
Distribution of Gonipterus Species and Their Egg Parasitoids in Australia: Implications for Biological Control

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect and Plant Collections
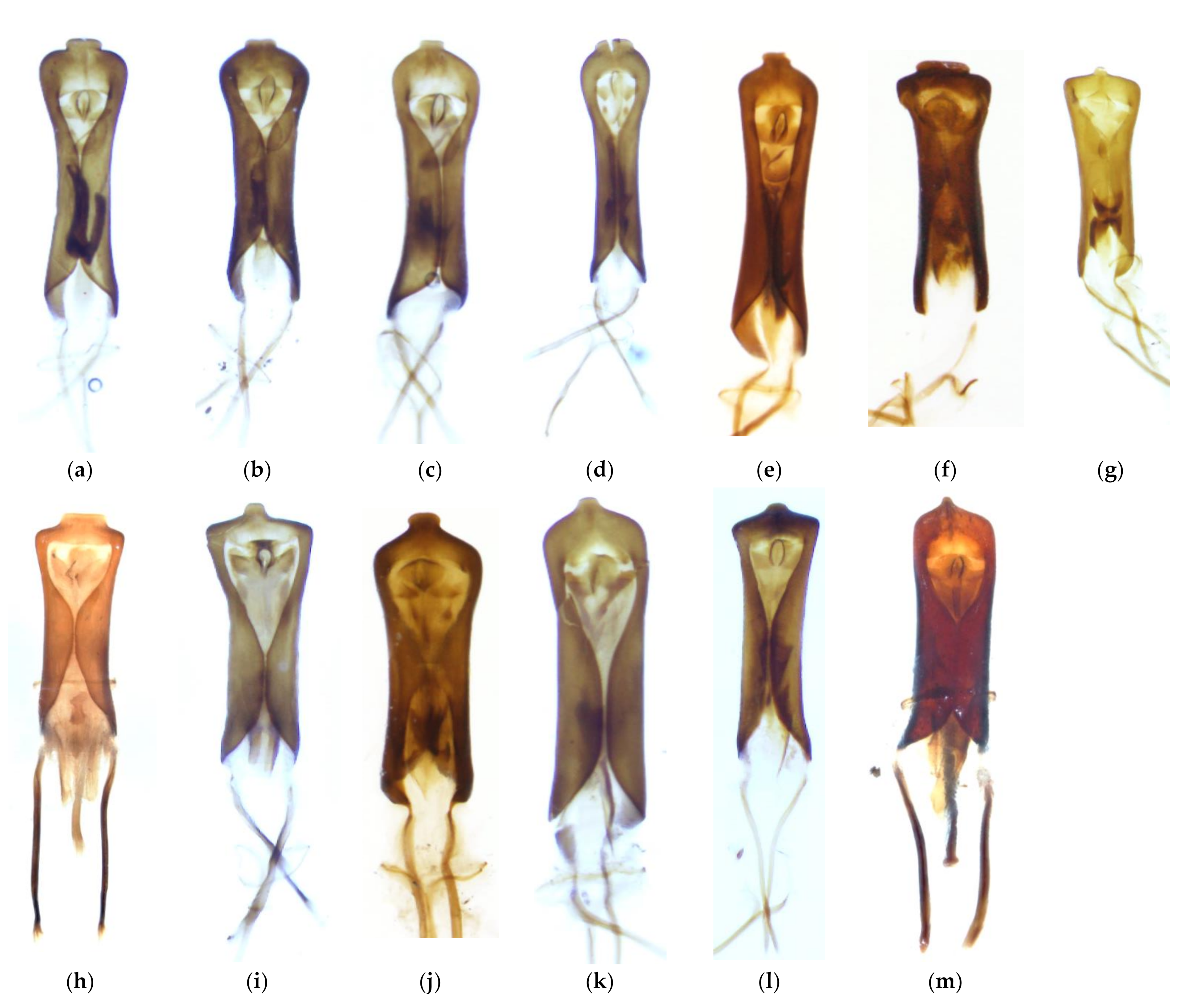
2.2. Species Identification
2.3. DNA Barcoding
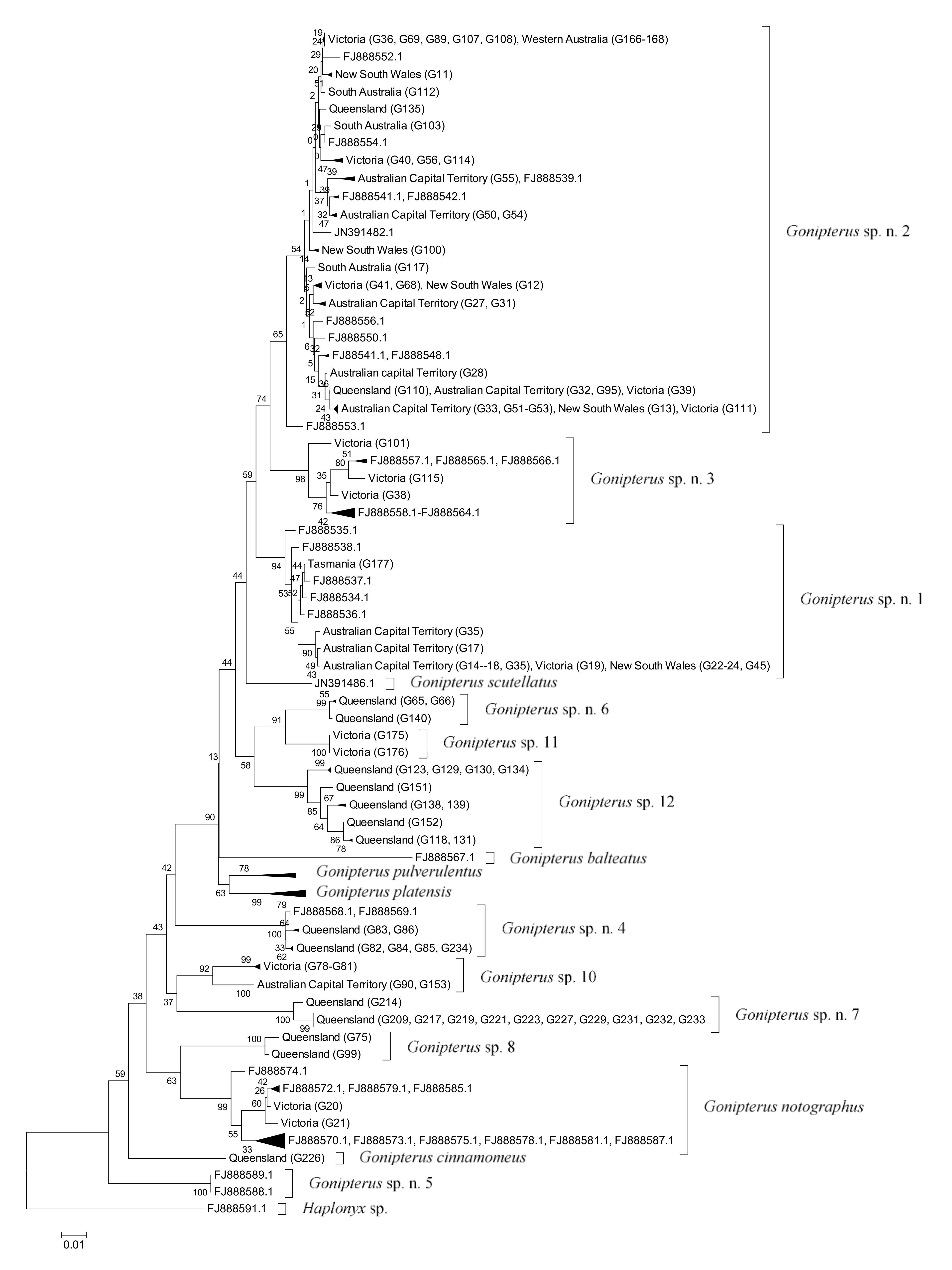
2.4. Data Analysis
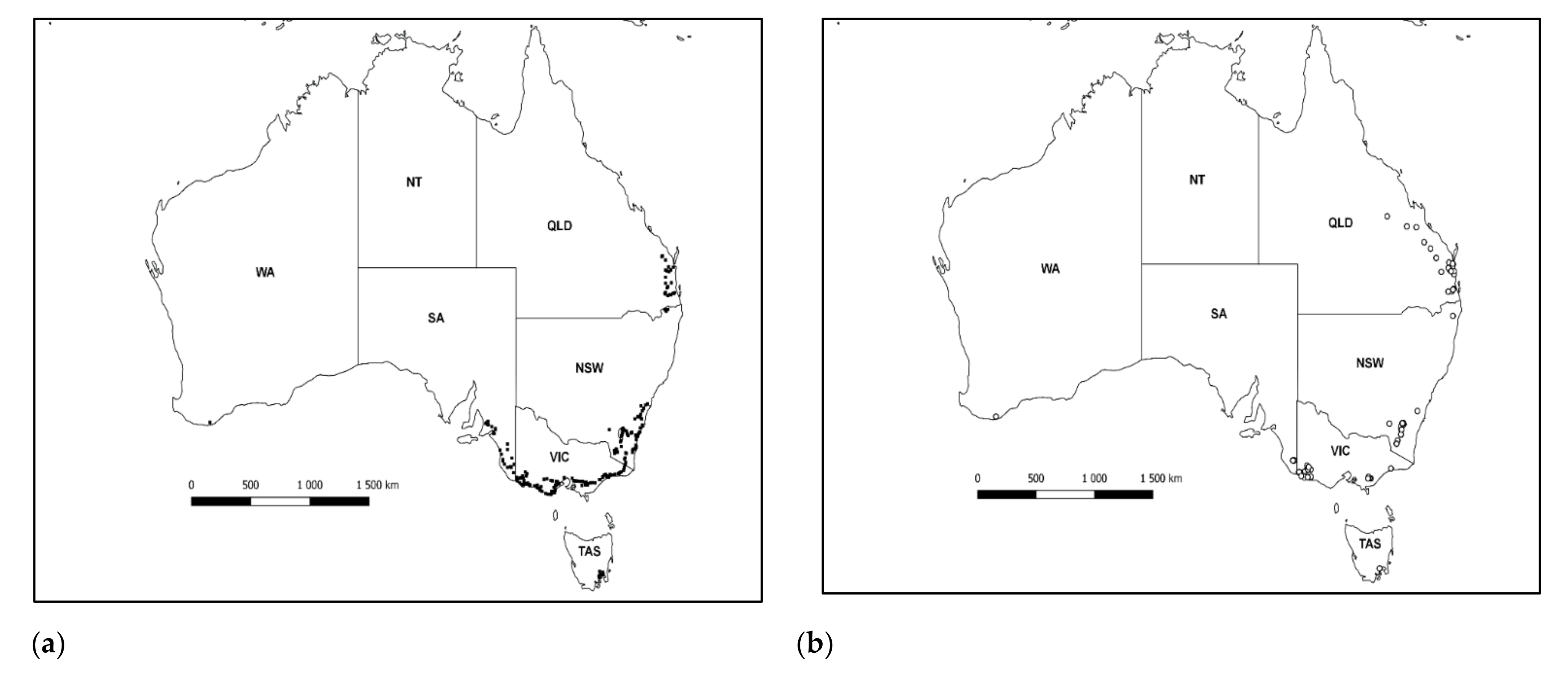
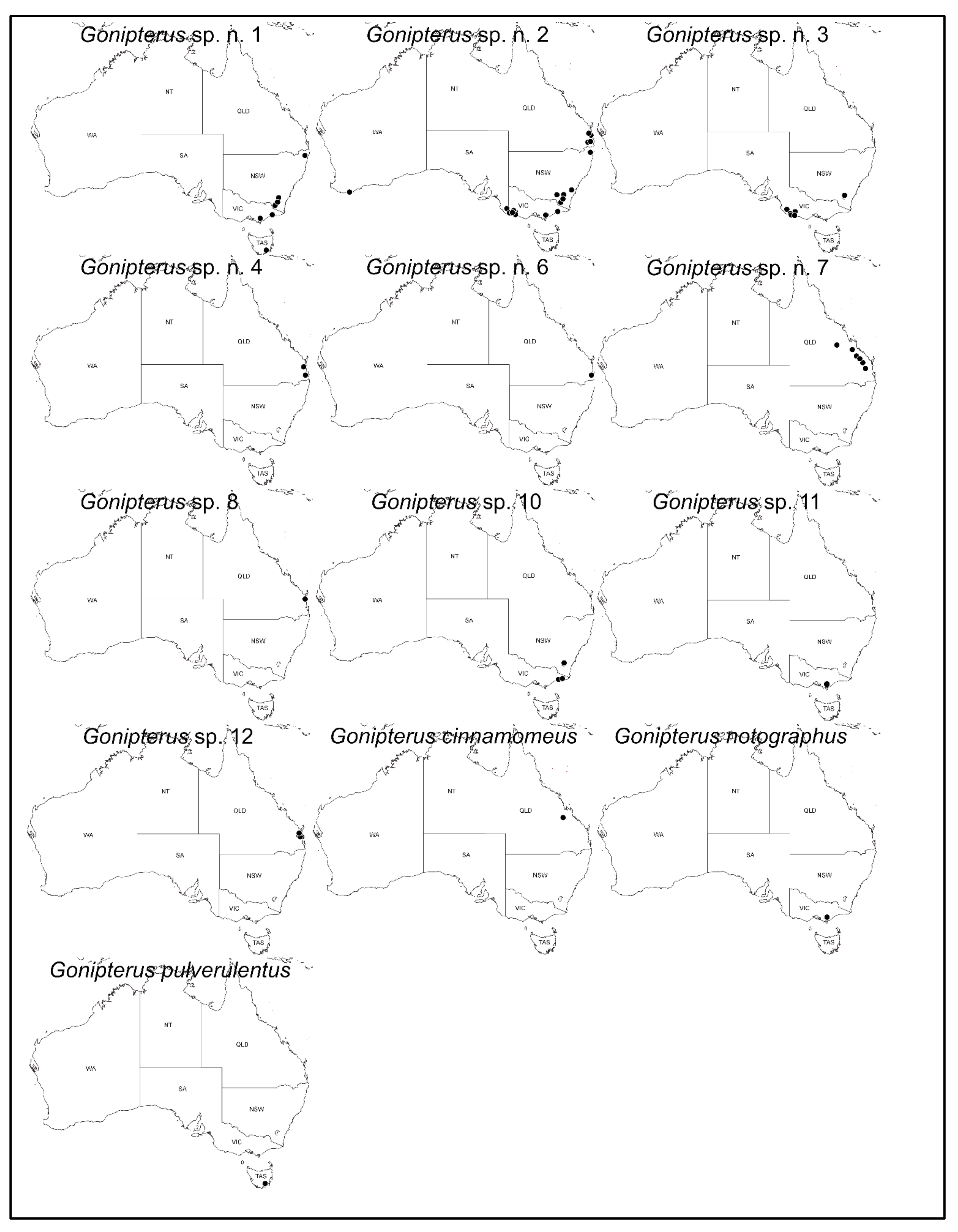
3. Results
3.1. Identification and Distribution of Gonipterus Species
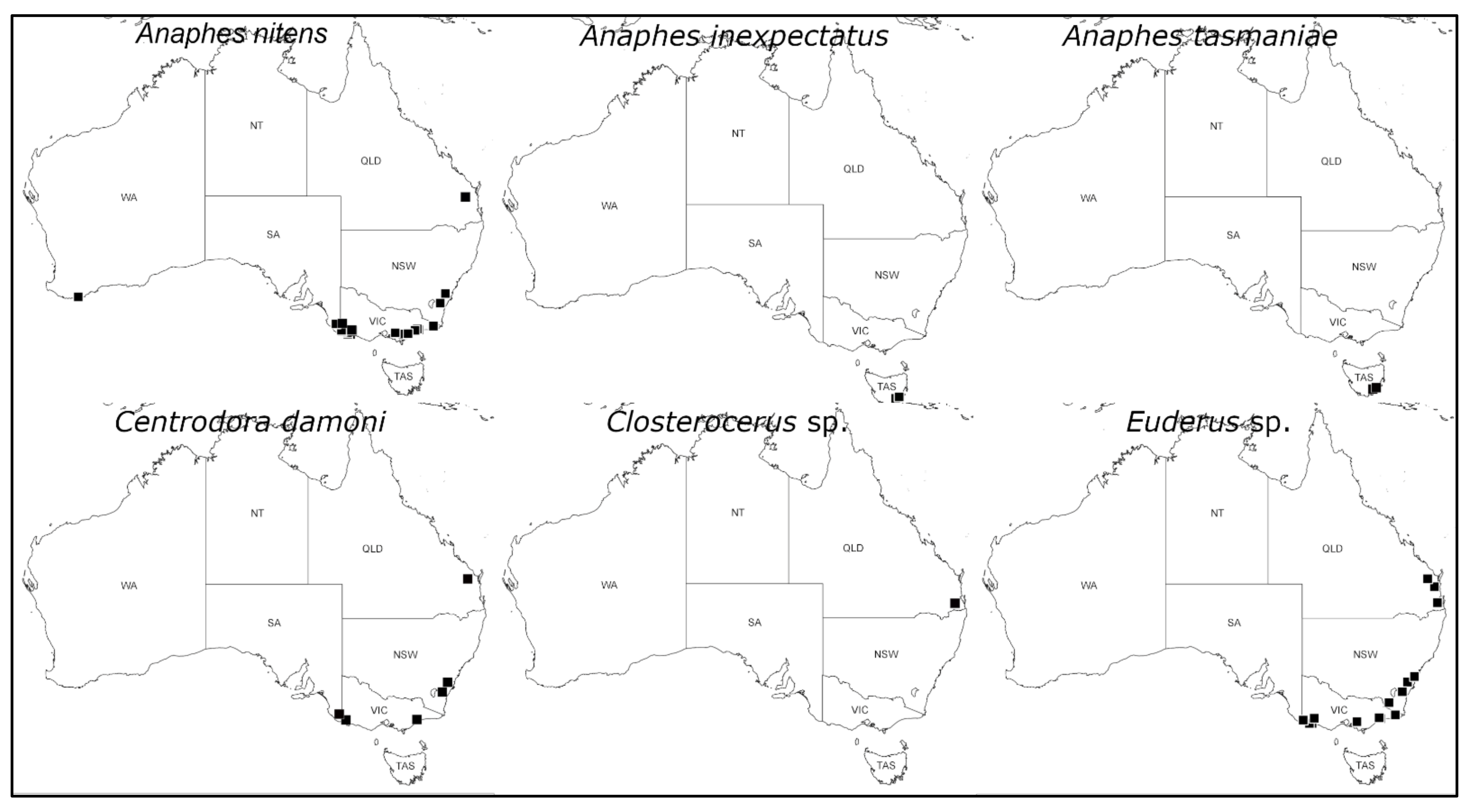
3.2. Identification and Distribution of Gonipterus Egg Parasitoids
3.3. Gonipterus Host Plant Association
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Klinken, R.D.; Raghu, S. A scientific approach to agent selection. Aust. J. Entomol. 2006, 45, 253–258. [Google Scholar] [CrossRef]
- Dhileepan, K.; Wilmot Senaratne, K.A.D.; Raghu, S. A systematic approach to biological control agent exploration and prioritisation for prickly acacia (Acacia nilotica ssp. indica). Aust. J. Entomol. 2006, 45, 303–307. [Google Scholar] [CrossRef]
- Hoelmer, K.A.; Kirk, A.A. Selecting arthropod biological control agents against arthropod pests: Can the science be improved to decrease the risk of releasing ineffective agents? Biol. Control 2005, 34, 255–264. [Google Scholar] [CrossRef]
- Goolsby, J.A.; De Barro, P.J.; Makinson, J.R.; Pemberton, R.W.; Hartley, D.M.; Frohlich, D.R. Matching the origin of an invasive weed for selection of a herbivore haplotype for a biological control programme. Mol. Ecol. 2006, 15, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Goolsby, J.A.; Guerrero, F.D.; Gaskin, J.; Bendele, K.G.; Azhahianambi, P.; Amalin, D.; Flores-Cruz, M.; Kashefi, J.; Smith, L.; Racelis, A.; et al. Molecular comparison of cattle fever ticks from native and introduced ranges, with insights into optimal search areas for classical biological control agents. Southwest. Entomol. 2016, 41, 595–604. [Google Scholar] [CrossRef]
