
Habitat Mosaics of Sand Steppes and Forest-Steppes in the Ipoly Valley in Hungary

 ,
,
Abstract
:1. Introduction
- (i)
- Do sand steppes and forest-steppes occur in the Ipoly Valley and if yes, then where?
- (ii)
- What kind of environmental effects influence the species composition on these areas?
- (iii)
- Besides classic habitat mapping, are the satellite data from Sentinel-2A useful for distinction of different areas?
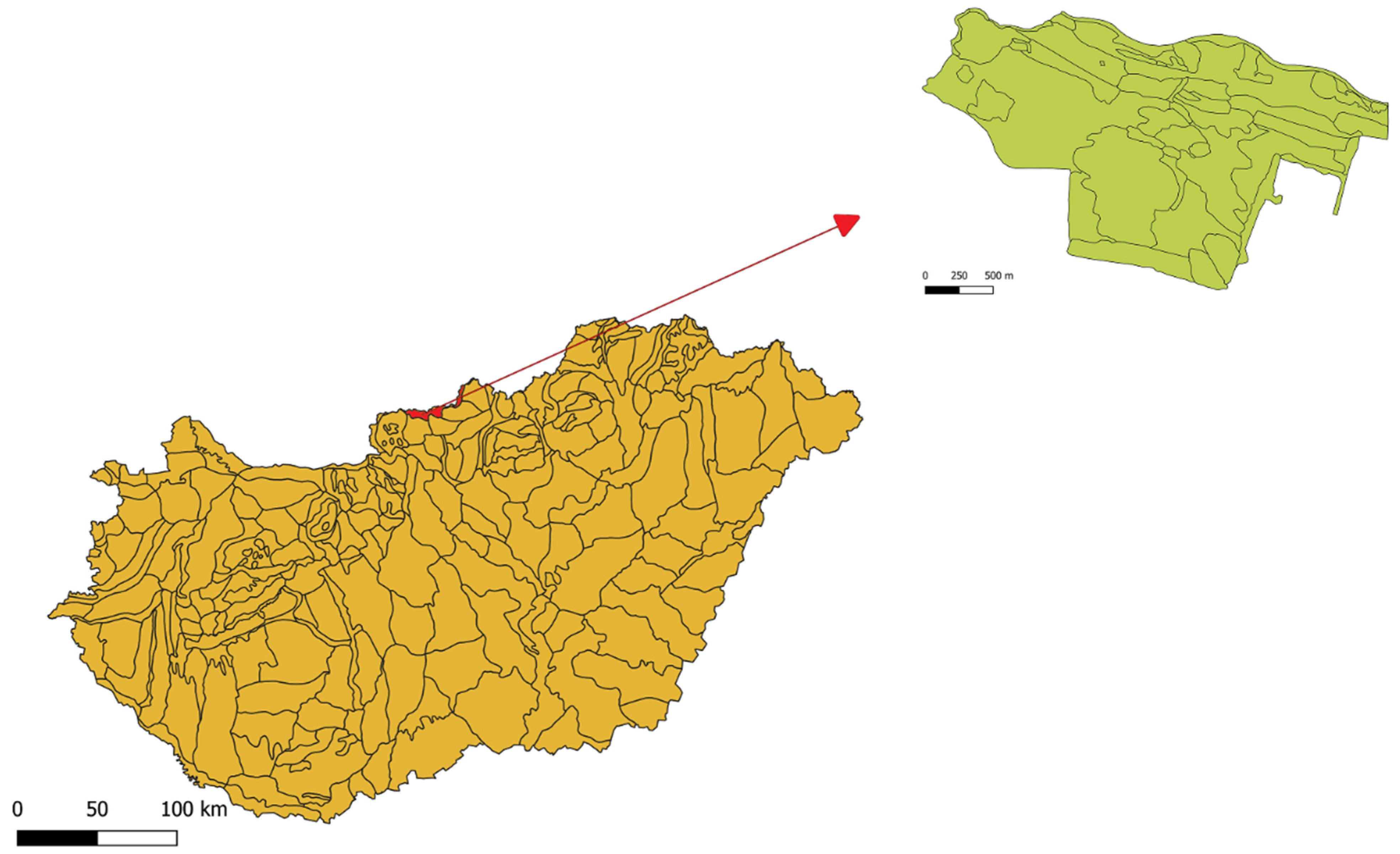
2. Materials and Methods
3. Results
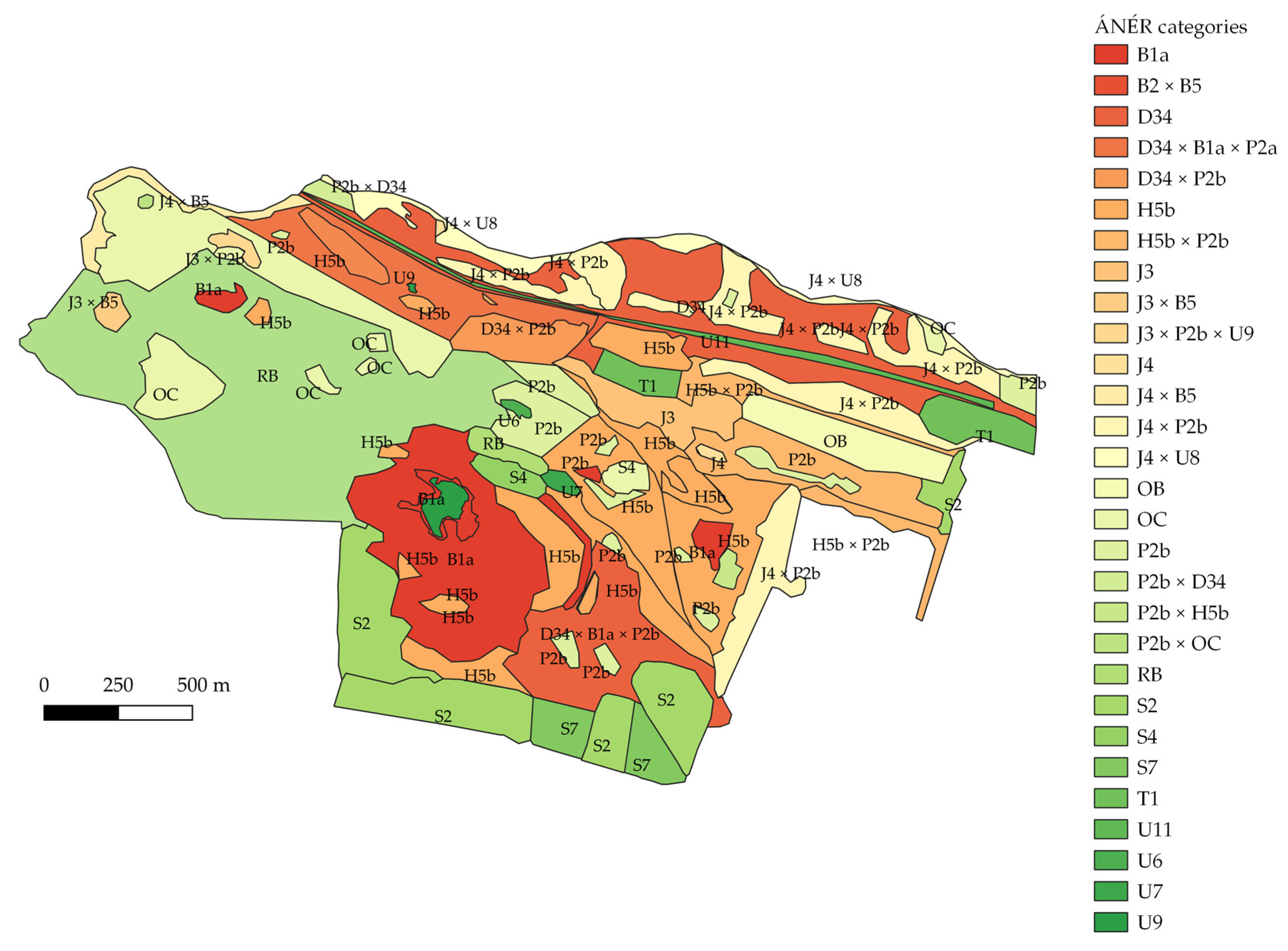
3.1. Habitat Map with ÁNÉR Categories
3.2. Habitat Types of the Study Area
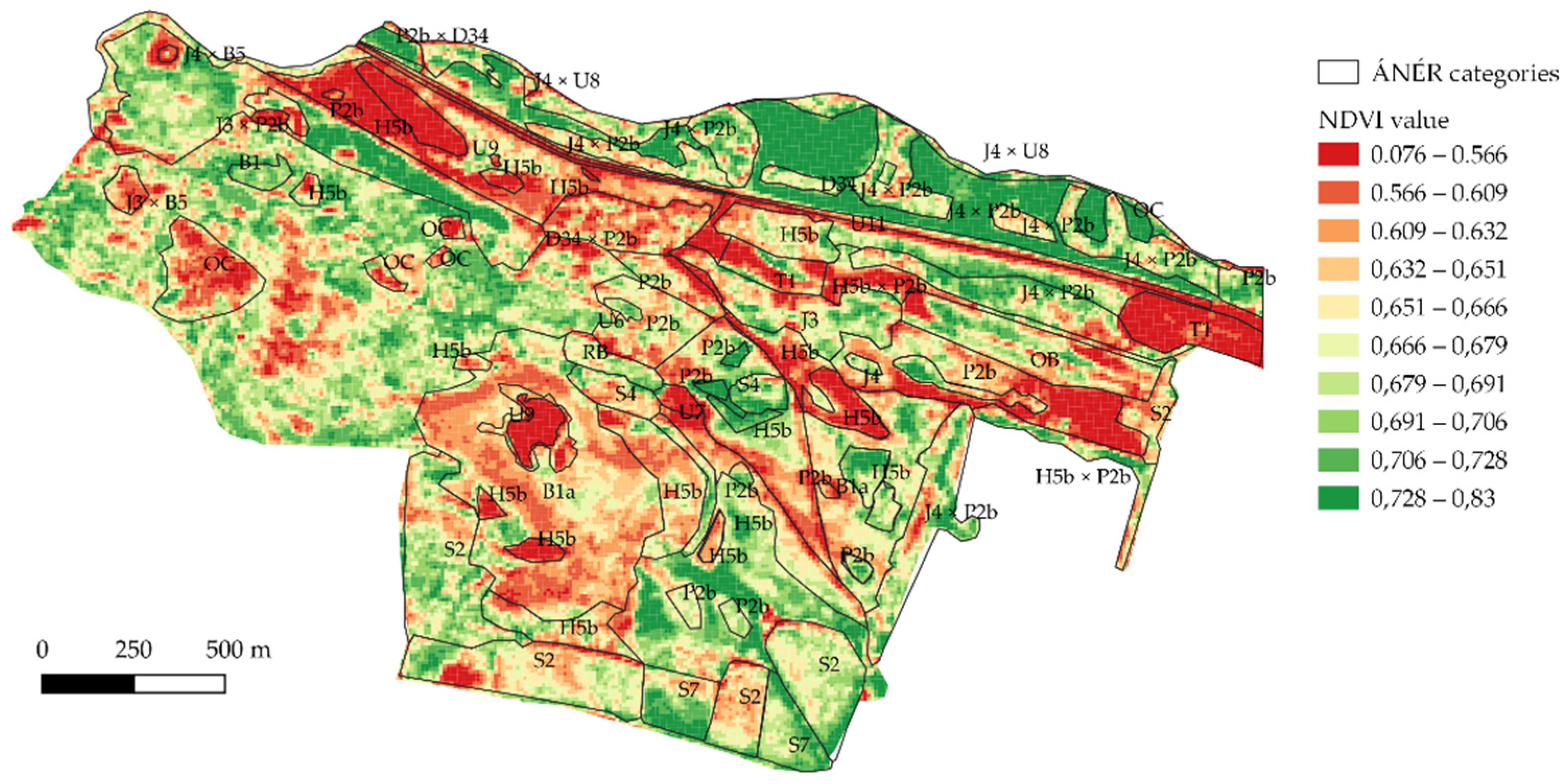
3.3. NDVI versus ÁNÉR Categories
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Török, P.; Penksza, K.; Tóth, E.; Kelemen, A.; Sonkoly, J.; Tóthmérész, B. Vegetation type and grazing intensity jointly shape grasing on grassland biodiversity. Ecol. Evol. 2018, 8, 10326–10335. [Google Scholar] [CrossRef]
- Wesche, K.; Ambarlı, D.; Kamp, J.; Török, P.; Treiber, J.; Dengler, J. The Palaearctic steppe biome: A new synthesis. Biodivers. Conserv. 2016, 25, 2197–2231. [Google Scholar] [CrossRef]
- Zólyomi, B. Budapest és Környékének Természetes Növénytakarója; Pécsi, M., Ed.; Budapest Természeti képe; Akadémiai Kiadó: Budapest, Hungary, 1958. [Google Scholar]
