
Roosting Behavior of Northern Long-Eared Bats (Myotis septentrionalis) in an Urban-Adjacent Forest Fragment

Department of Biological Sciences, Purdue University Fort Wayne, Fort Wayne, IN 46805, USA
*
Author to whom correspondence should be addressed.
Forests 2022, 13(12), 1972; https://0-doi-org.brum.beds.ac.uk/10.3390/f13121972
Submission received: 21 October 2022
/
Revised: 16 November 2022
/
Accepted: 20 November 2022
/
Published: 22 November 2022
(This article belongs to the Special Issue Biodiversity Conservation in Forest Fragments)
Abstract
:Throughout the Midwest United States, agricultural and urban development have fragmented natural areas, with a disproportionate effect on forests and wetlands. The resulting habitat loss, compounded with the spread of white-nose syndrome (WNS), has caused precipitous population declines in several forest-obligate bat species. In 2019, we discovered a remnant northern long-eared bat (Myotis septentrionalis Trouessart) maternity colony in a small forest fragment adjacent to a restored wetland in northeastern Indiana, USA. We investigated roost selection in this colony during the summers between 2019 and 2021 by attaching radio transmitters to northern long-eared bats and tracking them to day roosts. We measured tree, plot, and landscape-level characteristics for each roost and for a randomly selected available tree in the same landscape, then compared characteristics using paired t-tests. Over 70 net nights, we captured and tracked 4 individuals (1 juvenile male, 1 post-lactating female, and 2 lactating females) to 12 different roosts. There were, on average, 3.5 times more standing dead trees (snags) in plots around roosts compared to available trees (t = −4.17, p = 0.02). Bats in this maternity colony selected roosts near a stretch of flooded forest (which contained 83% of roosts) dominated by solar-exposed, flood-killed snags. These roosts likely provide warm microclimates that facilitate energy retention, fetal development, and milk production. By describing roosts within this landscape, we provide insight into the resources that enable an endangered bat species to persist in urbanized forest fragments.
1. Introduction
As human populations grow, urban and exurban expansion continues to replace forest land cover and impact forest communities [1,2,3]. Outside of urban and suburban areas, agricultural intensification is further fragmenting forests [4,5,6,7]. Despite successful forest restoration efforts in the East and Midwest United States [8], forest land cover is currently 25% lower than estimates of historic coverage [9]. Additionally, remnant forest patches are younger, smaller, and compositionally different than forests established prior to European settlement [10,11,12]. These changes to forest structure pose serious threats to organisms that evolved in the context of heavily wooded landscapes [13,14]. Forests offer critical sources of shelter and food for a wide range of species, whose activities reciprocally benefit the forest, nearby land cover types, and regional economies [15,16,17,18,19]. For example, insectivorous bats regulate populations of arthropods that can carry disease, damage crops, and infest native plant species [20,21,22]. By retaining forest patches that support bats, metropolitan areas will benefit more from the economic and ecological services that bats provide [23].
Many North American bat species rely on forests, which offer important foraging and roosting resources [24,25,26]. Studies in agricultural and urban areas have demonstrated that bat foraging activity and species richness increase in areas with more forested land and native plant communities [23,27,28]. Bats also take advantage of roosting opportunities in forests, using tree cavities, peeling bark, and bundles of dead leaves as daytime refugia throughout the summer [29]. Tree roosts protect bats from predators and inclement weather [30,31], facilitate important social interactions [32,33,34], and promote efficient thermoregulation [35,36]. For example, warmer roosts in buildings or solar-exposed snags (standing-dead trees) help bats remain normothermic while reducing heat loss to the environment [37,38]. The benefits of warm roosts with stable temperatures are especially important for colonies of pregnant and lactating bats, asentering torpor comes at the cost of delayed parturition and reduced milk production [39,40]. Conversely, bats that are not rearing young often select cooler roosts, such as shaded trees, to facilitate entry into torpor and significantly reduce overall energy expenditure [41,42,43]. Because roost selection in bats varies depending on species and the energetic needs of individuals, it is important to conserve forests with a diversity of roost types.
In the United States, forest-dependent bat species have suffered significant population declines due to habitat loss and fragmentation [44,45,46]. These negative impacts have been amplified by the spread of white-nose syndrome (WNS), a disease caused by the fungal pathogen Pseudogymnoascus destructans (Blehert and Gargas), which has increased winter mortality rates for many hibernating bat species in North America [47,48]. As a result, several formerly abundant bat species are listed as endangered at state and federal levels. Conservation strategies that focus on improving recruitment and survival of bat populations during spring and summer can help to offset WNS-related mortality during hibernation [49,50]. The continued conservation of large, remote forests will provide bats and other forest-dependent species the best chance of maintaining healthy populations. However, as urban and suburban developments expand farther into rural areas, it will also be important to identify aspects of urban forests that support imperiled species and develop management strategies that facilitate species diversity.
