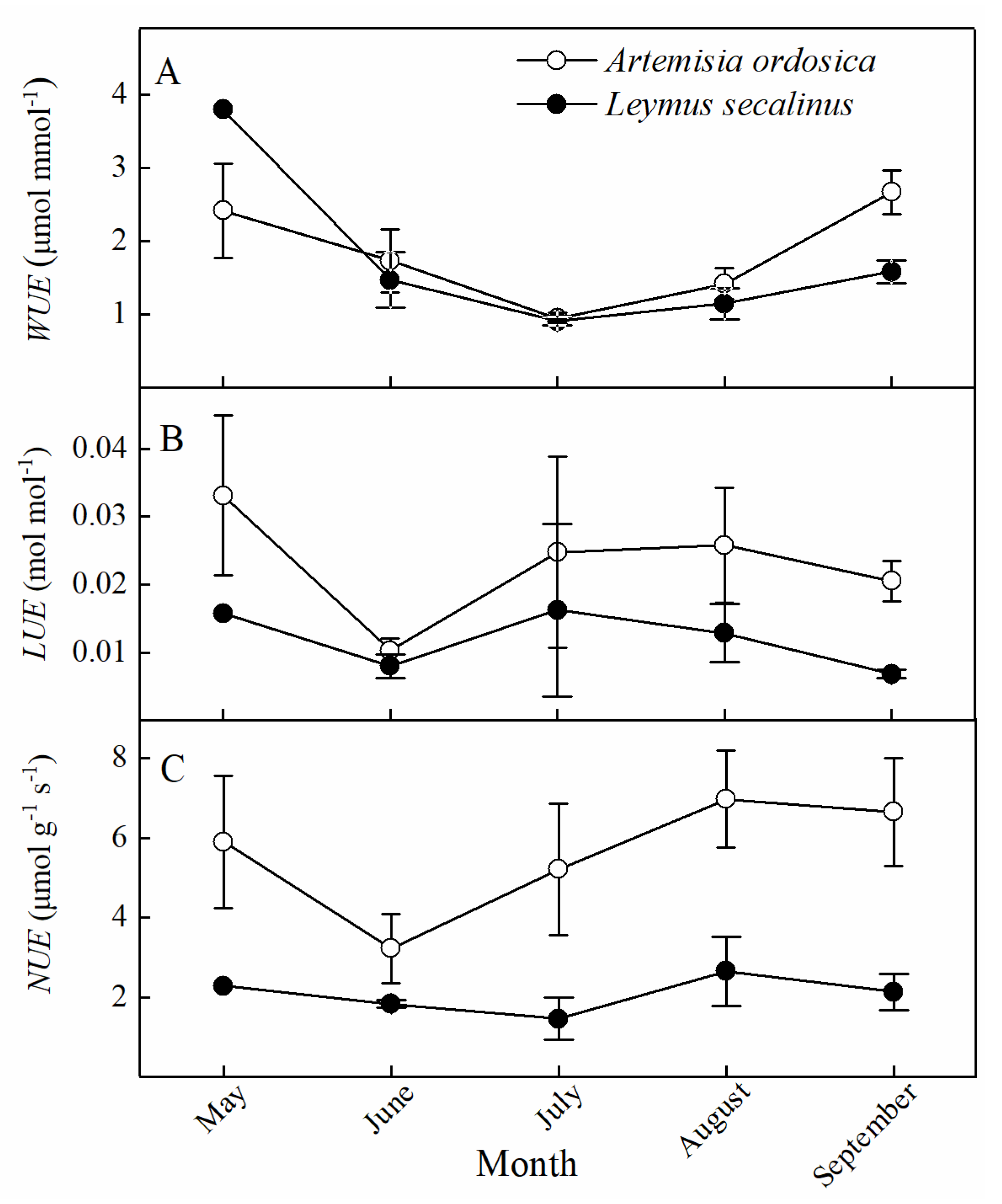
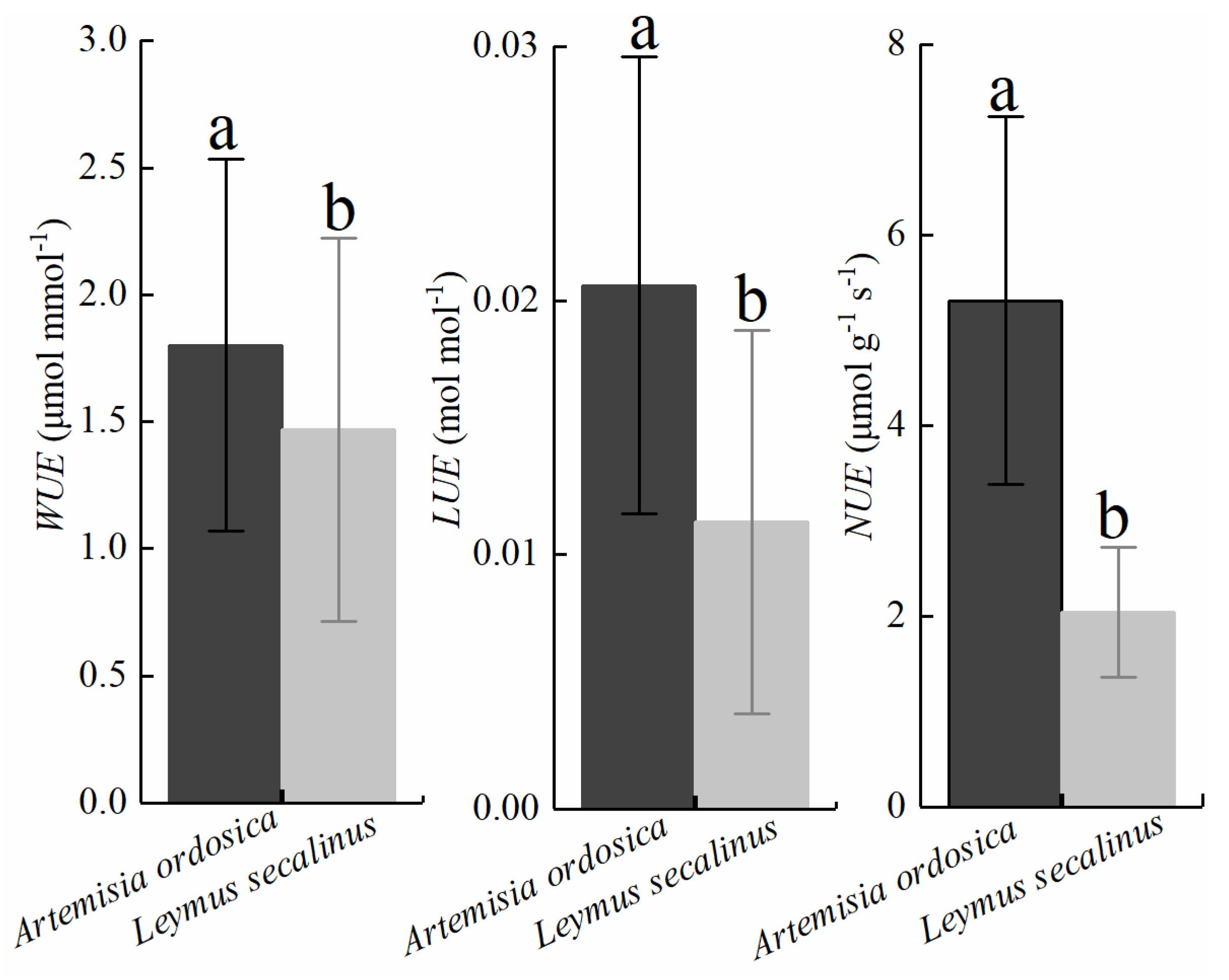
3.2. Seasonal Dynamics in Plant Resource Use Efficiencies
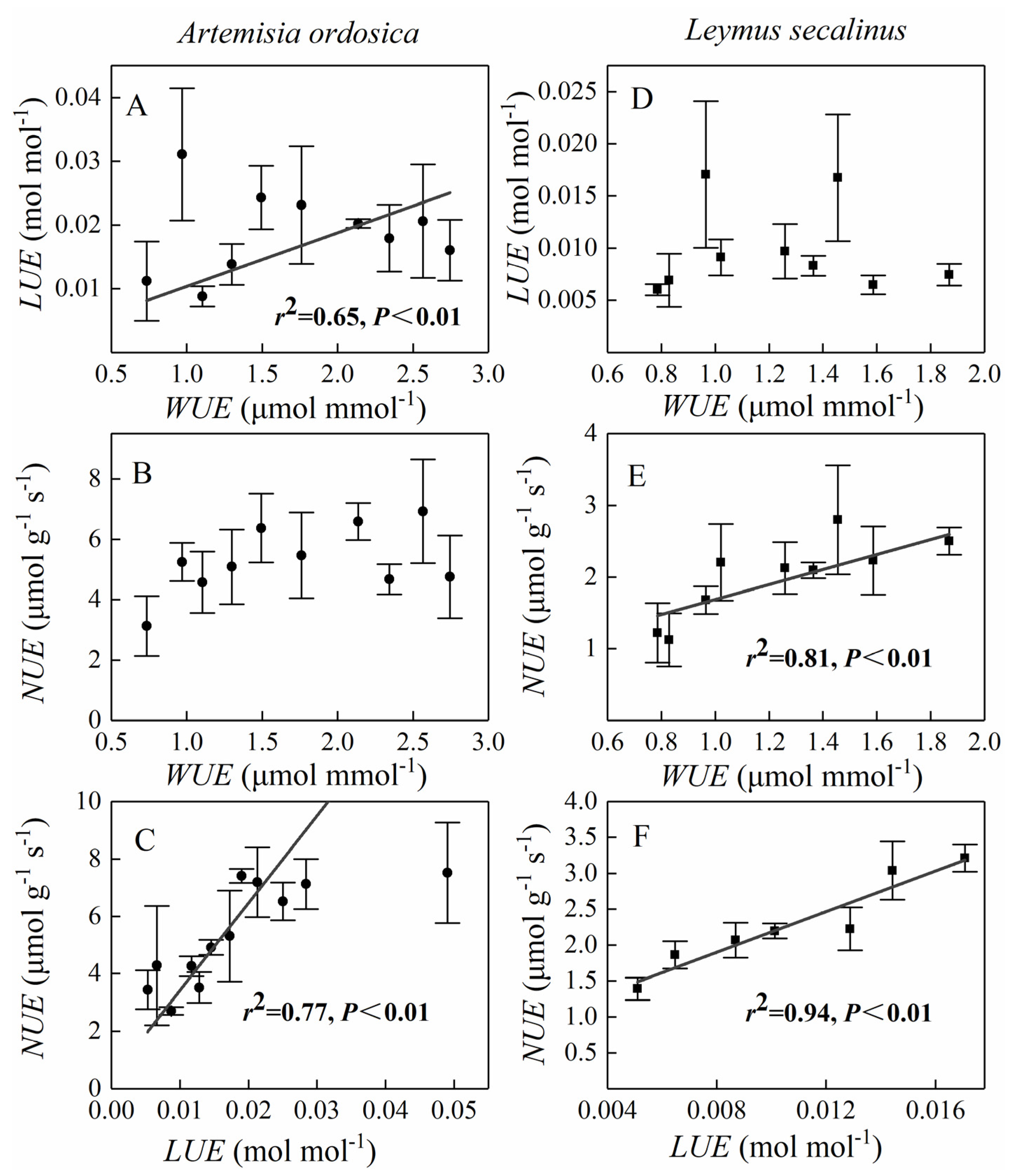
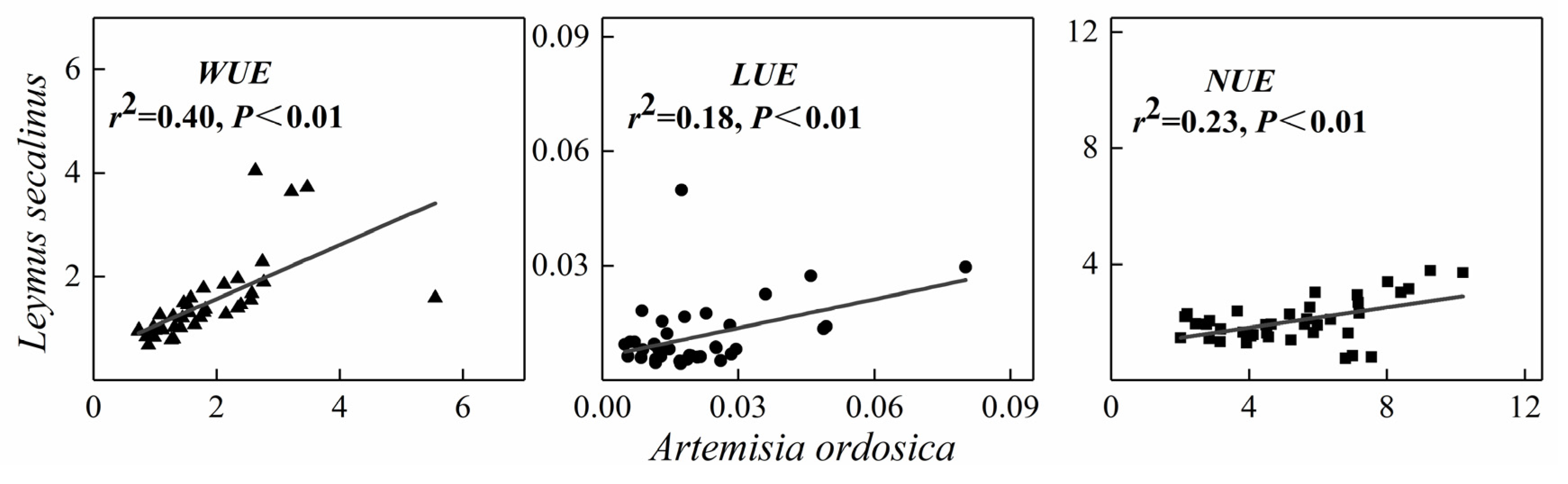
3.3. Relationships among RUEs for a Given Species and between Species
3.4. Controlling Factors on Variations in RUEs
4. Discussion
4.1. Variations in RUEs and Their Controlling Factors
4.2. Tradeoffs between RUEs
4.3. Relationships in RUEs between the Two Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full-Name | Abbreviation | Unit |
|---|---|---|
| Resource use efficiencies | RUEs | - |
| Water use efficiency | WUE | μmol·mmol−1 |
| Light use efficiency | LUE | mol·mol−1 |
| Nitrogen use efficiency | NUE | μmol·g−1·s−1 |
| Photosynthetic rate | Pn | μmol·m−2·s−1 |
| Transpiration rate | E | mmol·m−2·s−1 |
| Stomatal conductance | Gs | mol·m−2·s−1 |
| Specific leaf area | SLA | cm2·g−1 |
| Leaf nitrogen content per unit area | Narea | g·m−2 |
| Leaf nitrogen per unit dry mass | Nmass | g·kg−1 |
| Air temperature | T | °C |
| Relative humidity | RH | % |
| Net radiation | Rn | W·m−2 |
| Incident photosynthetically active radiation | PAR | μmol·m−2·s−1 |
| Soil water content at 30-cm depth | SWC | m3·m−3 |
| Vapor pressure deficient | VPD | kPa |
| Coefficient of variation | CV | % |
References
- Huang, J.; Yu, H.; Dai, A.; Wei, Y.; Kang, L. Drylands face potential threat under 2 °C global warming target. Nat. Clim. Chang. 2017, 7, 417–422. [Google Scholar] [CrossRef]
- Maestre, F.T.; Quero, J.L.; Gotelli, N.J.; Escudero, A.; Ochoa, V.; Delgado-Baquerizo, M.; García-Gómez, M.; Bowker, M.A.; Soliveres, S.; Escolar, C.; et al. Plant species richness and ecosystem multifunctionality in global drylands. Science 2012, 335, 214–218. [Google Scholar] [CrossRef] [Green Version]
- Stringer, L.C.; Reed, M.S.; Fleskens, L.; Thomas, R.J.; Le, Q.B.; Lala-Pritchard, T. A new dryland development paradigm grounded in empirical analysis of dryland systems science. Land Degrad. Dev. 2017, 28, 1952–1961. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.; Stafford-Smith, M.; Wang, Y.; Wu, B.; Yu, X.; Lv, N.; Ojima, D.S.; Lv, Y.; Fu, C.; Liu, Y.; et al. The global-DEP conceptual framework—Research on dryland ecosystems to promote sustainability. Curr. Opin. Environ. Sustain. 2021, 48, 17–28. [Google Scholar] [CrossRef]