- Szujkóné-Lacza, J.; Kováts, D. The Flora of the Kiskunság National Park; Magyar Természettudományi Múzeum: Budapest, Hungary, 1993; p. 469. [Google Scholar]
- Kárpáti, I.; Kárpáti, V. The aspects of the calciphilous turf (Festucetum vaginale danubiale) in the environs of Vácrátót in 1952. Acta Bot. Hung. 1954, 1, 129–157. [Google Scholar]
- Fekete, G.; Tuba, Z.; Melkó, E. Background processes at the population level during succession in grasslands on sand. Vegetatio 1988, 77, 33–41. [Google Scholar] [CrossRef]
- Varga, Z. Die Waldteppen des pannonischen Raumes aus biogeographischer Sicht. Düsseldorfer Geobotanisches Kolloquium 1989, 6, 35–50. [Google Scholar]
- Molnár, Z.; Biró, M.; Bartha, S.; Fekete, G. Past trends, present state and future prospects of Hungarian forest-steppes. In Ecological Problems and Livelihoods in a Changing World; Werger, M.J.A., van Staalduinen, M.A., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2012; pp. 209–252. [Google Scholar]
- Varga, Z.; Borhidi, A.; Fekete, G.; Debreczy, Z.; Bartha, D.; Bölöni, J.; Molnár, A.; Kun, A.; Molnár, Z.; Lendvai, G.; et al. Alföldi Erdőssztyeppmaradványok Magyarországon/Forest Steppe Remains in the Great Plain of Hungary; WWF-MTA ÖBKI: Budapest-Vácrátót, Hungary, 2000. [Google Scholar]
