Do Sandy Grasslands along the Danube in the Carpathian Basin Preserve the Memory of Forest-Steppes?

Abstract
:1. Introduction
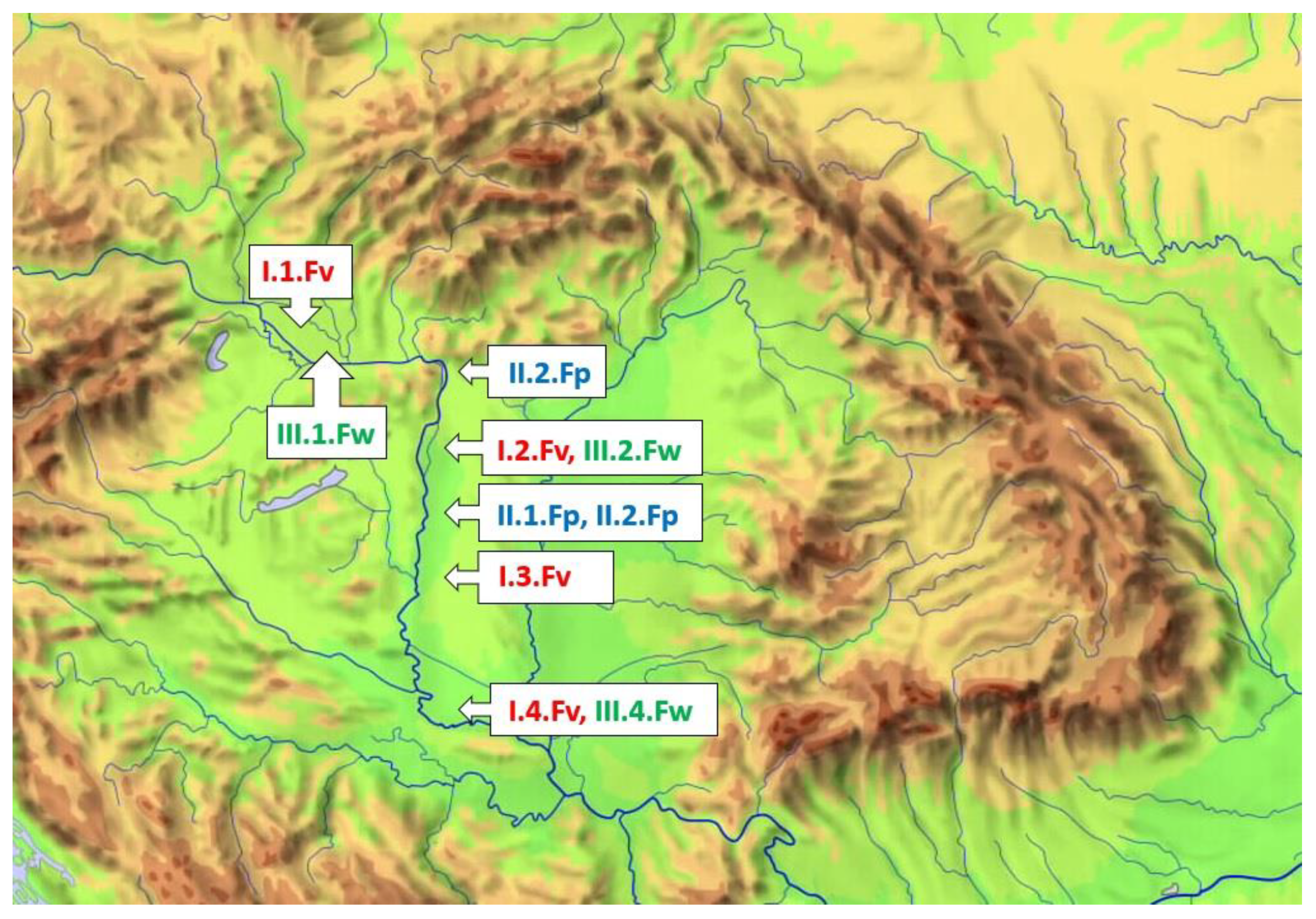
2. Materials and Methods
- Eu-oceanic species, occurs occasionally in Central Europe (not in Hungary);
- Oceanic species, occurs mainly in Western end Western Central Europe;
- Oceanic/suboceanic species, occurs mainly in Central Europe;
- Suboceanic species, occurs mainly in Central Europe and occasionally in Eastern Europe;
- Transitory types, with a slight suboceanic and subcontinental feature;
- Subcontinental species, occurs mainly in Central and Eastern Europe;
- Continental-subcontinental species, occurs mainly in Eastern Europe;
- Continental species, occurs occasionally in Central Europe;
- Eu-continental species, occurs mainly in Siberia and Eastern Europe (not in Central Europe).