- Leach, M.; Rockström, J.; Raskin, P.; Scoones, I.; Stirling, A.C.; Smith, A.; Thompson, J.; Millstone, E.; Ely, A.; Arond, E.; et al. Transforming innovation for sustainability. Ecol. Soc. 2012, 17, 11. [Google Scholar] [CrossRef] [Green Version]
- Ahlström, A.; Raupach, M.R.; Schurgers, G.; Smith, B.; Arneth, A.; Jung, M.; Reichstein, M.; Canadell, J.G.; Friedlingstein, P.; Jain, A.K.; et al. The dominant role of semiarid ecosystems in the trend and variability of the land CO2 sink. Science 2015, 348, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Berg, A.; Findell, K.; Lintner, B.; Giannini, A.; Seneviratne, S.I.; van den Hurk, B.; Lorenz, R.; Pitman, A.; Hagemann, S.; Meier, A.; et al. Land–atmosphere feedbacks amplify aridity increase over land under global warming. Nat. Clim. Chang. 2016, 6, 869–874. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2015, 6, 166–171. [Google Scholar] [CrossRef]
- Zha, T.; Qian, D.; Jia, X.; Bai, Y.; Tian, Y.; Bourque, C.P.A.; Ma, J.; Feng, W.; Wu, B.; Peltola, H. Soil moisture control of sap-flow response to biophysical factors in a desert-shrub species, Artemisia ordosica. Biogeosciences 2017, 14, 4533–4544. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.-H.; Zha, T.; Jia, X.; Wu, Y.; Wu, B.; Zhang, Y.; Guo, J.; Qin, S.; Chen, G.; Peltola, H. Leaf nitrogen is closely coupled to phenophases in a desert shrub ecosystem in China. J. Arid Environ. 2015, 122, 124–131. [Google Scholar] [CrossRef]
- Limousin, J.M.; Yepez, E.A.; McDowell, N.G.; Pockman, W.T.; Tjoelker, M. Convergence in resource use efficiency across trees with differing hydraulic strategies in response to ecosystem precipitation manipulation. Funct. Ecol. 2015, 29, 1125–1136. [Google Scholar] [CrossRef] [Green Version]
- Hyvonen, R.; Agren, G.I.; Linder, S.; Persson, T.; Cotrufo, M.F.; Ekblad, A.; Freeman, M.; Grelle, A.; Janssens, I.A.; Jarvis, P.G.; et al. The likely impact of elevated CO2, nitrogen deposition, increased temperature and management on carbon sequestration in temperate and boreal forest ecosystems: A literature review. New Phytol. 2007, 173, 463–480. [Google Scholar] [CrossRef]
- Han, J.; Chen, J.; Miao, Y.; Wan, S. Multiple Resource Use Efficiency (mRUE): A New Concept for Ecosystem Production. Sci. Rep. 2016, 6, 37453. [Google Scholar] [CrossRef] [Green Version]