Northern long-eared bats (Myotis septentrionalis) are one of the forest-dependent species currently experiencing population declines in North America due to habitat loss and WNS [51,52,53]. Northern long-eared bats are considered endangered in the state of Indiana and are currently under consideration for classification as federally endangered in the United States [54]. A total of 11 northern long-eared bats were captured in the state of Indiana during the summers of 2017 and 2018; compared to an average of 55.6 ± 14.7 bats per summer (total 278 bats) from 2012–2016 [55]. In 2019, only two bats were captured in the state. Both bats were captured at Fox Island County Park, part of a forest/wetland complex located on the fringes of the second largest city in Indiana, Fort Wayne [56]. The presence of a northern long-eared bat population in an exurban nature preserve warrants investigation, especially since one individual was a reproductive female that we tracked to a roost in a large, solar-exposed snag just outside of the largest nearby forest patch, a roost type more characteristic of Indiana bats in the Midwest United States [57,58]. Many studies have speculated that northern long-eared bats select more solar-exposed roosts when faced with increased energetic demands, such as those associated with reproduction [53,59,60]. However, few studies have directly studied northern long-eared bat body temperature and torpor use, especially in relation to the characteristics of roost trees [61,62]. There are also few studies describing habitat use by northern long-eared bats in urban areas, likely because they are uncommon in areas lacking large patches of forest [63,64]. Northern long-eared bats have been reported in a 348 ha forest patch in Toronto, Canada and a 248 ha forest patch in Long Island, New York; however, in both cases their roost selection and foraging activity was limited to forest edges and interiors [65,66].
Over the summers of 2020 and 2021, we conducted research with the goal of describing roost selection and torpor use in the remnant population of northern long-eared bats in Fort Wayne, Indiana. We expected roost selection to follow trends identified in previous research, with bats roosting in cavities of live trees within forest fragments [65,67,68]. We also predicted that the main forest patch at Fox Island (approximately 245 ha) would be an important resource, given the results of research conducted in other urbanized areas [65,66]. Regarding torpor use, we expected bats roosting in trees with greater solar exposure to remain normothermic more often [41,69]. By describing the behavior of the colony identified at Fox Island, we will improve our understanding of how northern long-eared bats are impacted by urban development and habitat loss in a part of their range where this response has not yet been recorded. Studying roost preferences and torpor patterns in this population will provide insight into how northern long-eared bats might persist in urban-agricultural landscape mosaics.
2. Materials and Methods
2.1. Study Area
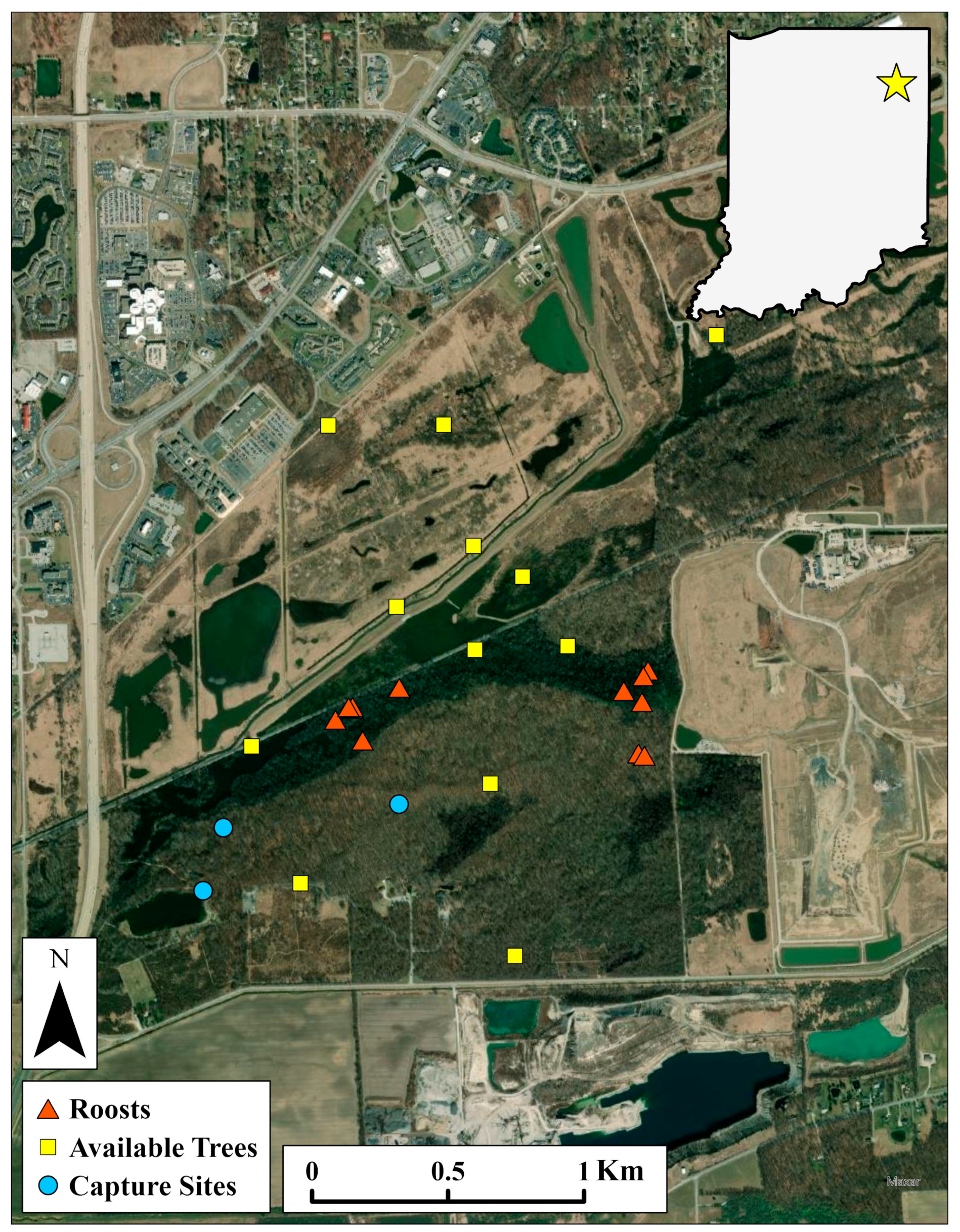