- Rédei, T.; Csecserits, A.; Barabás, S.; Lhotsky, B.; Botta-Dukát, Z. Homoki erdőssztyeppmozaikok kiterjedésének és változatosságának hatása a fajgazdagságra. The effect of size mand habitat diversity of sand forest steppe mosaics on plant species richness. Természetvédelem és Kutatás a Turjánvidék Északi Részén. Rosalia 2018, 10, 131–144. [Google Scholar]
- Magyari, E.K.; Chapman, J.C.; Passmore, D.G.; Allen, J.R.M.; Huntley, J.P.; Huntley, B. Holocene persistence of wooded steppe in the Great Hungarian Plain. J. Biogeogr. 2010, 37, 915–935. [Google Scholar] [CrossRef]
- Biró, M.; Czúcz, B.; Horváth, F.; Révész, A.; Csatári, B.; Molnár, Z. Drivers of grassland loss in Hungary during the post-socialist transformation (1987–1999). Landsc. Ecol. 2013, 28, 789–803. [Google Scholar] [CrossRef] [Green Version]
- Biró, M.; Horváth, F.; Révész, A.; Molnár, Z.; Vajda, Z. Száraz homoki élőhelyek és átalakulásuk a Duna–Tisza közén a 18. századtól napjainkig. In Természetvédelem és Kutatás a Duna–Tisza közi Homokhátságon. Rosalia 6. Duna–Ipoly Nemzeti Park Igazgatóság; Verő, G., Ed.; Duna Ipoly Nemzeti Park Igazgatóság: Budapest, Hungary, 2011; pp. 383–421. [Google Scholar]
- Tölgyesi, C.; Körmöczi, L. Structural changes of a Pannonian grassland plant community in relation to the decrease of water availability. Acta Bot. Hung. 2012, 54, 413–431. [Google Scholar] [CrossRef]