- DeBach, P. Biological Control by Natural Enemies; Cambridge University Press: Cambridge, UK, 1974; pp. 1–323. [Google Scholar]
- Rosen, D. The role of taxonomy in effective biological control programs. Agric. Ecosyst. Environ. 1986, 15, 121–129. [Google Scholar] [CrossRef]
- de Moraes, G.J. Importance of taxonomy in biological control. Insect. Sci. Appl. 1987, 8, 841–844. [Google Scholar] [CrossRef]
- Stiling, P. Why do natural enemies fail in classical biological control programs? Am. Entomol. 1993, 39, 31–37. [Google Scholar] [CrossRef]
- Fiaboe, K.K.M.; Fonseca, R.L.; De Moraes, G.J.; Ogol, C.K.P.O.; Knapp, M. Identification of priority areas in South America for exploration of natural enemies for classical biological control of Tetranychus evansi (Acari: Tetranychidae) in Africa. Biol. Control 2006, 38, 373–379. [Google Scholar] [CrossRef]
- Buffington, M.L.; Talamas, E.J.; Hoelmer, K.A. Team Trissolcus: Integrating taxonomy and biological control to combat the brown marmorated stink bug. Am. Entomol. 2018, 64, 224–232. [Google Scholar] [CrossRef]
- Debach, P. The importance of taxonomy to biological control as illustrated by the cryptic history of Aphytis holoxanthus n. sp. (Hymenoptera: Aphelinidae), a parasite of Chrysomphalus aonidum, and Aphytis coheni n. sp., a parasite of Aonidiella aurantii. Ann. Entomol. Soc. Ann. 1960, 53, 701–705. [Google Scholar] [CrossRef]
- Toševski, I.; Caldara, R.; Jovic, J.; Hernández-Vera, G.; Baviera, C.; Gassmann, A.; Emerson, B.C. Morphological, molecular and biological evidence reveal two cryptic species in Mecinus janthinus Germar (Coleoptera, Curculionidae), a successful biological control agent of Dalmatian toadflax, Linaria dalmatica (Lamiales, Plantaginaceae). Syst. Entomol. 2011, 36, 741–753. [Google Scholar] [CrossRef]