- Silvertown, J.; Araya, Y.; Gowing, D.; Cornwell, W. Hydrological niches in terrestrial plant communities: A review. J. Ecol. 2015, 103, 93–108. [Google Scholar] [CrossRef] [Green Version]
- Gong, X.Y.; Chen, Q.; Lin, S.; Brueck, H.; Dittert, K.; Taube, F.; Schnyder, H. Tradeoffs between nitrogen- and water-use efficiency in dominant species of the semiarid steppe of Inner Mongolia. Plant Soil 2010, 340, 227–238. [Google Scholar] [CrossRef]
- Tarvainen, L.; Rantfors, M.; Wallin, G. Seasonal and within-canopy variation in shoot-scale resource-use efficiency trade-offs in a Norway spruce stand. Plant Cell Environ. 2015, 38, 2487–2496. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.E.; Adams, C.A.; Holsinger, K.E. Intraspecific variation in stomatal traits, leaf traits and physiology reflects adaptation along aridity gradients in a South African shrub. Ann. Bot. 2016, 117, 195–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvajal, D.E.; Loayza, A.P.; Rios, R.S.; Gianoli, E.; Squeo, F.A. Population variation in drought-resistance strategies in a desert shrub along an aridity gradient: Interplay between phenotypic plasticity and ecotypic differentiation. Perspect. Plant Ecol. 2017, 29, 12–19. [Google Scholar] [CrossRef]
- Jia, X.; Zha, T.; Gong, J.; Zhang, Y.; Wu, B.; Qin, S.; Peltola, H. Multi-scale dynamics and environmental controls on net ecosystem CO2 exchange over a temperate semiarid shrubland. Agric. For. Meteorol. 2018, 259, 250–259. [Google Scholar] [CrossRef]
- Hodapp, D.; Hillebrand, H.; Striebel, M. “Unifying” the Concept of Resource Use Efficiency in Ecology. Front. Ecol. Evol. 2019, 6, 233–247. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ramos, I.M.; Roumet, C.; Cruz, P.; Blanchard, A.; Autran, P.; Garnier, E.; Aerts, R. Evidence for a ‘plant community economics spectrum’ driven by nutrient and water limitations in a Mediterranean rangeland of southern France. J. Ecol. 2012, 100, 1315–1327. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Drought resistance, water-use efficiency, and yield potential—Are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159–1168. [Google Scholar] [CrossRef]