We conducted research in a complex composed of two adjacent nature preserves—Eagle Marsh Nature Preserve (335 ha; Eagle Marsh) and Fox Island County Park (245 ha; Fox Island)—located in an exurban area on the southwest periphery of Fort Wayne, IN, USA (Figure 1; 41.0192° N, 85.2373° W, Datum: WGS84). The two areas, though separated by railroad tracks, are essentially contiguous and have been managed through a partnership between the Little River Wetland Project, Allen County Parks, and the Indiana Department of Natural Resources. Eagle Marsh, located north of the railroad tracks, was converted from monocultural farmland in 2005 and is now an actively managed, restored wetland. Much of the area currently consists of sedge meadows, early and mid-successional grasslands, and wetlands [70]. Fox Island, located south of the railroad tracks, has been established as a county park and nature preserve since 1971. Landcover types within Fox Island include tallgrass prairies, vernal pools, a man-made lake, and a wooded sand dune. However, most of the park consists of contiguous, mixed deciduous forest. As a result of intense and permanent flooding in 2015, forest stands on the northern border of Fox Island became a 31 ha wetland with a large number of mature snags (primarily silver maples; Acer saccharinum L.; hereafter referred to as “snag forest”). Together, Eagle Marsh and Fox Island create a large, continuous mosaic of forest and wetland that is otherwise uncommon in Allen County, IN. Dominant tree species throughout the complex include silver maple, oaks (Quercus L. spp.), and eastern cottonwood (Populus deltoides W. Bartram ex Marshall) [70]. Permanent and ephemeral water sources are available throughout the complex in the form of shallow wetlands, ponds, lakes, and large sections of the Graham McCulloch Ditch.
The area surrounding the preserve complex is heavily modified, either by urban areas to the north and east, or by agricultural land to the south and west. A rock quarry and a landfill are immediately adjacent to the complex on the south and east side, respectively. A large interstate highway (I-69) passes immediately west of the complex and powerlines cut directly through the northern section of Eagle Marsh. The greater surrounding landscape consists of privately-owned farmland to the south and west, urban and suburban development to the north and east, and Fort Wayne International Airport to the southeast. The average daily temperature from 15 May to 15 August was 22.3 ± 0.4 °C (minimum–maximum = 16.2–28.4 °C) in 2020 and 22.0 ± 0.4 °C (minimum–maximum = 16.5–27.4 °C) in 2021. Total precipitation over the same period was 285 mm in 2020 and 392 mm in 2021 (National Weather Service Station Coop ID: 123037) [71].
2.2. Data Collection
We captured bats using mist-nets at eight sites throughout the study area from May–August 2019–2021. To optimize capture success, we deployed mist-nets across flight corridors within forests (e.g., trails and roads) or within natural forest openings [72]. For all bats caught, we recorded species, sex, age (adult or juvenile, as determined by development of the epiphyses of finger bones) [73], body mass (g), forearm length (mm), and reproductive status (females: pregnant, lactating, post-lactating, or non-reproductive; males: scrotal or non-reproductive) [72]. We attached lipped metal bands with unique codes around the right forearm of each bat before release to identify recaptured individuals. For every northern long-eared bat we captured, we trimmed the fur between the scapulae and attached temperature-sensitive radio transmitters (model: LB-2X, Holohil Systems Ltd., Carp, ON, Canada; 0.33 g) to the exposed skin using latex surgical adhesive (Perma-Type Company, Inc., Plainville, CT, USA). We ensured that radio transmitters weighed ≤ 5% of the body mass of captured bats [74]. All bats were handled following the American Society of Mammalogists guidelines [75] and Purdue University Animal Care and Use (IACUC Protocol # 1902001857). All fieldwork was conducted under federal (TE70488C) and state (2667) permits held by S.M. Bergeson.