3. Results
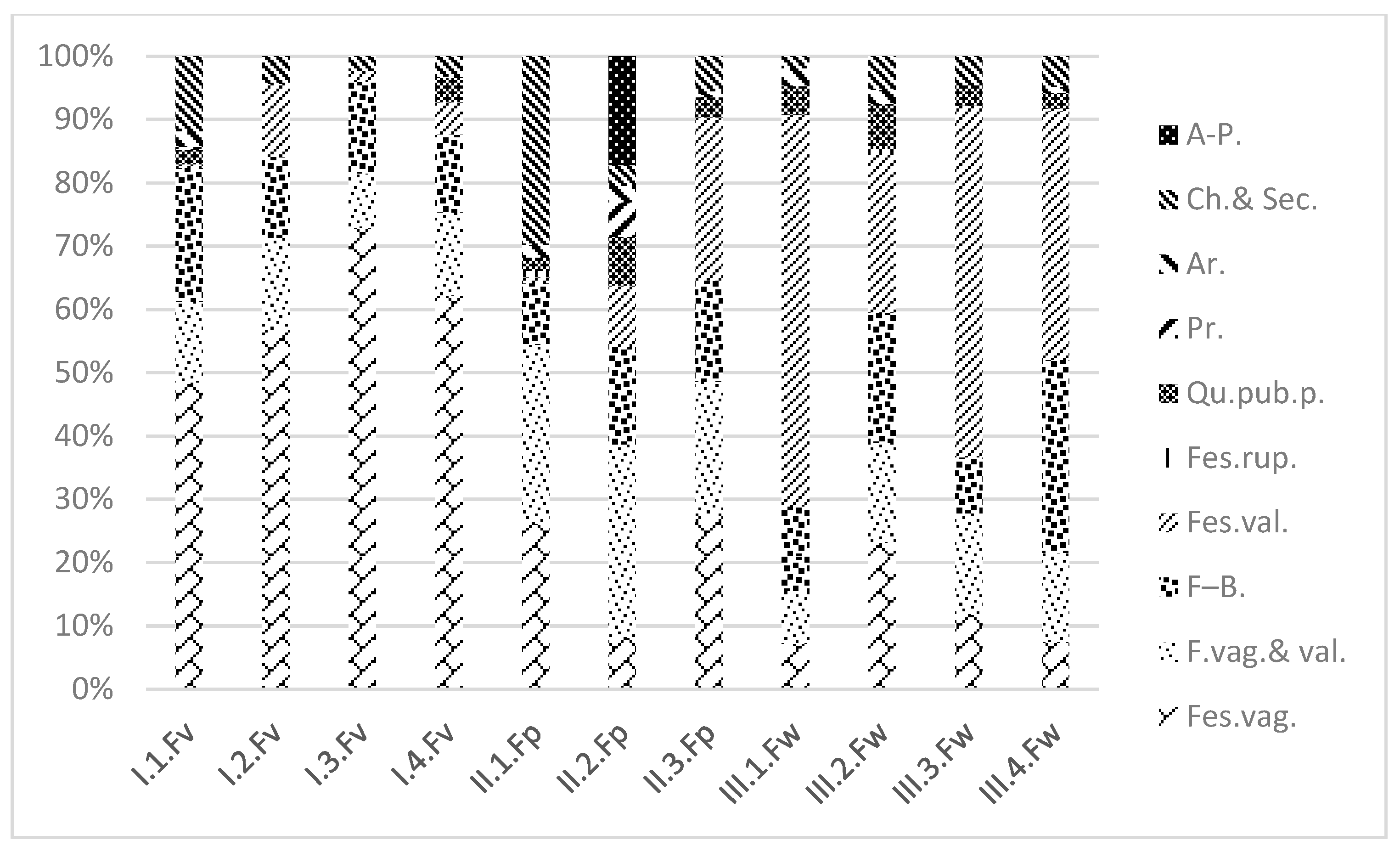
3.1. Coenosystematic Results
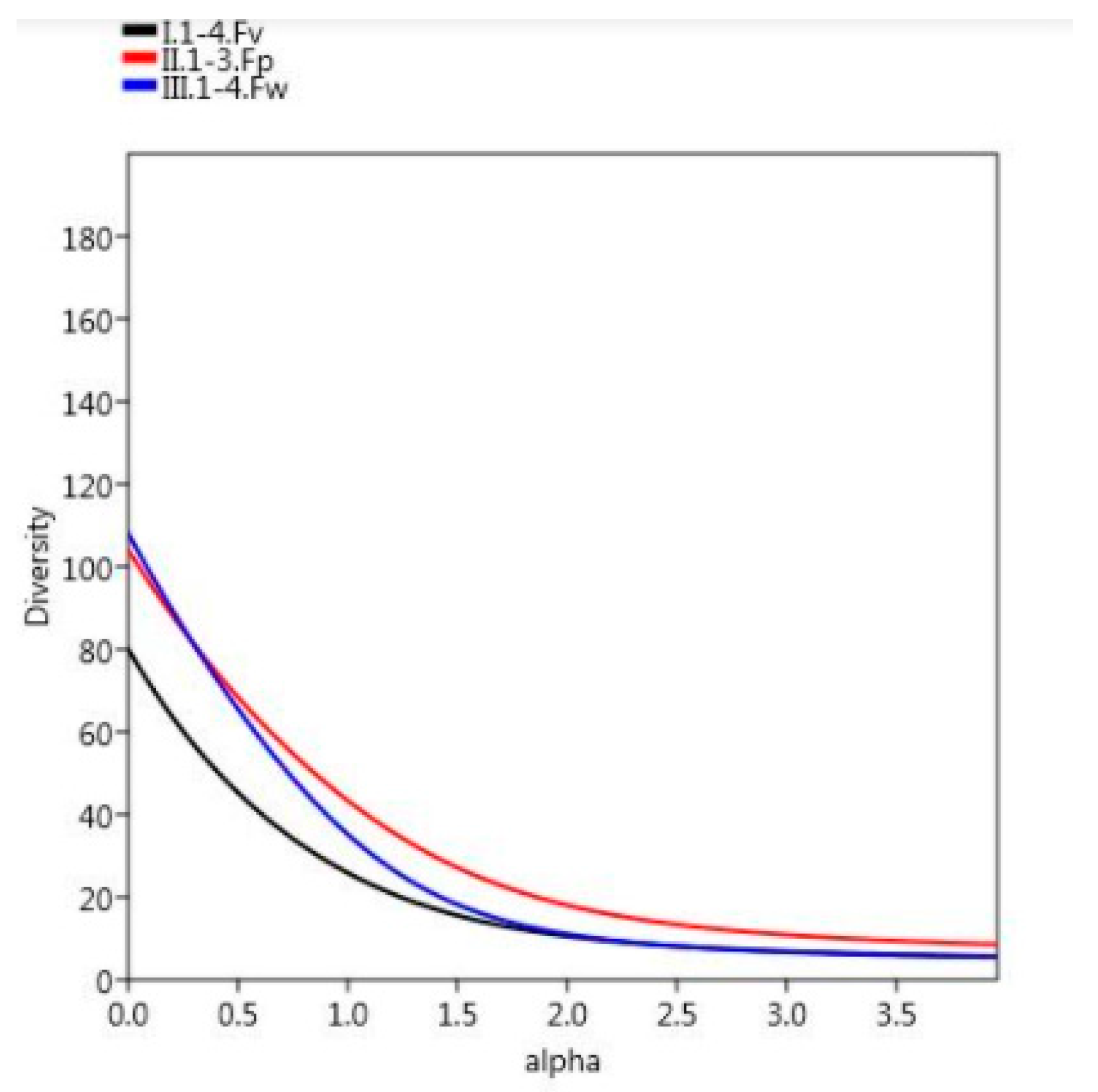
3.2. Diversity Results
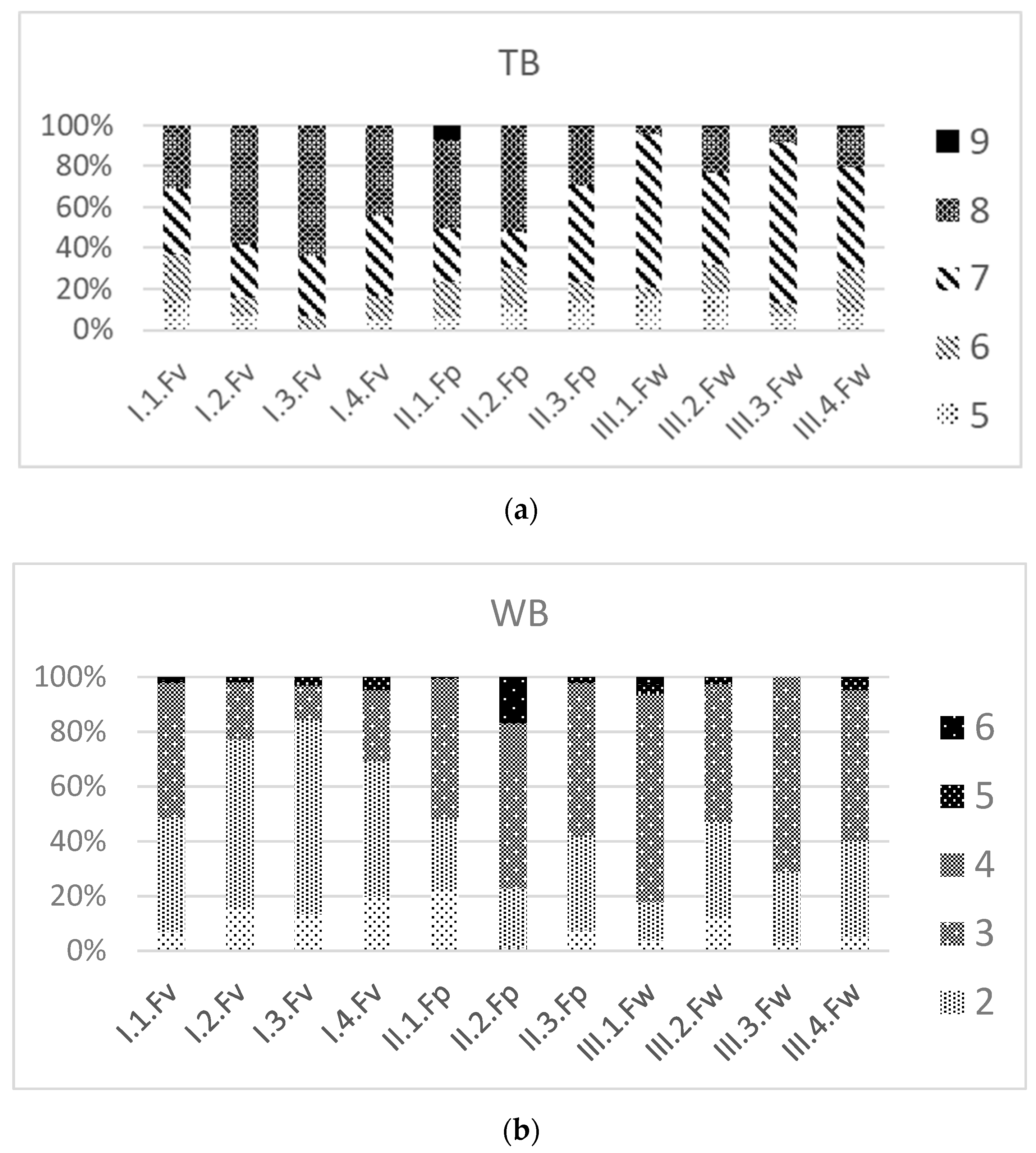
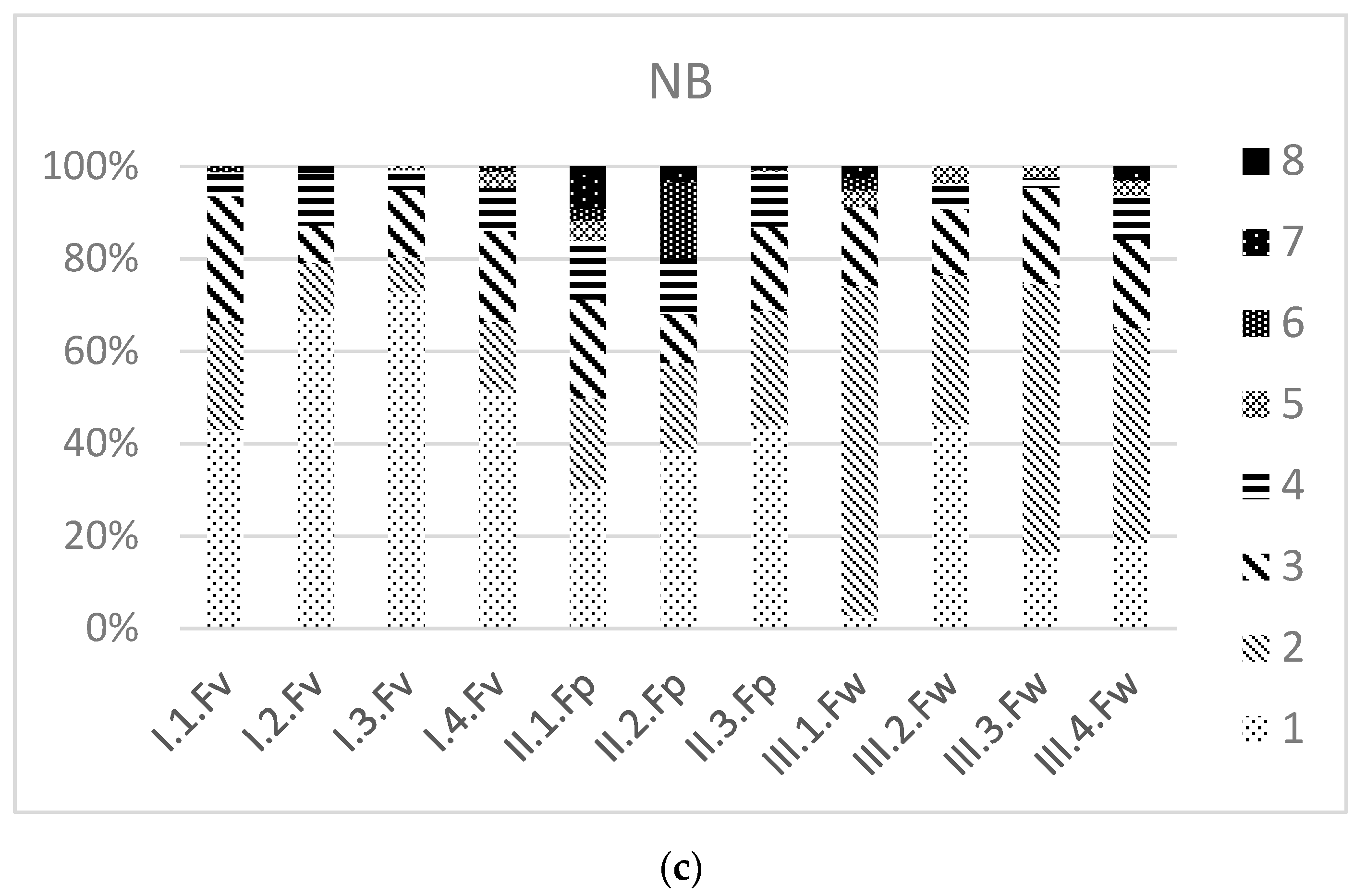
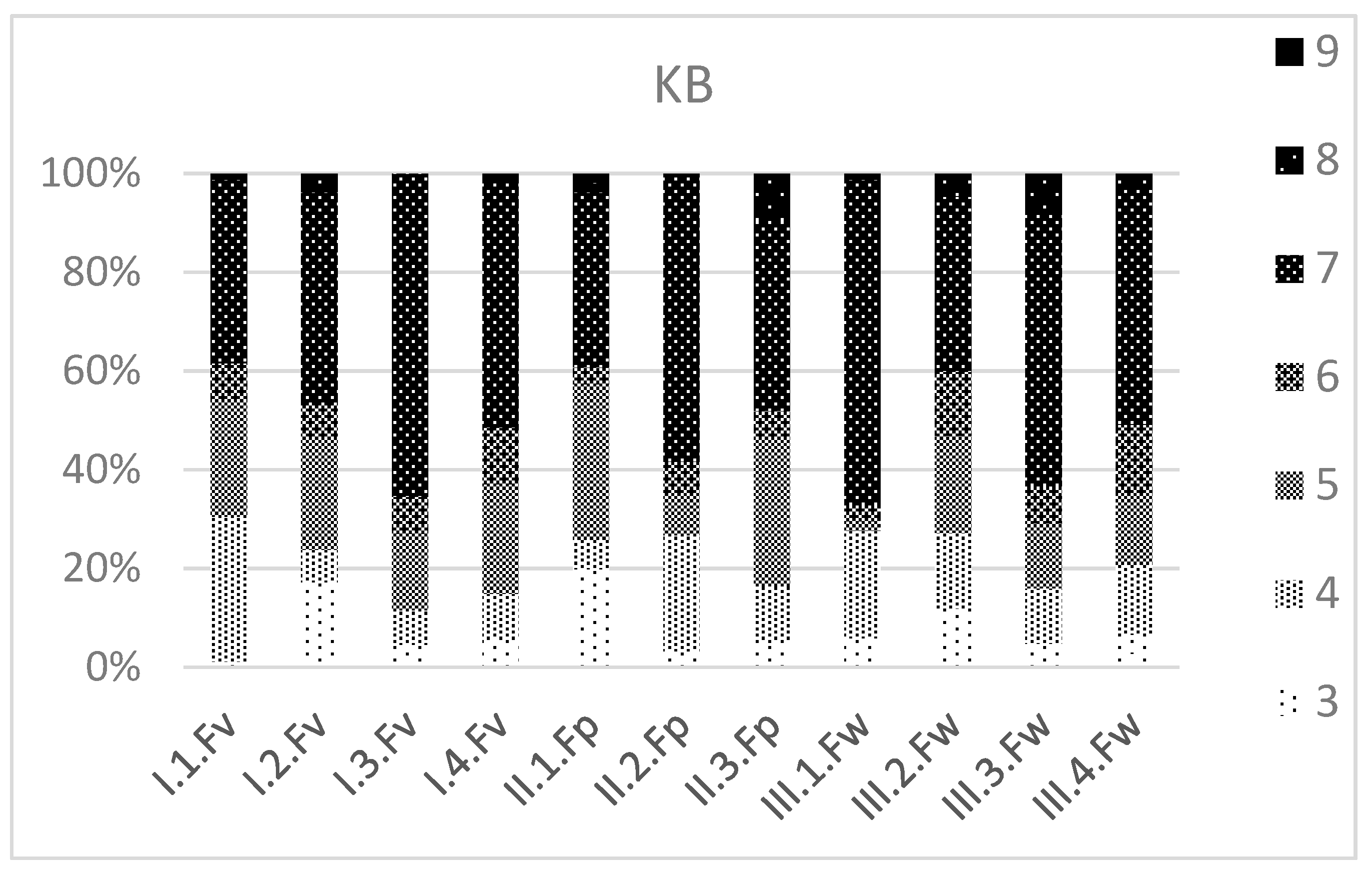
3.3. Ecological Values
3.4. Pedological Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Török, P.; Janišová, M.; Kuzemko, A.; Rūsiņa, A.; Stevanović, Z.D. Grassland of the World: Management and Conservation Grasslands, their Threats and Management in Eastern Europe. In Grasslands of the World: Diversity Management Conservation; Squires, V.R., Dengler, J., Hua, L., Feng, H., Eds.; CRC Press: London, UK, 2018; pp. 64–88. [Google Scholar]
- Török, P.; Wesche, K.; Ambarli, D.; Kamp, J.; Dengler, J. Step(pe) up! Raising the profile of the Palaearctic natural grasslands. Biodivers. Conserv. 2016, 25, 2187–2195. [Google Scholar] [CrossRef] [Green Version]
- Wesche, K.; Ambarlı, D.; Kamp, J.; Török, P.; Treiber, J.; Dengler, J. The Palaearctic steppe biome: A new synthesis. Biodivers. Conserv. 2016, 25, 2197–2231. [Google Scholar] [CrossRef]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Tichý, L. Vegetation of Europe: Hierarchical floristic classification system of plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. 1), 1–264. [Google Scholar] [CrossRef]
- Borhidi, A.; Kevey, B.; Lendvai, G.; Seregélyes, T. Plant Communities of Hungary; Akadémiai Kiadó: Budapest, Hungary, 2012. [Google Scholar]
- Molnár, Z.; Biró, M.; Bartha, S.; Fekete, G. Past trends, present state and future prospects of Hungarian forest-steppes. In Eurasian Steppes. Ecological Problems and Livelihoods in a Changing World; Werger, M.J.A., van Staalduinen, M.A., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2012; pp. 209–252. [Google Scholar]
- Varga, Z. Die Waldteppen des pannonischen Raumes aus biogeographischer Sicht. Düsseldorfer Geobot. Kolloqu. 1989, 6, 35–50. [Google Scholar]
- Tölgyesi, C.; Körmöczi, L. Structural changes of a Pannonian grassland plant community in relation to the decrease of water availability. Acta Bot. Hung. 2012, 54, 413–431. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Bátori, Z.; Erdős, L. Using statistical tests on relative ecological indicator values to compare vegetation units–Different approaches and weighting methods. Ecol. Indic. 2014, 36, 441–446. [Google Scholar] [CrossRef]
- Marosi, S.; Somogyi, S. Magyarország kistájainak katasztere I-II; MTA Földrajztudományi Kutató Intézet: Budapest, Hungary, 1990; 1023p. [Google Scholar]