- Fischer, R.A.; Maurer, R. Drought resistance in spring wheat cultivars. I Grain yield responses. Aust. J. Agric. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- Lübbe, T.; Schuldt, B.; Leuschner, C. Acclimation of leaf water status and stem hydraulics to drought and tree neighbourhood: Alternative strategies among the saplings of five temperate deciduous tree species. Tree Physiol. 2017, 37, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Chen, J.; Shi, W.; Song, J.; Hui, D.; Ru, J.; Wan, S. Asymmetric responses of resource use efficiency to previous-year precipitation in a semi-arid grassland. Funct. Ecol. 2021, 35, 807–814. [Google Scholar] [CrossRef]
- Wu, Y.J.; Ren, C.; Tian, Y.; Zha, T.S.; Liu, P.; Bai, Y.J.; Ma, J.Y.; Lai, Z.R.; Bourque, C.P.A. Photosynthetic gas-exchange and PSII photochemical acclimation to drought in a native and non-native xerophytic species (Artemisia ordosica and Salix psammophila). Ecol. Indic. 2018, 94, 130–138. [Google Scholar] [CrossRef]
- Niu, S.; Xing, X.; Zhang, Z.H.E.; Xia, J.; Zhou, X.; Song, B.; Li, L.; Wan, S. Water-use efficiency in response to climate change: From leaf to ecosystem in a temperate steppe. Glob. Chang. Biol. 2011, 17, 1073–1082. [Google Scholar] [CrossRef]
- Xie, J.; Zha, T.; Zhou, C.; Jia, X.; Yu, H.; Yang, B.; Chen, J.; Zhang, F.; Wang, B.; Bourque, C.P.A.; et al. Seasonal variation in ecosystem water use efficiency in an urban-forest reserve affected by periodic drought. Agric. For. Meteorol. 2016, 221, 142–151. [Google Scholar] [CrossRef]
- Lübbe, T.; Schuldt, B.; Coners, H.; Leuschner, C. Species diversity and identity effects on the water consumption of tree sapling assemblages under ample and limited water supply. Oikos 2016, 125, 86–97. [Google Scholar] [CrossRef]
- Wei, H.; Luo, T.; Wu, B. Optimal balance of water use efficiency and leaf construction cost with a link to the drought threshold of the desert steppe ecotone in northern China. Ann. Bot. 2016, 118, 541–553. [Google Scholar] [CrossRef] [Green Version]
- Lai, Z.; Zhang, Y.; Liu, J.; Wu, B.; Qin, S.; Fa, K. Fine-root distribution, production, decomposition, and effect on soil organic carbon of three revegetation shrub species in northwest China. For. Ecol. Manag. 2016, 359, 381–388. [Google Scholar] [CrossRef]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development; General Assembly Resolution A/RES/70/1; United Nations: New York, NY, USA, 2015. [Google Scholar]