- Mapondera, T.S.; Burgess, T.; Matsuki, M.; Oberprieler, R.G. Identification and molecular phylogenetics of the cryptic species of the Gonipterus scutellatus complex (Coleoptera: Curculionidae: Gonipterini). Aust. J. Entomol. 2012, 51, 175–188. [Google Scholar] [CrossRef]
- Barratt, B.I.P.; Cock, M.J.W.; Oberprieler, R.G. Weevils as targets for biological control, and the importance of taxonomy and phylogeny for efficacy and biosafety. Diversity 2018, 10, 73. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.; Cristofaro, M.; Bon, M.-C.; De Biase, A.; Petanović, R.; Vidović, B. The importance of cryptic species and subspecific populations in classic biological control of weeds: A North American perspective. BioControl 2018, 63, 417–425. [Google Scholar] [CrossRef]
- Beard, J.J. Taxonomy and biological control: Neoseiulus cucumeris (Acari: Phytoseiidae), a case study. Aust. J. Entomol. 1999, 38, 51–59. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rissler, L.J.; Apodaca, J.J. Adding more ecology into species delimitation: Ecological niche models and phylogeography help define cryptic species in the Black Salamander (Aneides flavipunctatus). Syst. Biol. 2007, 56, 924–942. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.M.; Janzen, D.H.; Hajibabaei, M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes and cryptic species of skipper butterflies in the genus Perichares in Area de Conservación Guanacaste, Costa Rica. Proc. Natl. Acad. Sci. USA 2008, 105, 6350–6355. [Google Scholar] [CrossRef] [Green Version]