- Tölgyesi, C.; Török, P.; Kun, R.; Csathó, A.I.; Bátori, Z.; Erdős, L.; Vadász, C. Recovery of species richness lags behind functional recovery in restored grasslands. Land Degrad. Dev. 2019, 30, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- Bartha, S.; Campetella, G.; Ruprecht, E.; Kun, A.; Házi, J.; Horváth, A.; Virágh, K.; Molnár, Z. Will inter-annual variability in sand grassland communities increase with climate change? Community Ecol. 2008, 9, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Bartha, S.; Molnár, Z.; Fekete, G. Patch dynamics in sand grasslands: Connecting primary and secondary succession. In The KISKUN LTER, Long-Term Ecological Research in the Kiskunság; Kovács-Láng, E., Molnár, E., Kröel-Dulay, G., Barabás, S., Eds.; Institute of Ecology and Botany: Vácrátót, Hungary, 2008; Volume 37–40. [Google Scholar]
- Bátori, Z.; Lengyel, A.; Maróti, K.; Körmöczi, L.; Tölgyesi, C.; Bíró, A.; Tóth, M.; Kincses, Z.; Cseh, V.; Erdős, L. Microclimate-vegetation relationships in natural habitat islands: Species preservation and conservation perspectives. Időjárás Q. J. Hung. Meteorol. Serv. 2014, 118, 257–281. [Google Scholar]
- Tölgyesi, C.; Valkó, O.; Deák, B.; Kelemen, A.; Bragina, T.M.; Gallé, R.; Erdős, L.; Bátori, Z. Tree–herb co-existence and community assembly in natural forest-steppe transitions. Plant Ecol. Divers. 2018, 11, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Blaser, W.J.; Sitters, J.; Hart, S.P.; Edwards, P.J.; Venterink, H.O. Facilitative or competitive effects of woody plants on understorey vegetation depend on N-fixation, canopy shape and rainfall. J. Ecol. 2013, 101, 1598–1603. [Google Scholar] [CrossRef] [Green Version]
- Bertness, M.; Callaway, R.M. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Collins, S.L.; Knapp, A.K.; Briggs, J.M.; Blair, J.M.; Steinauer, E.M. Modulation of diversity by grazing and mowing in native tallgrass prairie. Science 1998, 280, 745–747. [Google Scholar] [CrossRef] [PubMed]
- Güsewell, S.; Le Nédic, C. Effects of winter mowing on vegetation succession in a lakeshore fen. Appl. Veg. Sci. 2004, 7, 41–48. [Google Scholar] [CrossRef]
- Csecserits, A.; Rédei, T. Secondary succession on sandy old fields in Hungary. Appl. Veg. Sci. 2001, 4, 63–74. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Bátori, Z.; Erdős, L.; Gallé, R.; Körmöczi, L. Plant diversity patterns of a Hungarian steppe-wetland mosaic in relation to grazing regime and land use history (Muster der Phytodiversität in ungarischen Steppen-Feuchtwiesen Mosaiken in Abhängigkeit von der Beweidungsintensität und Landnutzungsgeschichte). Tuexenia 2015, 35, 399–416. [Google Scholar]
- Erdős, L.; Tölgyesi, C.; Bátori, Z.; Semenishchenkov, A.; Yu, A.; Magnes, M. The influence of forest/grassland proportion on the species composition, diversity and natural values of an Eastern Austrian forest-steppe. Russ. J. Ecol. 2017, 48, 350–357. [Google Scholar] [CrossRef] [Green Version]