- She, W.; Bai, Y.; Zhang, Y.; Qin, S.; Liu, Z.; Wu, B. Plasticity in meristem allocation as an adaptive strategy of a desert shrub under contrasting environments. Front. Plant Sci. 2017, 8, 1933. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Poorter, H.; Jaureguiberry, P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; et al. Corrigendum to: New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 2016, 64, 715–716. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Wu, Y.J.; Zha, T.S.; Jia, X.; Tian, Y.; Bai, Y.J.; Bourque, C.P.-A.; Ma, J.Y.; Feng, W. Seasonal changes in photosynthetic energy utilization in a desert shrub (Artemisia ordosica Krasch.) during its different phenophases. Forests 2018, 9, 176. [Google Scholar] [CrossRef] [Green Version]
- Zha, T.-S.; Wu, Y.J.; Jia, X.; Zhang, M.Y.; Bai, Y.J.; Liu, P.; Ma, J.Y.; Bourque, C.P.-A.; Peltola, H. Diurnal response of effective quantum yield of PSII photochemistry to irradiance as an indicator of photosynthetic acclimation to stressed environments revealed in a xerophytic species. Ecol. Indic. 2017, 74, 191–197. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- De la Riva, E.G.; Tosto, A.; Pérez-Ramos, I.M.; Navarro-Fernández, C.M.; Olmo, M.; Anten, N.P.R.; Marañón, T.; Villar, R.; de Bello, F. A plant economics spectrum in Mediterranean forests along environmental gradients: Is there coordination among leaf, stem and root traits? J. Veg. Sci. 2016, 27, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, S.; Zha, T.; Jia, X.; Hayat, M.; Qian, D.; Bourque, C.P.A.; Tian, Y.; Bai, Y.; Liu, P.; Yang, R.; et al. Interannual variation in sap flow response in three xeric shrub species to periodic drought. Agric. For. Meteorol. 2021, 297, 108276. [Google Scholar] [CrossRef]
- Cowan, I.R. Regulation of Water Use in Relation to Carbon Gain in Higher Plants. In Physiological Plant Ecology II; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1982; Volume 12/B, pp. 589–613. [Google Scholar]
- Sangines de Carcer, P.; Vitasse, Y.; Penuelas, J.; Jassey, V.E.J.; Buttler, A.; Signarbieux, C. Vapor-pressure deficit and extreme climatic variables limit tree growth. Glob. Chang. Biol. 2018, 24, 1108–1122. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Jia, X.; Zha, T.; Bourque, C.P.A.; Tian, Y.; Bai, Y.; Liu, P.; Yang, R.; Li, C.; Li, C.; et al. Ecosystem water use efficiency in a young plantation in Northern China and its relationship to drought. Agric. For. Meteorol. 2019, 275, 1–10. [Google Scholar] [CrossRef]