Following transmitter attachment, we followed radio-tagged bats to day roosts using TRX-2000 WR telemetry receivers (Wildlife Materials, Inc., Murphysboro, IL, USA) and a 3- or 5-element Yagi antenna (Wildlife Materials, Inc., Murphysboro, IL, USA). We tracked bats every day until transmitters fell off or failed (e.g., battery death) or the bat left the landscape. We deployed telemetry data loggers (model: SRX-800, Lotek Wireless, Newmarket, ON Canada) affixed with 3-element Yagi antennas in the vicinity of occupied roosts to record radio pulses throughout the lifespan of the transmitter. We converted the radio pulse data to bat skin temperature (Tsk), accurate to the nearest 0.1 °C, using calibration curves provided by the manufacturer. To estimate roost colony size and confirm the presence of tracked bats in roosts, we conducted emergence counts for every roost [76]. We analyzed roost fidelity by calculating the mean number of roost-switching events per tracking period for each bat (1 event = 1 day in which a bat used a roost it did not use the day before).
We characterized located day roosts by calculating roost tree condition (live or dead), level of snag decay (1–4) [77], tree height (m), roost entrance height (m), roost aspect (°), and roost canopy cover (%). We also characterized plot-scale roost characteristics in a 0.1 ha circular plot centered on the roost tree. In each plot, we recorded canopy cover (%) and the number of trees/snags surrounding the roost. Roost and plot canopy cover were determined visually to the nearest 25% for each of four quadrants, then averaged across all four quadrants for a final value [78]. Finally, we used ArcGIS (ESRI 2022, ArcGIS Version 10.8.1, Redlands, CA: Environmental Systems Research Institute) to measure landscape-level roost characteristics, including proximity to water (m), proximity to forest edge (m), proximity to edge of the snag forest (m), and proximity to edge of the largest forest patch (m). Additionally, we calculated minimum convex polygons (MCPs) to obtain an average roost area (ha) for individuals that used ≥ 3 roosts and an overall colony roost area (ha) based on the roosts that we identified from 2019–2021.
To assess roost selection, we characterized one available tree for every confirmed roost tree. We assumed that these trees were unused by northern long-eared bats, as tagged bats were never observed using them. To locate available trees, we used ArcGIS to select a random point within a 0.9 ± 0.4 km radius (average distance between roosts and capture sites) of each roost. From the random point, we located the nearest tree with a DBH ≥ 10 cm (i.e., larger than a sapling) and in the same condition as the corresponding roost tree (live or dead) [78,79]. We measured the same plot and landscape-level characteristics for available trees as for observed roost trees, excluding roost height and roost aspect.
Several temperature-sensitive radio transmitters used in this study fell off tagged bats but remained lodged in roosts, as determined by vertical telemetry triangulation. We assumed that these transmitters were within the area that bats used to roost, although it is possible that transmitters had fallen below the roost area but remained within the roost tree interior. Given that we did not observe radio transmitters on the ground or the exterior of roost trees, the data we collected from the transmitters after they were dropped may be a valuable indicator of roost microclimates. Thus, we used temperature data collected during these periods to measure the internal temperature of roosts (Ttree) and compared it to ambient temperature (Ta) collected from the Fort Wayne Airport.
2.3. Data Analysis
We measured Tsk as a proxy for internal body temperature (Tb) to study daily torpor use [80]. We considered Tsk data in analyses only if there were measurements throughout a full roost day, which we defined as the time the radio-tagged bat entered the roost in the morning to the time it left in the evening. For each full roost day, we collected data on the number of torpor bouts, torpor duration (min), and torpor depth (°C; minimum Tsk while torpid). We considered a radio-tagged bat to be using torpor any time its Tsk dropped below a torpor onset threshold (TOT) for at least 10 consecutive minutes. This TOT was calculated using an equation developed by Willis [81] for calculating a torpor onset threshold minus one standard error (TOT − 1 SE) in small heterotherms:
For the data collected on roosts and available trees, we used a Shapiro–Wilk test to check for normality (W ≥ 0.9) and transformed any data that did not fit a normal distribution using either a logarithmic, square root, or arcsin square root transformation. If either the original or transformed data were normally distributed, we performed paired t-tests to compare characteristics of roosts and available trees, pairing roosts with their corresponding available trees. Otherwise, we performed Wilcoxon matched pairs signed-rank tests on the original data. We used the Benjamini–Hochberg method to maintain a false discovery rate of ≤5% associated with making multiple comparisons of the same paired data [82]. Finally, we used a two-tailed Fisher’s exact test to assess the selection of roost tree species by northern long-eared bats. All statistical tests were performed in R (Version 4.1.1) [83] with 95% confidence intervals (α = 0.05), and data are reported as mean ± standard error.
3. Results
Our overall survey effort included 104 net nights (1 net night = 1 mist-net set up on 1 night) over 30 calendar nights across eight sites. We captured 172 bats total, of which 84% were big brown bats (Eptesicus fuscus Palisot de Beauvois), 10% were red bats (Lasiurus borealis Müller), 3% were northern long-eared bats (Myotis septentrionalis), and 2% were silver-haired bats (Lasionycteris noctivagans Le Conte). Out of the five northern long-eared bats we captured, we tracked four individuals (one non-reproductive juvenile male, one lactating female, and two post-lactating females) to 12 roost trees throughout the study area. We captured two northern long-eared bats in 2019 (one non-reproductive adult male and one lactating female), but we only tracked the female due to a limited number of radio transmitters. We did not collect body temperature data on the female we tracked, but we identified one roost that it used and included that in our dataset. In 2021, we tracked two post-lactating bats, one of which we tracked over two separate periods seven days apart. We tracked bats for an average of 5.5 ± 1.2 days (range = 2–7 days) before they dropped their radio transmitters, a time frame consistent with other work on northern long-eared bats [60,65,84]. Bats switched roosts an average of 3.0 ± 1.2 times throughout a tracking period (0.5 ± 0.2 switching events/days tracked). The average roost area for individual bats was 0.53 ± 0.52 ha (range = 0.004–1.576 ha; n = 3). Overall, including roosts from 2019–2021, the colony had a total roost area of 26.4 ha (1.5 ± 0.1 ha per year).