- Tölgyesi, C.; Bátori, Z.; Erdős, L. Using statistical tests on relative ecological indicator values to compare vegetation units–Different approaches and weighting methods. Ecol. Indic. 2014, 36, 441–446. [Google Scholar] [CrossRef]
- Erdős, L.; Tölgyesi, C.; Horzse, M.; Tolnay, D.; Hurton, Á.; Schulcz, N.; Körmöczi, L.; Lengyel, A.; Bátori, Z. Habitat complexity of the Pannonian forest-steppe zone and its nature conservation implications. Ecol. Complex. 2014, 17, 107–118. [Google Scholar] [CrossRef]
- Erdős, L.; Kröel-Dulay, G.; Bátori, Z.; Kovács, B.; Németh, C.; Kiss, P.J.; Tölgyesi, C. Habitat heterogeneity as a key to high conservation value in forest-grassland mosaics. Biol. Conserv. 2018, 226, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Erdős, L.; Török, P.; Szitár, K.; Bátori, Z.; Tölgyesi, C.; Kiss, P.J.; Bede-Fazekas, Á.; Kröel-Dulay, G. Beyond the forest-grassland dichotomy: The gradient-like organization of habitats in forest-steppes. Front. Plant Sci. 2020, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Tölgyesi, C.; Valkó, O.; Deák, B.; Kelemen, A.; Bragina, T.M.; Gallé, R.; Erdős, L.; Bátori, Z. Tree–herb co-existence and community assembly in natural forest-steppe transitions. Plant Ecol. Divers. 2018, 11, 465–477. [Google Scholar] [CrossRef] [Green Version]