- Heethoff, M.; Laumann, M.; Weigmann, G.; Raspotnig, G. Integrative taxonomy: Combining morphological, molecular and chemical data for species delineation in the parthenogenetic Trhypochthonius tectorum complex (Acari, Oribatida, Trhypochthoniidae). Front. Zool. 2011, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peixoto, L.; Allen, G.R.; Ridenbaugh, R.D.; Quarrell, S.R.; Withers, T.M.; Sharanowski, B.J. When taxonomy and biological control researchers unite: Species delimitation of Eadya parasitoids (Braconidae) and consequences for classical biological control of invasive paropsine pests of Eucalyptus. PLoS ONE 2018, 13, e0201276. [Google Scholar] [CrossRef] [Green Version]
- Seehausen, M.L.; Ris, N.; Driss, L.; Racca, A.; Girod, P.; Warot, S.; Borowiec, N.; Toševski, I.; Kenis, M. Evidence for a cryptic parasitoid species reveals its suitability as a biological control agent. Sci. Rep. 2020, 10, 19096. [Google Scholar] [CrossRef] [PubMed]
- Tooke, F.G.C. The Eucalyptus Snout Beetle Gonipterus scutellatus Gyll: A Study of Its Ecology and Control by Biological Means; Department of Agriculture, Devision of Entomology: Pretoria, South Africa, 1955; pp. 1–282. [Google Scholar]
- Reis, A.R.; Ferreira, L.; Tomé, M.; Araujo, C.; Branco, M. Efficiency of biological control of Gonipterus platensis (Coleoptera: Curculionidae) by Anaphes nitens (Hymenoptera: Mymaridae) in cold areas of the Iberian Peninsula: Implications for defoliation and wood production in Eucalyptus globulus. For. Ecol. Manag. 2012, 270, 216–222. [Google Scholar] [CrossRef]
- Garcia, A.; Allen, G.R.; Oberprieler, R.G.; Ramos, A.P.; Valente, C.; Reis, A.; Franco, J.C.; Branco, M. Biological control of Gonipterus: Uncovering the associations between eucalypts, weevils and parasitoids in their native range. For. Ecol. Manag. 2019, 443, 106–116. [Google Scholar] [CrossRef]
- Hanks, L.; Millar, J.; Paine, T.D.; Campbell, C. Classical biological control of the Australian weevil Gonipterus scutellatus (Coleoptera: Curculionidae) in California. Environ. Entomol. 2000, 29, 369–375. [Google Scholar] [CrossRef]