We conducted 21 emergence counts on 11 of the 12 identified roosts (1–4 per roost) and observed at least one bat leaving a roost on nine occasions. We conducted emergence count surveys on roosts up to seven days after they were first identified, although bats were only confirmed exiting roosts up to three days after roosts were first identified. We never directly observed bats emerging from three of the roosts we surveyed. In one of these cases, the transmitter signal moved away from the identified tree at 21:40, suggesting the bat left but was not observed. In the other two cases, the transmitter signal came consistently from the identified tree and did not move throughout the life of the transmitter, suggesting the transmitter had detached from the tracked bat. The emergence counts we conducted indicate that bats roosted solitarily or in small groups (average = 2 bats/roost/emergence, range = 1–8 bats/roost/emergence).
The majority of roosts (83%) were under exfoliating bark on snags within the boundaries of the snag forest, while the remaining roosts were located under exfoliating bark on live trees in the Fox Island forest interior. However, these live trees were still <200 m from the edge of the snag forest. The bats we tracked showed a strong preference for roosting in silver maples compared to other tree species available in their foraging range (p < 0.01; Figure 2). Of the 12 roosts used, 11 were silver maples and one was an eastern cottonwood. The tree-level characteristics we measured did not differ significantly between roost and available trees, although roost trees were slightly taller than available trees on average (Table 1). At the plot-level, plots surrounding roost trees had 3.6 times more snags (t = −4.17, p = 0.02) compared to plots surrounding available trees. At the landscape-level, roosts were located 16 times closer to the snag forest (V = 55.0, p = 0.03) than available trees. Due to the low number of bats tracked in this study, we did not compare roost selection between bats based on sex or reproductive stage. However, it should be noted that female bats were tracked exclusively to snags within the snag forest. The juvenile male tracked in this study used several snags in the snag forest, but also used two live trees in the Fox Island forest nearby.
Due to technical issues deploying dataloggers and the short lifespan of transmitters, we were only able to collect three full days of Tsk data from a juvenile male tracked in late July 2020. This individual spent the first day in a live silver maple in the Fox Island forest interior, and the next two days in a solar-exposed snag in the northeastern portion of the snag forest. Because we were only able to collect sufficient Tsk data on this juvenile male bat, the TOT value calculated (32.3 °C) is specific to that individual. We subtracted 2 °C from this value to obtain the final TOT value (30.3 °C) as Tsk is approximately 2 °C lower than Tb on average [80,85]. The juvenile male we studied remained normothermic while roosting until the third day, during which it conducted five separate torpor bouts, spent 130 min torpid, and had an average torpor depth of 28.3 ± 0.5 °C (minimum temperature while torpid; range = 26.4–29.7 °C). Specifically, the bat conducted three torpor bouts in the early morning (between 05:45 and 06:45) and two in the late afternoon (between 16:30 pm and 19:00 pm). Average torpor bout length was 26.0 ± 9.8 min (range = 12–65 min).