- Fodor, I.; Gálosi-Kovács, B. A Kárpát-medence határokon átnyúló természeti értékei. In A környezet és a Határok Kutatója. Tiszteletkötet Nagy Imre 65. Születésnapja Alkalmából, Regionális Tudományi Társaság; Gál, Z., Ricz, A., Eds.; pp. 39–51. ISBN 978-86-86929-07-5. Available online: http://hdl.handle.net/11155/2088 (accessed on 19 December 2019).
- Brow, A.G.; Lespez, L.; ASear, D.; Macaire, J.; Houben, P.; Klimek, K.; Brazier, R.E.; Van Oost, K.; Pears, B. Natural vs. anthropogenic streams in Europe: History, ecology and implications for restoration, river-rewilding and riverine ecosystem services. Earth-Sci. Rev. 2018, 180, 185–205. [Google Scholar] [CrossRef]
- Penksza, K.; Nagy, A.; Laborczi, A.; Pintér, B.; Házi, J. Wet habitats along River Ipoly (Hungary) in 2000 (extremely dry) and 2010 (extremely wet). J. Maps 2012, 8, 157–164. [Google Scholar] [CrossRef]
- Mjazovszky, Á.; Csontos, P.; Tamás, J. A patakkísérő növényzet vizsgálata négy hazai táj viszonylatában. Botanikai Közlemények 2007, 94, 45–55. [Google Scholar]
- Verrasztó, Z. Környezeti monitoring vizsgálatok az Ipoly vízgyűjtőjén (célkitűzések és általános tájékoztatás). Tájökológiai Lapok 2010, 8, 535–561. [Google Scholar]
- Járdi, I.; Pápay, G.; Fekete, G.; S.-Falusi, E. Marhalegelők vegetációjának vizsgálata az Ipoly-völgy homoki gyepeiben. Gyepgazdálkodási Közlemények 2017, 15, 9–21. [Google Scholar]
- Sentinel-2A Website. Available online: https://scihub.copernicus.eu/ (accessed on 23 June 2015).