- Schröder, M.L.; Slippers, B.; Wingfield, M.J.; Hurley, B.P. Invasion history and management of Eucalyptus snout beetles in the Gonipterus scutellatus species complex. J. Pest Sci. 2020, 93, 11–25. [Google Scholar] [CrossRef]
- Clark, A.F. The parasite control of Gonipterus scutellatus Gyll. N. Z. J. Sci. Tech. 1931, 13, 22–28. [Google Scholar]
- Williams, J.R.; Moutia, L.A.; Hermelin, P.R. The biological control of Gonipterus scutellatus Gyll. (Col. Curculionidae) in Mauritius. Bull. Entomol. Res. 1951, 42, 23–28. [Google Scholar] [CrossRef]
- Cadahia, D. Proximidad de dos nuevos enemigos de los Eucalyptus en España. Bol. Serv. Plagas 1980, 6, 165–192. [Google Scholar]
- Pinet, C. Patasson nitens, parasite spécifique de Gonipterus scutellatus en France. EPPO 1986, 16, 285–287. [Google Scholar] [CrossRef]
- Lanfranco, D.; Dungey, H.S. Insect damage in Eucalyptus: A review of plantations in Chile. Austral Ecol. 2001, 26, 477–481. [Google Scholar] [CrossRef]
- Cordero Rivera, A.; Santolamazza Carbone, S.; Andrés, J.A. Life cycle and biological control of the Eucalyptus snout beetle (Coleoptera, Curculionidae) by Anaphes nitens (Hymenoptera, Mymaridae) in north-west Spain. Agric. For. Entomol. 1999, 1, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Souza, N.M.; Junqueira, L.R.; Wilcken, C.F.; Soliman, E.P.; Camargo, M.B.; Nickele, M.A.; Barbosa, L.R. Ressurgência de uma antiga ameaça: Gorgulho-do-eucalipto Gonipterus platensis (Coleoptera: Curculionidae). In Instituto de Pesquisas e Estudos Florestais; Instituto de Pesquisas e Estudos Florestais: Piracicaba, Brazil, 2016; p. 20. [Google Scholar]