- Chen, S.; Bai, Y.; Lin, G.; Huang, J.; Han, X. Isotopic carbon composition and related characters of dominant species along an environmental gradient in Inner Mongolia, China. J. Arid Environ. 2007, 71, 12–28. [Google Scholar] [CrossRef]
- Wittmer, M.; Auerswald, K.; Tungalag, R.; Bai, Y.F.; Schäufele, R.; Bai, C.H.; Schnyder, H. Carbon isotope discrimination of C3 vegetation in Central Asian Grassland as related to long-term and short-term precipitation patterns. Biogeosciences 2008, 5, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Hayat, M.; Zha, T.; Jia, X.; Iqbal, S.; Qian, D.; Bourque, C.P.A.; Khan, A.; Tian, Y.; Bai, Y.; Liu, P.; et al. A multiple-temporal scale analysis of biophysical control of sap flow in Salix psammophila growing in a semiarid shrubland ecosystem of northwest China. Agric. For. Meteorol. 2020, 288–289, 107985. [Google Scholar] [CrossRef]
- Chasmer, L.; Mccaughey, H.; Barr, A.; Black, A.; Shashkov, A.; Treitz, P.; Zha, T. Investigating light-use efficiency across a jack pine chronosequence during dry and wet years. Tree Physiol. 2008, 28, 1395–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbulsky, M.F.; Peñuelas, J.; Papale, D.; Ardö, J.; Goulden, M.L.; Kiely, G.; Richardson, A.D.; Rotenberg, E.; Veenendaal, E.M.; Filella, I. Patterns and controls of the variability of radiation use efficiency and primary productivity across terrestrial ecosystems. Glob. Ecol. Biogeogr. 2010, 19, 253–267. [Google Scholar] [CrossRef]
- Stocker, B.D.; Zscheischler, J.; Keenan, T.F.; Prentice, I.C.; Penuelas, J.; Seneviratne, S.I. Quantifying soil moisture impacts on light use efficiency across biomes. New Phytol. 2018, 218, 1430–1449. [Google Scholar] [CrossRef] [Green Version]
- Lü, X.T.; Kong, D.L.; Pan, Q.M.; Simmons, M.E.; Han, X.G. Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grassland. Oecologia 2012, 168, 301–310. [Google Scholar] [CrossRef]
- Bai, Y.; Zha, T.; Bourque, C.P.A.; Jia, X.; Ma, J.; Liu, P.; Yang, R.; Li, C.; Du, T.; Wu, Y. Variation in ecosystem water use efficiency along a southwest-to-northeast aridity gradient in China. Ecol. Indic. 2020, 110, 105932. [Google Scholar] [CrossRef]