- Burai, P.; Lenart, C.; Valkó, O.; Bekő, L.; Szabó, Z.; Deák, B. Fátlan vegetációtípusok azonosítása légi hiperspektrális távérzékelési módszerrel. Tájökológiai Lapok 2016, 14, 1–12. [Google Scholar]
- Marosi, S.; Somogyi, S. Magyarország Kistájainak Katasztere I-II; MTA Földrajztudományi Kutató Intézet: Budapest, Hungary, 1990; 1023p. [Google Scholar]
- Fekete, G.; Molnár, Z.; Horváth, F. (Eds.) A Magyarországi Élőhelyek Leírása, Határozója és a Nemzeti Élőhely-osztályozási Rendszer. (Description of and Guide to Hungarian Habitats); Magyar Természettudományi Múzeum: Budapest, Hungary, 1997. [Google Scholar]
- Bölöni, J.; Molnár, Z.; Horváth, F.; Illyés, E. Naturalness-based habitat quality of the hungarian (semi-) natural habitats. Acta Bot. Hung. 2008, 50, 149–159. [Google Scholar] [CrossRef]
- Bölöni, J.; Molnár, Z.; Kun, A. (Eds.) Magyarország Élőhelyei. A Hazai Vegetációtípusok Leírása és Határozója; ÁNÉR 2011; MTA ÖBKI: Vácrátót, Hungary, 2011; p. 441. [Google Scholar]
- Borhidi, A.; Kevey, B.; Lendvai, G.; Seregélyes, T. Plant Communities of Hungary; Akadémiai Kiadó: Budapest, Hungary, 2012. [Google Scholar]
- Király, G. (Ed.) Új Magyar Füvészkönyv. Magyarország Hajtásos Növényei. Határozókulcsok [New Hungarian Herbal. The Vascular Plants of Hungary. Identification Key]; ANP Igazgatóság: Jósvafő, Hungary, 2009. [Google Scholar]
- Didan, D. MOD13Q1 MODIS/Terra Vegetation Indices 16-Day L3 Global 250m SIN Grid V006; NASA EOSDIS Land Processes DAAC; NASA: College Park, MD, USA, 2015.
- Hammer, Ø.; Harper, D.A.T.; Paul, D.R. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electr. 2001, 4, 9. [Google Scholar]
- Borhidi, A. Magyarország Növénytárulásai; Akadémiai Kiadó: Budapest, Hungary, 2003; p. 610. [Google Scholar]
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian flora. Acta Bot. Acad. Sci. Hung. 1995, 39, 97–181. [Google Scholar]
- Simon, T. A Magyarországi Edényes Flóra Határozója; Tankönyvkiadó: Budapest, Hungary, 2000. [Google Scholar]
- Soó, R. A Magyar Flóra és Vegetáció Rendszertani-Növényföldrajzi Kézikönyve I. (Synopsis Systematico-Geobotanica Florae Vegetationisque Hungariae I); Akadémia Kiadó: Budapest, Hungary, 1964. [Google Scholar]
- Pongrácz, R.; Bartholy, J.; Szabó, P.; Gelybó, G. A comparison of observed trends and simulated changes in extreme climate indices in the Carpathian Basin by the end of this century. Int. J. Glob. Warm. 2009, 1, 336–355. [Google Scholar] [CrossRef]
- Malatinszky, Á.; Ádám, S.; Falusi, E.; Saláta, D.; Penksza, K. Climate change related land user problems in protected wetlands: A study in a seriously affected Hungarian area. Clim. Chang. 2013, 118, 671–683. [Google Scholar] [CrossRef]
- Uj, B.; Nagy, A.; Saláta, D.; Laborczi, A.; Malatinszky, A.; Bakó, G.; Danyik, T.; Tóth, A.; Falusi, E.S.; Gyuricza, C.; et al. Habitat map of Kis-Sárrét (Körös-Maros National Park, Hungary) with special regard to the changes of wetlands. J. Maps 2015, 10, 211–221. [Google Scholar] [CrossRef]
- Körner, C. A reassessment of high-elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar]
- Courtwright, J.; Findlay, S.E.G. Effects of microtopography on hydrology, physicochemistry, and vegetation in a tidal swamp of the Hudson River. Wetlands 2011, 31, 239–249. [Google Scholar] [CrossRef]
- Bátori, Z.; Farkas, T.; Erdős, L.; Tölgyesi, C.; Körmöczi, L.; Vojtkó, A. A comparison of the vegetation of forested and non-forested solution dolines in Hungary: A preliminary study. Biologia 2014, 69, 1339–1348. [Google Scholar] [CrossRef]
- Bekkema, M.; Eleveld, M. Mapping Grassland Management Intensity Using Sentinel-2 Satellite Data. GI_Forum 2018, 1, 194–213. [Google Scholar] [CrossRef]
- Kaplan, G. Mapping and Monitoring Wetlands Using Sentinel 2 Satellite Imagery. Available online: https://pdfs.semanticscholar.org/a101/515a9d639c896364cec0b589172af3649717.pdf (accessed on 15 October 2017).