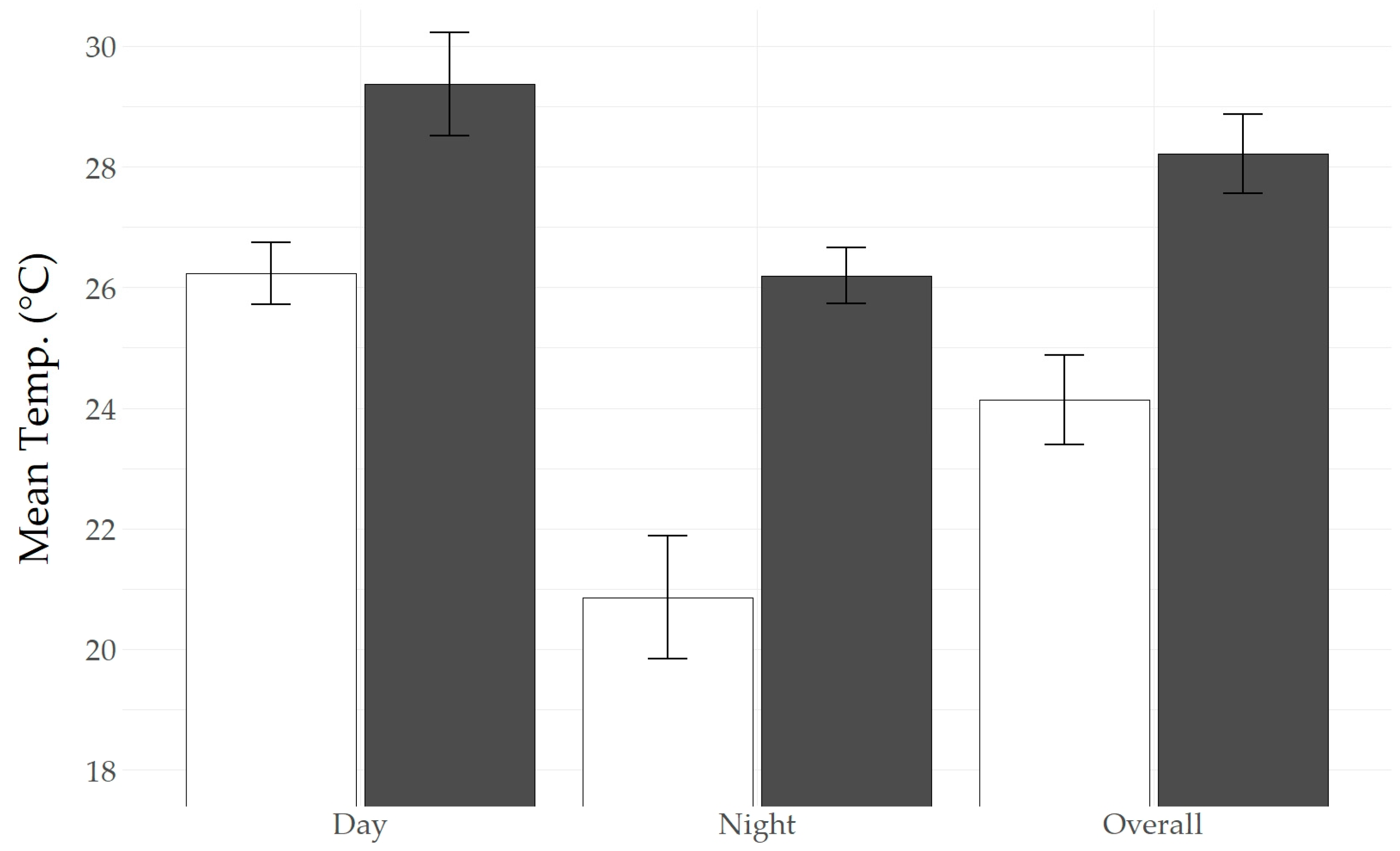
Four of the transmitters in our project detached from tracked bats and remained lodged in the last-used roost tree. We collected an average of 5.5 ± 1.5 days of Ttree data per roost tree (range = 2–8 days). Roosts included a live eastern cottonwood within the Fox Island forest used in July of 2020 (bat likely roosted in a tree crevice), two silver maple snags in the snag forest (bats used exfoliating bark), both surveyed simultaneously in June of 2021, and another silver maple snag in the snag forest surveyed later the same month (bat used a tree cavity). Given the small sample size (n = 4), we could not perform meaningful statistical analyses to assess trends in roost temperatures. Rather, we report descriptive statistics of hourly Ttree compared to Ta during the same period. Average Ttree was warmer than Ta for all four roost trees containing dropped transmitters (average difference between Troost and Ta = 4.0 ± 1.4 °C; range = 0.4–6.1 °C; Figure 3). The temperature difference between Ttree and Ta was more pronounced at night (21:54–06:54; average difference = 5.4 ± 0.7 °C; range = 1.9–8.1 °C) and less extreme during the day (06:54–21:54; average difference = 3.1 ± 0.7 °C; range = −0.4–5.4 °C). Daytime roost tree temperatures averaged 29.4 ± 0.9 °C (average maximum = 36.3 ± 1.5 °C; average minimum = 22.0 ± 1.4 °C; range = 18.5–38.9 °C), whereas the average Ta during the same period was 26.2 ± 0.5 °C (average maximum = 31.9 ± 0.8 °C; average minimum = 18.1 ± 2.5 °C; range = 12.2–32.8 °C).
4. Conclusions
Contrary to our prediction that northern long-eared bats would use shaded trees in the forest interior, selected roosts were predominantly under exfoliating bark of large, solar-exposed snags within the 31 ha flooded snag forest. Individuals in this study selected roost trees based on plot- and landscape-level characteristics associated with the snag forest. We did not identify any tree-level characteristics that were significantly different between roosts and available trees. This is consistent with previous reports, which suggest that northern long-eared bats are fairly flexible in their roosting habits, selecting trees of various sizes and stages of decay, so long as they are situated within forest plots and offer suitable cavities or exfoliating bark [68,86]. Northern long-eared bats also roost in a wide range of tree species depending on local forest composition, often choosing species in proportion to their abundance [60,87,88,89]. Such behavior may offer an explanation as to why the bats in our study roosted almost exclusively in silver maples. These bats may be taking advantage of a sudden abundance of snags created by the flooding of a forest patch in which silver maples were the dominant species. As our sample of bats was limited, our results may not reflect roost selection for the entire population. However, if the maximum exit count of 8 bats is an appropriate estimate of population size, our sample size may be adequate to make interpretations for this small, remnant population.