- Valente, C.; Gonçalves, C.I.; Monteiro, F.; Gaspar, J.; Silva, M.; Sottomayor, M.; Paiva, M.R.; Branco, M. Economic outcome of classical biological control: A case study on the Eucalyptus Snout Beetle, Gonipterus platensis, and the parasitoid Anaphes nitens. Ecol. Econ. 2018, 149, 40–47. [Google Scholar] [CrossRef]
- Valente, C.; Vaz, A.; Pina, J.; Manta, A.; Sequeira, A. Control strategy against the Eucalyptus Snout Beetle, Gonipterus scutellatus Gyllenhal (Coleoptera, Curculionidae), by the portuguese cellulose industry. In Eucalyptus in a Changing World, International IUFRO Conference of the WP2.08.03 on Silviculture and Improvement of Eucalypts, Aveiro, Portugal, 11–15 October 2004; Borralho, N.M.G., Ed.; RAIZ—Instituto de Investigacao da Floresta e Papel: Aveiro, Portugal, 2004. [Google Scholar]
- Valente, C.; Gonçalves, C.I.; Reis, A.; Branco, M. Pre-selection and biological potential of the egg parasitoid Anaphes inexpectatus for the control of the Eucalyptus snout beetle, Gonipterus platensis. J. Pest Sci. 2017, 90, 911–923. [Google Scholar] [CrossRef]
- Huber, J.T.; Prinsloo, G.L. Redescription of Anaphes nitens (Girault) and description of two new species of Anaphes (Haliday) (Hymenoptera: Mymaridae), parasites of Gonipterus scutellatus Gyllenhal (Coleoptera: Curculionidae) in Tasmania. J. Aust. Ent. Soc. 1990, 29, 333–341. [Google Scholar] [CrossRef]
- Howland, D.E.; Hewitt, G.M. Phylogeny of the Coleoptera based on mitochondrial cytochrome oxidase I sequence data. Insect Mol. Biol. 1995, 4, 203–215. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. Ali View: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Brooker, M.I.H. A new classification of the genus Eucalyptus L’Her (Myrtaceae). Aust. Syst. Bot. 2000, 13, 79–148. [Google Scholar] [CrossRef]
- Noyes, J.S. Universal Chalcidoidea Database. World Wide Web Electronic Publication. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 15 November 2019).