- Majasalmi, T.; Rautiainen, M. The potential of Sentinel-2 data for estimating biophysical variables in a boreal forest: A simulation study. Remote Sens. Lett. 2016, 7, 427–436. [Google Scholar] [CrossRef]
- Veloso, A.; Stéphane, M.; Bouvet, A.; Le Toan, T.; Planells, M.; Dejoux, J.; Ceschia, E. Understanding the temporal behavior of crops using Sentinel-1 and Sentinel-2-like data for agricultural applications. Remote Sens. Environ. 2017, 199, 415–426. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Jönsson, P.; Høgda, K.-A.; Karlsen, S.R.; Eklundh, L.; Skidmore, A.K. A ground-validated NDVI dataset for monitoring vegetation dynamics and mapping phenology in Fennoscandia and the Kola peninsula. Int. J. Remote Sens. 2007, 28, 4311–4330. [Google Scholar] [CrossRef]
- Bekkema, M.E. The Potential of Sentinel-2 Data for Detecting Grassland Management Intensity to Support Monitoring of Meadow Bird Populations. Master’s Thesis, UNIGIS, Amsterdam, The Netherlands, 2017. [Google Scholar]
- Maglocký, Š.; Feráková, V. Red list of ferns and flowering plants (Pteridophyta and Spermathophyta) of the flora of Slovakia. Biologia 1993, 48, 361–385. [Google Scholar]
- Zsilinszki, A.; Dezső, Z.; Bartholy, J.; Pongrácz, R. Synoptic-climatological analysis of high level air flow over the Carpathian Basin. Időjárás/Q. J. Hung. Meteorol. Serv. 2019, 123, 19–38. [Google Scholar] [CrossRef] [Green Version]
- Bartholy, J.; Pongrácz, R. Tendencies of extreme climate indicates based on daily precipitation in the Carpathian Basin for the 20th century. Időjárás 2005, 109, 1–20. [Google Scholar]
- Bartholy, J.; Pongrácz, R. Comparing tendencies of some temperature and precipitation on global and regional scales. Időjárás 2006, 110, 35–48. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ÁNÉR Categories | Description |
|---|---|
| B1a | Eu- and mesotrophic reed and Typha beds |
| B5 | Non-tussock tall-sedge beds |
| D34 | Mesotrophic wet meadows |
| H5b | Closed sand steppes |
| P2b | Dry and semi-dry pioneer scrub |
| P2a | Wet and mesic pioneer scrub |
| J3 | Riverine willow scrub |
| J4 | Riverine willow-poplar woodlands |
| OB | Uncharacteristic mesic grasslands |
| OC | Uncharacteristic dry and semi-dry grasslands |
| P2b | Dry and semi-dry pioneer scrub |
| RB | Uncharacteristic or pioneer softwood forests |
| S2 | Populus × euramericana plantations |
| S4 | Scots and black pine plantations |
| T1 | Annual intensive arable fields |
| U8 | Water streams |
| U7 | Sand, gravel, clay and peat mines, loess walls |
| U9 | Standing waters |
| U11 | Roads and railroads |
| Type of Habitats | Number of Habitat Patches (Pieces) | Area (Hectares) |
|---|---|---|
| B1a | 4 | 46.0 |
| B2 × B5 | 1 | 17.5 |
| D34 | 1 | 34.1 |
| D34 × B1a × P2a | 1 | 16.5 |
| H5b | 16 | 38.0 |
| H5b × P2b | 2 | 28.1 |
| J3 | 1 | 6.8 |