- Erdős, L.; Török, P.; Szitár, K.; Bátori, Z.; Tölgyesi, C.; Kiss, P.J.; Bede-Fazekas, Á.; Kröel-Dulay, G. Beyond the forest-grassland dichotomy: The gradient-like organization of habitats in forest-steppes. Front. Plant Sci. 2020, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Erdős, L.; Kröel-Dulay, G.; Bátori, Z.; Kovács, B.; Németh, C.; Kiss, P.J.; Tölgyesi, C. Habitat heterogeneity as a key to high conservation value in forest-grassland mosaics. Biol. Conserv. 2018, 226, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Erdős, L.; Tölgyesi, C.; Horzse, M.; Tolnay, D.; Hurton, Á.; Schulcz, N.; Körmöczi, L.; Lengyel, A.; Bátori, Z. Habitat complexity of the Pannonian forest-steppe zone and its nature conservation implications. Ecol. Complex. 2014, 17, 107–118. [Google Scholar] [CrossRef]
- Biró, M.; Révész, A.; Molnár, Z.; Horváth, F.; Czúcz, B. Regional habitat pattern of the Danube-Tisza Interfluve in Hungary II. Acta Biol. Hung. 2008, 50, 19–60. [Google Scholar] [CrossRef]
- Czúcz, B.; Révész, A.; Horváth, F.; Bíró, M. Loss of seminatural grasslands in the Hungarian forest steppe zone in the last fifteen years: Causes and fragmentation patterns. In Proceedings of the 13th Annual IALE (UK) Conference: Planning, People and Practice: The Landscape Ecology of Sustainable Landscapes, Northampton, UK, 12 September 2005; McCollinn, D., Jackson, J.I., Eds.; University of Northampton: Northampton, UK, 2005; pp. 73–80. [Google Scholar]
- Csecserits, A.; Czúcz, B.; Halassy, M.; Kröel-Dulay, G.; Rédei, T.; Szabó, R.; Szitár, K.; Török, K. Regeneration of sandy old fields in the forest steppe region of Hungary. Plant Biosyst. 2011, 145, 715–729. [Google Scholar] [CrossRef]
- Bartha, S.; Zimmermann, Z.; Horváth, A.; Szentes, S.; Sutyinszki, Z.; Szabó, G.; Házi, J.; Komoly, C.; Penksza, K. High resolution vegetation assessment with beta-diversity—A moving window approach. Agric. Inform. 2011, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Albert, Á.-J.; Kelemen, A.; Valkó, O.; Miglécz, T.; Csecserits, A.; Rédei, T.; Deák, B.; Tóthmérész, B.; Török, P. Trait-based analysis of spontaneous grassland recovery in sandy old-fields. Appl. Veg. Sci. 2014, 17, 214–224. [Google Scholar] [CrossRef]
- Pándi, I.; Penksza, K.; Botta-Dukát, Z.; Kröel-Dulay, G. People move but cultivated plants stay: Abandoned farmsteads support the persistence and spread of alien plants. Biodivers. Conserv. 2014, 23, 1289–1302. [Google Scholar] [CrossRef]