While the characteristics of specific roost trees in our study were variable, plots surrounding roost trees had consistently more snags than those surrounding available trees. This trend is likely related to the conditions in the snag forest, where nearly all standing trees are snags. Because of the lack of live foliage, roosts in the snag forest were also heavily solar-exposed. Despite this, canopy cover was not statistically different between roosts and available trees. This may be a result of small sample size, or a product of randomly selecting available trees based on circular radii that were calculated using the short flight distances from capture site to first roost site (typical of northern long-eared bats) [84,90]. Most of the available trees were situated in the snag forest or in the northern wetland, where canopy cover is comparable to that of the roost trees. Because snags with low canopy cover were not limited within this landscape, bats likely focused their roosting activity in areas with other desirable and more limited characteristics, such as a high density of available roosts. The high abundance of potential roosts within the snag forest is, itself, a beneficial characteristic. High roost availability promotes roost-switching behavior [91], which reduces the risk of roost predation [92], allows for flexibility in thermal refugia choice [41], and facilitates fission–fusion social group dynamics [93,94]. The bats in our study area switched roosts often, suggesting that this feature may be one of the reasons the colony remained in the snag forest. Additionally, the hydric nature of the snag forest and its proximity to a large forest patch likely provides an optimal foraging habitat for northern long-eared bats [90,95,96].
Although solar-exposed trees were not limited in our study area, solar exposure is likely still important for northern long-eared bat maternity colonies. Trees in areas with lower levels of canopy cover offer warmer microclimates, which can reduce the cost of remaining normothermic during energetically expensive periods of gestation and lactation [40,42,97]. Alternatively, shaded stands offer cooler, more stable temperatures that facilitate entry into torpor and allow non-reproductive bats to conserve energy [41,69,98]. The solar-exposed roosts in the snag forest were warmer on average than ambient conditions, which potentially makes them a critical resource for the success of pregnant or lactating bats and their offspring. The only bat to use shaded trees in the forest interior was a juvenile, non-reproductive male. However, this individual also used several solar-exposed snags resembling those used by lactating and post-lactating females. Although our sample size was not large enough to make statistical inferences about roost use based on sex or reproductive condition, the fact that northern long-eared bats in various reproductive stages used the same stretch of snags suggests it is an important resource for the entire remnant population.
The roost preferences of the maternity colony described in this study differ from preferences reported in other studies in the Midwest United States, where northern long-eared bats preferentially roosted in cavities of live trees and snags in forest interiors [60,99,100]. The roosts used in our study have characteristics more typical of roosts used by Indiana bat maternity colonies in the region [57,101,102]. Whereas Indiana bats are heavily reliant on exfoliating bark of solar-exposed snags, northern long-eared bats select a mixture of shaded live and dead trees [67,78]. However, there is still overlap in roost preferences, as both species tend to select trees in areas with relatively open canopies compared to the surrounding forest [88,103]. No Indiana bats have been captured in our study area, despite three consecutive years of netting [56]. Timpone et al. [104] note that northern long-eared bats in areas outside of the known distribution of Indiana bats roost more frequently in snags than in areas where the two species co-occur. This maternity colony of northern long-eared bats may be using a wider range of roosts in the absence of competition with Indiana bats. Alternatively, if Indiana bats are present but undetected, this snag forest may provide such an abundance of high-quality, solar-exposed roosts that interspecific competition and niche partitioning between the two species need not occur [86,105].
Northern long-eared bats are less common in areas with greater urban land-use [90,106]. Although the exact reasons for such rarity in developed areas are not reported, the depletion of forest and wetland resources in urbanized landscapes [1] and the introduction of anthropogenic stressors such as noise and light pollution [107,108] likely play a role in their exclusion. However, healthy reproductive populations have been reported in heavily forested parks within large cities including Long Island, New York; Washington, D.C; Toronto; Canada [64,65,66]. In these studies, the forest fragments ranged in size from 248–710 ha and contained permanent sources of water, such as rivers and ponds. Northern long-eared bats in these sites roosted and foraged exclusively along forest edges and in the forest interior, and used anthropogenic roost sources (e.g., houses, utility poles) infrequently, if at all [65,66]. Thus, it is improbable that northern long-eared bats would be present in urban areas without one or more large forest patches, where natural roost options are readily available. The population we studied may be similarly restricted to areas near forests with sufficient natural roost options. Although the snag forest in which bats roosted may not be considered forest interior, it stretches along the northern boundary of Fox Island, the largest forest patch in the nearby landscape, which also contains permanent sources of water.
It is clear from our results that the northern long-eared bat population in our site is taking advantage of abundant solar-exposed snags created by a permanent flooding event in 2015. Because we did not conduct surveys of the area prior to this flood, it is unknown whether northern long-eared bats were present in the study area beforehand or if they began roosting there only after the development of these snags. Regardless, the use of this feature by reproductive females and their offspring suggests that it is a valuable resource for a remnant maternity colony of a state-endangered species. However, the snags currently available will not sustain bat populations indefinitely. In early June 2022, over 1000 trees in the Fox Island forest and many of the trees in the snag forest were toppled due to a derecho storm that passed through the region. Over the next 20 to 30 years, as more snags fall due to severe weather and natural decay, the number of standing trees with exfoliating bark or suitable cavities will dwindle [109,110]. Whether northern long-eared bats will shift their roosting behavior into the forest interior or abandon the study area entirely is not clear.