- Edwards, C.M.; La Salle, J. A new species of Closterocerus Westwood (Hymenoptera: Eulophidae), a parasitoid of serpentine leafminers (Diptera: Agromyzidae) from Australia. Aust. J. Entomol. 2004, 43, 129–132. [Google Scholar] [CrossRef]
- Protasov, A.; Blumberg, D.; Brand, D.; La Salle, J.; Mendel, Z. Biological control of the eucalyptus gall wasp Ophelimus maskelli (Ashmead): Taxonomy and biology of the parasitoid species Closterocerus chamaeleon (Girault), with information on its establishment in Israel. Biol. Control 2007, 42, 196–206. [Google Scholar] [CrossRef]
- Souza, N.M.; Schröder, M.L.; Hayes, R.A.; Bello, J.E.; Nahrung, H.F. Cuticular hydrocarbons of Gonipterus weevils: Are there species differences? Chemoecology 2021, 31, 159–167. [Google Scholar] [CrossRef]
- Lea, A.M. Australian Curculionidae of the subfamily Gonipterides. Proc. R. Soc. Vic. 1927, 39, 76–112. [Google Scholar]
- Loch, A.D. Parasitism of the Eucalyptus weevil, Gonipterus scutellatus Gyllenhal, by the egg parasitoid, Anaphes nitens Girault, in Eucalyptus globulus plantations in southwestern Australia. Biol. Control 2008, 47, 1–7. [Google Scholar] [CrossRef]
- Ward, S.E.; Valente, C.; Gonçalves, C.; Polaszek, A. Rediscovery and redescription of Centrodora damoni (Girault) (Hymenoptera: Aphelinidae) from Australia, an egg parasitoid of Gonipterus spp. (Coleoptera: Curculionidae), after nearly a century. Biodivers. Data J. 2016, e7766. [Google Scholar] [CrossRef] [Green Version]
- Valente, C.; Afonso, C.; Gonçalves, C.I.; Alonso-Zarazaga, M.A.; Reis, A.; Branco, M. Environmental risk assessment of the egg parasitoid Anaphes inexpectatus for classical biological control of the Eucalyptus snout beetle, Gonipterus platensis. BioControl 2017, 62, 457–468. [Google Scholar] [CrossRef]
- Stern, H.; De Hoedt, G.; Ernst, J. Objective classification of Australian climates. Aust. Met. Mag. 2000, 49, 87–96. [Google Scholar]
- Obinna, A.; Peter, N.; Korie, S. Niche separation between Encarsia dispersa and Encarsia guadeloupae, two biological control agents of the spiraling whitefly Aleurodicus dispersus, in Benin, West Africa. BioControl 2011, 56, 277–282. [Google Scholar] [CrossRef]
- Berger, J.; Jönsson, M.; Hedlund, K.; Anderson, P. Niche separation of pollen beetle parasitoids. Front. Ecol. Evol. 2015, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Pekas, A.; Tena, A.; Harvey, J.A.; Garcia-Marí, F.; Frago, E. Host size and spatiotemporal patterns mediate the coexistence of specialist parasitoids. Ecology 2016, 97, 1345–1356. [Google Scholar] [CrossRef]
- Santolamazza Carbone, S.; Cordero Rivera, A. Egg load and adaptive superparasitism in Anaphes nitens, an egg parasitoid of the Eucalyptus snout-beetle Gonipterus scutellatus. Entomol. Exp. Appl. 2003, 106, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Santolamazza Carbone, S.; Pestaña Nieto, M.; Cordero Rivera, A. Egg resorption behaviour by the solitary egg parasitoid Anaphes nitens under natural conditions. Ent. Exp. Appl. 2008, 127, 191–198. [Google Scholar] [CrossRef]
- Santolamazza Carbone, S.; Pestaña Nieto, M.; Cordero Rivera, A. Maternal size and age affect offspring sex ratio in the solitary egg parasitoid Anaphes nitens. Entomol. Exp. Appl. 2007, 125, 23–32. [Google Scholar] [CrossRef]
- Santolamazza Carbone, S.; Rodríguez Illamola, A.; Cordero Rivera, A. Host finding and host discrimination ability in Anaphes nitens Girault, an egg parasitoid of the Eucalyptus snout-beetle Gonipterus scutellatus Gyllenhal. Biol. Control 2004, 29, 24–33. [Google Scholar] [CrossRef]
- Santolamazza-Carbone, S.; Cordero Rivera, A. Superparasitism and sex ratio adjustment in a wasp parasitoid: Results at variance with Local Mate Competition? Oecologia 2003, 136, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Valente, C.; Afonso, C.; Gonçalves, C.I.; Branco, M. Assessing the competitive interactions between two egg parasitoids of the Eucalyptus snout beetle, Gonipterus platensis, and their implications for biological control. Biol. Control 2019, 130, 80–87. [Google Scholar] [CrossRef]