- Török, P.; Kelemen, A.; Valkó, O.; Deák, B.; Lukács, B.; Tóthmérész, B. Lucerne-dominated fields recover native grass diversity without intensive management actions. J. Appl. Ecol. 2011, 48, 257–264. [Google Scholar] [CrossRef]
- Török, P.; Deák, B.; Vida, E.; Valkó, O.; Lengyel, S.; Tóthmérész, B. Restoring grassland biodiversity: Sowing low-diversity seed mixtures can lead to rapid favourable changes. Biol. Conserv. 2010, 143, 806–812. [Google Scholar] [CrossRef]
- Prach, K.; Pyšek, P.; Šmilauer, P. Prediction of vegetation succession in human-disturbed habitats using an expert system. Restor. Ecol. 1999, 7, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Prach, K.; Pyšek, P.; Bastl, M. Spontaneous vegetation succession in human-disturbed habitats: A pattern across seres. Appl. Veg. Sci. 2001, 4, 83–88. [Google Scholar] [CrossRef]
- Prach, K.; Bartha, S.; Joyce, C.B.; Pyšek, P.; van Diggelen, R.; Wiegleb, G. The role of spontaneous vegetation succession in ecosystem restoration: A perspective. Appl. Veg. Sci. 2001, 4, 111–114. [Google Scholar] [CrossRef]
- Szodfridt, I. Termõhelytípusok és vegetáció kapcsolata a Duna-Tisza közi homokháton. [Correlation of site conditions and vegetation in the Duna-Tisza köze-Sand plants as indicators for the sand afforestation]. Abstr. Bot. 1974, 2, 35–37. [Google Scholar]
- Szodfridt, I. Talajvíz és vegetáció kapcsolata a Duna-Tisza köze homokterületén. [Correlation of the soil water table and the vegetation]. Abstr. Bot. 1974, 2, 39–42. [Google Scholar]
- Szodfridt, I. A Duna-Tisza közi homokhátság növénytársulásainak fatermõképessége. [Timber growing capacity of the woodlands in Duna-Tisza köze]. Erdészettudományi Közlemények 1989, 10, 99–105. [Google Scholar]
- Járó, Z. A Duna-Tisza-közi homokhát termőhely típusai. Abstr. Bot. 1974, 2, 31–34. [Google Scholar]
- Dövényi, Z. Magyarország Kistájainak Katasztere; MTA Földrajztudományi Kutatóintézet: Budapest, Hungary, 2010; 876p. [Google Scholar]
- Stefanovits, P. A tájak talajviszonyai. In Talajtan; Stefanovits, P., Filep, G., Füleki, G., Eds.; Mezőgazda Kiadó: Budapest, Hungary, 2010. [Google Scholar]
- Barczi, A.; Joó, K. The role of Kurgans in the palaeopedological and palaeoecological reconstruction of the Hungarian Great Plain. Z. Geomorphol. 2009, 53, 131–137. [Google Scholar] [CrossRef]
- Barczi, A.; Joó, K.; Pető, Á.; Bucsi, T. Survey of the buried palaeosol under Lyukas–mound. Eurasian Soil Sci. 2006, 39, 133–140. [Google Scholar] [CrossRef]
- Fekete, G.; Molnár, Z.; Kun, A.; Botta-Dukát, Z. On the structure of the Pannonian forest steppe: Grasslands on sand. Acta Zool. Acad. Sci. Hung. 2002, 48, 137–150. [Google Scholar]