- De Bello, F.; Lavorel, S.; Albert, C.H.; Thuiller, W.; Grigulis, K.; Dolezal, J.; Janeček, Š.; Lepš, J. Quantifying the relevance of intraspecific trait variability for functional diversity. Methods Ecol. Evol. 2011, 2, 163–174. [Google Scholar] [CrossRef]
- Liu, P.; Black, T.A.; Jassal, R.S.; Zha, T.; Nesic, Z.; Barr, A.G.; Helgason, W.D.; Jia, X.; Tian, Y.; Stephens, J.J.; et al. Divergent long-term trends and interannual variation in ecosystem resource use efficiencies of a southern boreal old black spruce forest 1999–2017. Glob. Chang. Biol. 2019, 25, 3056–3069. [Google Scholar] [CrossRef]
- Bloom, A.J.; Chapin, F.S., III; Mooney, H.A. Resource limitation in plants-an economic analogy. Ann. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Tarvainen, L.; Rantfors, M.; Wallin, G. Vertical gradients and seasonal variation in stem CO2 efflux within a Norway spruce stand. Tree Physiol. 2014, 34, 488–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auzmendi, I.; Marsal, J.; Girona, J.; Lopez, G. Daily photosynthetic radiation use efficiency for apple and pear leaves: Seasonal changes and estimation of canopy net carbon exchange rate. Eur. J. Agron. 2013, 51, 1–8. [Google Scholar] [CrossRef]
- Patterson, T.B.; Guy, R.D.; Dang, Q.L. Whole-plant nitrogen- and water-relations traits, and their associated tradeoffs, in adjacent muskeg and upland boreal spruce species. Oecologia 1997, 110, 160–168. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Tenhunen, J.D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant Cell Environ. 1997, 20, 845–866. [Google Scholar] [CrossRef]
- Niinemets, U.; Keenan, T.F.; Hallik, L. A worldwide analysis of within-canopy variations in leaf structural, chemical and physiological traits across plant functional types. New Phytol. 2015, 205, 973–993. [Google Scholar] [CrossRef] [Green Version]
- Carvajal, D.E.; Loayza, A.P.; Rios, R.S.; Delpiano, C.A.; Squeo, F.A.; Cahill, J. A hyper-arid environment shapes an inverse pattern of the fast–slow plant economics spectrum for above-, but not below-ground resource acquisition strategies. J. Ecol. 2018, 107, 1079–1092. [Google Scholar] [CrossRef]
- Liu, G.; Freschet, G.T.; Pan, X.; Cornelissen, J.H.; Li, Y.; Dong, M. Coordinated variation in leaf and root traits across multiple spatial scales in Chinese semi-arid and arid ecosystems. New Phytol. 2010, 188, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Sperdouli, I.; Moustakas, M. Spatio-temporal heterogeneity in Arabidopsis thaliana leaves under drought stress. Plant Biol. 2012, 14, 118–128. [Google Scholar] [CrossRef]