| J3 × B5 | 1 | 1.0 |
| J3 × P2b × U9 | 1 | 0.9 |
| J4 | 2 | 0.5 |
| J4 × B5 | 1 | 3.3 |
| J4 × P2b | 7 | 25.7 |
| J4 × P2b × U9 | 1 | 2.7 |
| J4 × U8 | 2 | 11.3 |
| OB | 1 | 8.1 |
| OC | 8 | 30.6 |
| P2b | 15 | 18.0 |
| P2b × D34 | 1 | 1.1 |
| P2b × H5b | 1 | 13.1 |
| P2b × OC | 1 | 0.2 |
| RB | 2 | 84.9 |
| S2 | 5 | 30.4 |
| S4 | 2 | 3.5 |
| S7 | 2 | 6.2 |
| T1 | 2 | 7.1 |
| U11 | 3 | 6.8 |
| U6 | 1 | 0.4 |
| U7 | 1 | 0.7 |
| U9 | 2 | 1.7 |
| Total | 88 | 445.1 |
| B2 × B5 | D34 | H5b | H5b × P2b | D34 × B1a × P2a | J3 × B5 | J4 × B5 | J4 × P2b | OB | OC | P2b | RB | S2 | S4 | T1 | U7 | U9 | U11 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B1a | ns | ns | * | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | * | ** | *** | *** | *** |
| B2 × B5 | ns | *** | ns | ns | ns | ns | ns | *** | *** | ns | ** | *** | ns | *** | *** | *** | *** | |
| D34 | *** | ns | ns | ns | ns | ns | * | *** | ns | * | *** | ns | *** | *** | *** | *** | ||
| H5b | * | *** | *** | *** | *** | ns | ns | ** | ns | ns | *** | ns | ns | ns | ns | |||
| H5b × P2b | ns | ns | ns | ns | ns | ns | ns | ns | ns | ** | ** | *** | *** | *** | ||||
| D34 × B1a × P2a | ns | ns | ns | ns | *** | ns | ns | * | ns | *** | *** | *** | *** | |||||
| J3 × B5 | ns | ns | ns | ** | ns | ns | ns | ns | *** | *** | *** | *** | ||||||
| J4 × B5 | ns | *** | *** | ns | ** | *** | ns | *** | *** | *** | *** | |||||||
| J4 × P2b | ns | * | ns | ns | ns | ns | *** | *** | *** | *** | ||||||||
| OB | ns | ns | ns | ns | *** | ns | * | ** | * | |||||||||
| OC | ns | ns | ns | *** | ns | ns | ns | ns | ||||||||||
| P2b | ns | ns | ns | *** | *** | *** | *** | |||||||||||
| RB | ns | *** | ns | ** | ** | ** | ||||||||||||
| S2 | *** | ns | ns | ns | ns | |||||||||||||
| S4 | *** | *** | *** | *** | ||||||||||||||
| T1 | ns | ns | ns | |||||||||||||||
| U7 | ns | ns | ||||||||||||||||
| U9 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Járdi, I.; Saláta, D.; S.-Falusi, E.; Stilling, F.; Pápay, G.; Zachar, Z.; Falvai, D.; Csontos, P.; Péter, N.; Penksza, K. Habitat Mosaics of Sand Steppes and Forest-Steppes in the Ipoly Valley in Hungary. Forests 2021, 12, 135. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020135
Járdi I, Saláta D, S.-Falusi E, Stilling F, Pápay G, Zachar Z, Falvai D, Csontos P, Péter N, Penksza K. Habitat Mosaics of Sand Steppes and Forest-Steppes in the Ipoly Valley in Hungary. Forests. 2021; 12(2):135. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020135
Chicago/Turabian StyleJárdi, Ildikó, Dénes Saláta, Eszter S.-Falusi, Ferenc Stilling, Gergely Pápay, Zalán Zachar, Dominika Falvai, Péter Csontos, Norbert Péter, and Károly Penksza. 2021. "Habitat Mosaics of Sand Steppes and Forest-Steppes in the Ipoly Valley in Hungary" Forests 12, no. 2: 135. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020135