- Kun, A. Analysis of precipitation year types and their regional frequency distributions in the Danube-Tisza mid-region, Hungary. Acta Bot. Hung. 2002, 43, 175–187. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Zalatnai, M.; Erdős, L.; Bátori, Z.; Hupp, N.R.; Körmöczi, L. Unexpected ecotone dynamics of a sand dune vegetation complex following water table decline. J. Plant Ecol. 2016, 9, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Pawlus, M. Systematyka i rozmieszczenie gatunków grupy Festuca ovina L. w. Polsce [Classification and distribution of Festuca ovina L. in Poland]. Fragm. Flor. Geobot. 2005, 29, 219–295. [Google Scholar]
- Penksza, K. Kiegészítések a hazai Festuca taxonok ismeretéhez I. A Festuca psammophila series Festuca vaginata alakkörei [Additions to the knowledge of Hungarian Festuca taxa I. Taxa of Festuca psammophila series in the Festuca vaginata species complex]. Bot. Közlemények 2019, 106, 65–70. [Google Scholar] [CrossRef]
- Penksza, K.; Szabó, G.; Zimmermann, Z.; Lisztes-Szabó, Z.; Pápay, G.; Járdi, I.; Fűrész, A.; Falusi, S.-E. A Festuca vaginata alakkör taxonómiai problematikája és ennek cönoszisztematikai vonatkozásai [The taxonomic problems of the Festuca vaginata agg. and their coenosystematic aspects]. Georg. Agric. 2019, 23, 63–76. [Google Scholar]
- Korneck, D.; Gregor, T. Festuca tomanii sp. nov., ein Dünen-Schwingel des nördlichen oberrhein-, des mittleren main- und des böhmischen Elbetales. Kochia 2015, 9, 37–58. [Google Scholar]
- Bajor, Z.; Zimmermann, Z.; Szabó, G.; Fehér, Z.; Járdi, I.; Lampert, R.; Kerényi-Nagy, V.; Penksza, P.L.; Szabó, Z.; Székely, Z.; et al. Effect of conservation management practices on sand grassland vegetation in Budapest, Hungary. Appl. Ecol. Environ. Res. 2016, 14, 233–247. [Google Scholar] [CrossRef]
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian flora. Acta Bot. Acad. Sci. Hung. 1995, 39, 97–181. [Google Scholar]
- Király, G. Új Magyar Füvészkönyv. Magyarország Hajtásos Növényei (New Hungarian Herbal. The Vascular Plants of Hungary. Identification Key); Aggteleki Nemzeti Park: Jósvafő, Hungary, 2009. (In Hungarian) [Google Scholar]
- Ellenberg, H. Zeigerwerte der Gefässpflanzen Mitteleuropas. Scr. Geobot. 1974, 9, 1–97. [Google Scholar]
- Meusel, H.; Schubert, R. Volk. und Wissen; Akademie Verlag: Berlin, Germany, 1972. [Google Scholar]
- FAO. Guidelines for Soil Description, 4th ed.; FAO: Rome, Italy, 2006. [Google Scholar]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014 (Updated 2015); World Soil Resources Reports 106; Food and Agriculture Organization (FAO) of United Nations: Rome, Italy, 2016. [Google Scholar]