- Brouillette, L.C.; Mason, C.M.; Shirk, R.Y.; Donovan, L.A. Adaptive differentiation of traits related to resource use in a desert annual along a resource gradient. New Phytol. 2014, 201, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Ivey, C.T.; Carr, D.E. Tests for the joint evolution of mating system and drought escape in Mimulus. Ann. Bot. 2012, 109, 583–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefcheck, J.S.; Byrnes, J.E.K.; Isbell, F.; Gamfeldt, L.; Griffin, J.N.; Eisenhauer, N.; Hensel, M.J.S.; Hector, A.; Cardinale, B.J.; Duffy, J.E. Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats. Nat. Commun. 2015, 6, 6936. [Google Scholar] [CrossRef] [PubMed]









| RUEs | Model | R2 | F | p | |
|---|---|---|---|---|---|
| Artemisia ordosica | WUE | y = −1.40VPD–0.71Gs + 3.50 | 0.24 | 7.55 (1,40) | 0.002 |
| LUE | y = 0.42SWC + 0.08Gs + 0.03 | 0.32 | 10.46 (1,40) | <0.001 | |
| NUE | Y = 8.99Gs + 2.58 | 0.56 | 53.86 (1,40) | <0.001 | |
| Leymus secalinus | WUE | y = −0.01SLA − 2.98Gs – 1.25VPD + 4.19 | 0.37 | 8.38 (1,37) | <0.001 |
| LUE | y = 0.24SWC + 0.16Gs + 1.74 | 0.41 | 17.66 (1,37) | <0.001 | |
| NUE | y = 0.02SLA + 3.88Gs + 0.06 | 0.36 | 11.46 (1,37) | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Tian, Y.; Zha, T.; Jia, X.; Bourque, C.P.-A.; Liu, P.; Jin, C.; Jiang, X.; Li, X.; Wei, N.; et al. Dynamic Changes in Plant Resource Use Efficiencies and Their Primary Influence Mechanisms in a Typical Desert Shrub Community. Forests 2021, 12, 1372. https://0-doi-org.brum.beds.ac.uk/10.3390/f12101372
Jiang Y, Tian Y, Zha T, Jia X, Bourque CP-A, Liu P, Jin C, Jiang X, Li X, Wei N, et al. Dynamic Changes in Plant Resource Use Efficiencies and Their Primary Influence Mechanisms in a Typical Desert Shrub Community. Forests. 2021; 12(10):1372. https://0-doi-org.brum.beds.ac.uk/10.3390/f12101372
Chicago/Turabian StyleJiang, Yan, Yun Tian, Tianshan Zha, Xin Jia, Charles P.-A. Bourque, Peng Liu, Chuan Jin, Xiaoyan Jiang, Xinhao Li, Ningning Wei, and et al. 2021. "Dynamic Changes in Plant Resource Use Efficiencies and Their Primary Influence Mechanisms in a Typical Desert Shrub Community" Forests 12, no. 10: 1372. https://0-doi-org.brum.beds.ac.uk/10.3390/f12101372