- Mills, N.J. Parasitoid interactions and biological control. In 1st International Symposium on Biological Control of Arthropods; USDA Forest Service: Honolulu, HI, USA, 2003. [Google Scholar]
- Pedersen, B.S.; Mills, N.J. Single vs. multiple introduction in biological control: The roles of parasitoid efficiency, antagonism and niche overlap. Appl. Ecol. 2004, 41, 973–984. [Google Scholar] [CrossRef]
- Souza, N.M.; Lawson, S.A.; Nahrung, H.F. Gonipterus (Coleoptera: Curculionidae) in subtropical Australia: Host associations and natural enemies. Austral Entomol. 2021. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Direction | Region | Location of 3′ End | Reference | Sequence (5′-3′) |
|---|---|---|---|---|---|
| C1-J-2183 | F | CO1 | 2183 | Simon et al., 1994 | CAA CAT TTA TTT TGA TTT TTT GG |
| C1-N-2659c | R | CO1 | 2659 | Laffin et al., 2005 | ACT AAT CCT GTG AAT AAA GG |
| TL2-N-3014 | R | CO1 | 3014 | Simon et al., 1994 | TCC AAT GCA CTA ATC TGC CAT ATT A |
| Ron | F | CO1 | 1751 | Simon et al., 1994 | GGA TCA CCT GAT ATA GCA TTC CC |
| K698 | F | CO1 | 1460 | Simon et al., 1994 | TAC AAT TTA TCG CCT AAA CTT CAG CC |
| K741 1999 | R | CO1 | 2578 | Caterino and Sperling 1999 | TGG AAA TGT GCA ACT ACA TAA TA |
| Gon-F | F | CO1 | 2215 | Mapondera et al., 2012 | GGA GTA CTC GGG ATA ATT TAC G |
| G118RT | F | CO1 | 2120 | This study | GGA GGG GGT GAC CCT ATT T |
| G118RT | R | CO1 | 2778 | This study | AGT CCG AGT ATC GAC GAG GT |
| Gonipterus Species | Eucalyptus Species | Section | Subgenus |
|---|---|---|---|
| Gonipterus sp. n. 1 | Eucalyptus conspicua * | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 1 | Eucalyptus cinerea subsp. cinerea | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 1 | Eucalyptus crenulata * | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 1 | E. morrisbyi * | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 1, Gonipterus sp. n. 2, Gonipterus sp. n. 3, Gonipterus sp. 11 | Eucalyptus globulus | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 2 | Eucalyptus longifolia | Similaris | Symphyomyrtus |
| Gonipterus sp. n. 2 | E. nicholii | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 2 | Eucalyptus dunnii | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 2 | Eucalyptus benthamii | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 2 | E. scoparia | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 2, Gonipterus sp. n. 3 | E. viminalis | Maidenaria | Symphyomyrtus |
| Gonipterus sp. n. 2, Gonipterus sp. n. 4, Gonipterus sp. n. 6 | E. microcorys | Alveolata | |
| Gonipterus sp. n. 4, Gonipterus sp. 12, Gonipterus sp. n. 6 | E. propinqua | Latoangulatae | Symphyomyrtus |
| G. cinnamomeus | E. melanophloia * | Adnataria | Symphyomyrtus |
| Gonipterus sp. n. 7 | |||
| G. notographus | E. radiata subsp. radiata * | Aromatica | Eucalyptus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schröder, M.L.; Nahrung, H.F.; de Souza, N.M.; Lawson, S.A.; Slippers, B.; Wingfield, M.J.; Hurley, B.P. Distribution of Gonipterus Species and Their Egg Parasitoids in Australia: Implications for Biological Control. Forests 2021, 12, 969. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080969
Schröder ML, Nahrung HF, de Souza NM, Lawson SA, Slippers B, Wingfield MJ, Hurley BP. Distribution of Gonipterus Species and Their Egg Parasitoids in Australia: Implications for Biological Control. Forests. 2021; 12(8):969. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080969
Chicago/Turabian StyleSchröder, Michelle L., Helen F. Nahrung, Natalia M. de Souza, Simon A. Lawson, Bernard Slippers, Michael J. Wingfield, and Brett P. Hurley. 2021. "Distribution of Gonipterus Species and Their Egg Parasitoids in Australia: Implications for Biological Control" Forests 12, no. 8: 969. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080969