- Buzás, I. Soil and Agrochemistry Analysing Method Book No. 1. Physical, Hydrological and Mineralogy Analyses of Soils; INDA 4231 Publishing: Budapest, Hungary, 2013; p. 357. (In Hungarian) [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. Methods of Soil Analysis. Part 3. Chemical Methods. Soil Sci. Soc. Am. Book Ser. 1996, 5, 961–1010. [Google Scholar]
- Hammer, Ø. PAST–PAleontological STatictics Version 3.06 Reference Manual; Natural History Museum, University of Oslo: Oslo, Norway, 2015; 225p. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST–Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1994, 4, 406–425. [Google Scholar]
- Tóthmérész, B. Comparison of different methods for diversity ordering. J. Veg. Sci. 1995, 6, 283–290. [Google Scholar] [CrossRef]
- Bátori, Z.; Erdős, L.; Kelemen, A.; Deák, B.; Valkó, O.; Gallé, R.; Bragina, T.M.; Kiss, P.J.; Kröel-Dulay, G.; Tölgyesi, C. Diversity patterns in sandy forest-steppes: A comparative study from the western and central Palearctic. Biodivers Conserv. 2018, 27, 1011–1030. [Google Scholar] [CrossRef]
- Csontos, P.; Bózsing, E.; Cseresnyés, I.; Penksza, K. Reproductive potential of Asclepias syriaca stands in the rural surroundings of Budapest, Hungary. Pol. J. Ecol. 2009, 57, 383–388. [Google Scholar]
- Penksza, K.; Péter, N.; Saláta, D.; Pápay, G.; Lisztes-Szabó, Z.; Bajor, Z. Result of conservation management and restauration in open sandy grasslands in the Homoktövis Nature Conversation area (Budapest, Hungary). In Proceedings of the International Conference on Veterinary, Agriculture and Life Sciences (ICVALS), Antalya, Turkey, 29 October–1 November 2020, Abstract Book; Mehmet, O., Ed.; ISRES Publishing: Antalya, Turkey, 2020; p. 7. [Google Scholar]
- Pócs, T. A rákoskereszturi “Akadémiai erdö” vegetációja. [Die Vegetation des “Akademischen Waldes” in Rákoskeresztur]. Bot. Közlemények 1954, 45, 283–294. [Google Scholar]
- Penksza, K.; Engloner, A. Taxonomic study of Festuca wagneri (Degen Thaisz et Flatt) in Degen Thaisz et Flatt. 1905. Acta Bot. Sci. Hung. 2000, 42, 257–264. [Google Scholar]
- Penksza, K. Festuca pseudovaginata, a new species from sandy areas of the carpathian basin. Acta Bot. Hung. 2003, 45, 365–372. [Google Scholar] [CrossRef]
- Penksza, K.; Csontos, P.; Pápay, G. Festucetum pseudovaginatae ass. nova is a new indigenous association in the Pannonian basin. Haquetia 2020, in press. [Google Scholar]
- Szabó, G.; Zimmermann, Z.; Catorci, A.; Csontos, P.; Wichmann, B.; Szentes, S.; Barczi, A.; Penksza, K. Comparative study on grasslands dominated by Festuca vaginata and F. pseudovaginata in the Carpathian Basin. Tuexenia 2017, 37, 415–429. [Google Scholar]
- Šmarda, P.; Šmerda, J.; Knoll, A.; Bureš, P.; Danihelka, J. Revision of Central European taxa of Festuca ser. Psammophilae Pawlus: Morphometrical, karyological and AFLP analysis. Plant Syst. Evol. 2007, 266, 197–232. [Google Scholar]
- Fűrész, A.; Hurta, A.; Kiss, E.; Pápay, G.; Kovács, L.; Péter, N.; Lantos, C.; Penksza, K. Morphotaxonomic and ploidy analysis of dominant Festuca species in sandy grasslands along the Danube. In 62. Georgikon Napok Nemzetközi Tudományos Konferencia: A klímaváltozás kihívásai a következő évtizedekben: Előadások kivonatai; Szent István Egyetem Georgikon Kar: Keszthely, Hungary, 2020; p. 35. [Google Scholar]
- Erdős, L.; Krstonošić, D.; Kiss, P.J.; Bátori, Z.; Tölgyesi, C.; Škvorc, Ž. Plant composition and diversity at edges in a semi-natural forest–grassland mosaic. Plant Ecol. 2019, 220, 279–292. [Google Scholar] [CrossRef] [Green Version]
- Erdős, L.; Tölgyesi, C.; Cseh, V.; Tolnay, D.; Cserhalmi, D.; Körmöczi, L.; Gellény, K.; Bátori, Z. Vegetation history, recent dynamics and future prospects of a Hungarian sandy forest-steppe reserve: Forest-grassland relations, tree species composition and size-class distribution. Community Ecol. 2015, 16, 95–105. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Genetic Horizon | Depth (cm) | SOM (%) | CaCO3 (%) | pH (H2O) | pH (KCl) | Salt (%) |
|---|---|---|---|---|---|---|---|
| F. vaginata | ACk | 0–5 | 0.2 | 11.3 | 8.2 | 8.0 | 0.02 |
| Ck | 5- | 0.1 | 11.4 | 8.6 | 8.2 | 0.02 | |
| F. pseudovaginata | Ak | 0–5 | 1.1 | 6.9 | 8.1 | 7.7 | 0.03 |
| Bwk | 5–18 | 0.6 | 8.6 | 8.3 | 8.1 | 0.03 | |
| 2Ck | 18- | 0.2 | 10.5 | 8.8 | 8.5 | 0.02 | |
| F. wagneri | A | 0–20 | 1.3 | 0 | 6.7 | 6.4 | 0.01 |
| AB | 20–60 | 0.8 | 0 | 6.9 | 6.1 | 0.01 | |
| Bw | 60–95 | 0.5 | 0 | 7.6 | 7.2 | 0.01 | |
| BC | 95–125 | 0.2 | 0 | 7.6 | 7.0 | 0.01 | |
| 2Ckl | 125- | 0.1 | 5.2 | 8.0 | 7.7 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penksza, K.; Saláta, D.; Pápay, G.; Péter, N.; Bajor, Z.; Lisztes-Szabó, Z.; Fűrész, A.; Fuchs, M.; Michéli, E. Do Sandy Grasslands along the Danube in the Carpathian Basin Preserve the Memory of Forest-Steppes? Forests 2021, 12, 114. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020114
Penksza K, Saláta D, Pápay G, Péter N, Bajor Z, Lisztes-Szabó Z, Fűrész A, Fuchs M, Michéli E. Do Sandy Grasslands along the Danube in the Carpathian Basin Preserve the Memory of Forest-Steppes? Forests. 2021; 12(2):114. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020114
Chicago/Turabian StylePenksza, Károly, Dénes Saláta, Gergely Pápay, Norbert Péter, Zoltán Bajor, Zsuzsa Lisztes-Szabó, Attila Fűrész, Márta Fuchs, and Erika Michéli. 2021. "Do Sandy Grasslands along the Danube in the Carpathian Basin Preserve the Memory of Forest-Steppes?" Forests 12, no. 2: 114. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020114