The findings of this study may provide guidance for management to support bat diversity within or near urban areas. Maintaining contiguous patches of wetlands and mature woodlands can attract forest-dependent species that might otherwise struggle. Within forests, it is important to promote roost trees with a range of microclimates, including solar-exposed snags, which benefit maternity colonies and are important for the recruitment of new individuals in a population. In the case of the snag forest and similar flooded woodlands, bats will benefit from efforts to maintain old snags and recruit new snags to replace them. The benefits of maintaining forests with riparian foraging habitat and an assortment of roost types will not only help forest-adapted species; generalist bat species that are more tolerant of urban areas will also benefit from increased habitat heterogeneity and a wider selection of roosting sites.
Author Contributions
Conceptualization, G.E.B. and S.M.B.; methodology, G.E.B. and S.M.B.; validation, S.M.B.; formal analysis, G.E.B. and S.M.B.; investigation, G.E.B. and S.M.B.; resources, S.M.B.; data curation, G.E.B.; writing—original draft preparation, G.E.B.; writing—review and editing, G.E.B. and S.M.B.; visualization, G.E.B. and S.M.B.; supervision, S.M.B.; project administration, S.M.B.; funding acquisition, G.E.B. and S.M.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Fox Island Alliance, Little River Wetlands Project, Purdue University Fort Wayne Research and Innovation, and the Indiana Space Grant Consortium (INSGC).
Data Availability Statement
The data presented in this study are openly available in Open Science Framework (OSF) at https://0-doi-org.brum.beds.ac.uk/10.17605/OSF.IO/PKA9T (20 October 2022).
Acknowledgments
We thank The Fox Island Alliance and Little River Wetlands Project for providing numerous volunteers. We thank the Little River Wetlands Project and the Allen County Parks Department for site access. We also thank the numerous volunteers that helped collect data in the field.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Bagan, H.; Yamagata, Y. Land-cover change analysis in 50 global cities by using a combination of Landsat data and analysis of grid cells. Environ. Res. Lett. 2014, 9, 064015. [Google Scholar] [CrossRef]
- Brown, D.G.; Johnson, K.M.; Loveland, T.R.; Theobald, D.M. Rural land-use trends in the conterminous United States, 1950–2000. Ecol. Appl. 2005, 15, 1851–1863. [Google Scholar] [CrossRef] [Green Version]
- Shifley, S.R.; Moser, W.K.; Nowak, D.J.; Miles, P.D.; Butler, B.J.; Aguilar, F.X.; DeSantis, R.D.; Greenfield, E.J. Five anthropogenic factors that will radically alter forest conditions and management needs in the northern United States. For. Sci. 2014, 60, 914–925. [Google Scholar] [CrossRef]
- Petit, S.; Firbank, L.; Wyatt, B.; Howard, D. MIRABEL: Models for integrated review and assessment of biodiversity in European landscapes. Ambio 2001, 30, 81–88. [Google Scholar] [CrossRef]
- Billeter, R.; Liira, J.; Bailey, D.; Bugter, R.; Arens, P.; Augenstein, I.; Aviron, S.; Baudry, J.; Bukacek, R.; Burel, F.; et al. Indicators for biodiversity in agricultural landscapes: A pan-European study. J. Appl. Ecol. 2007, 45, 141–150. [Google Scholar] [CrossRef]
- Foley, J.A.; Defries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [Green Version]
- Matson, P.A.; Parton, W.J.; Power, A.G.; Swift, M.J. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.; Shao, G.; Swihart, R.; Parker, G.; Tang, L. Implications of shared edge length between land cover types for landscape quality: The case of Midwestern US, 1940–1998. Landsc. Ecol. 2008, 23, 391–402. [Google Scholar] [CrossRef]
- Smith, W.B.; Darr, D.U.S. Forest Resource Facts and Historical Trends; FS-801; United States Department of Agriculture Forest Service: Washington, DC, USA, 2004; pp. 7–10.
- Goring, S.J.; Mladenoff, D.J.; Cogbill, C.V.; Record, S.; Paciorek, C.J.; Jackson, S.T.; Dietze, M.C.; Dawson, A.; Matthes, J.H.; McLachlan, J.S.; et al. Novel and lost forests in the upper Midwestern United States, from new estimates of settlement-era composition, stem density, and biomass. PLoS ONE 2016, 11, e0151935. [Google Scholar] [CrossRef] [Green Version]
- Riitters, K.H.; Wickham, J.D.; O’Neill, R.V.; Jones, K.B.; Smith, E.R.; Coulston, J.W.; Wade, T.G.; Smith, J.H. Fragmentation of continental United States forests. Ecosystems 2002, 5, 815–822. [Google Scholar] [CrossRef]
- Thompson, J.R.; Carpenter, D.N.; Cogbill, C.V.; Foster, D.R. Four centuries of change in northeastern United States forests. PLoS ONE 2013, 8, e72540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betts, M.G.; Phalan, B.; Frey, S.J.K.; Rousseau, J.S.; Yang, Z. Old-growth forests buffer climate-sensitive bird populations from warming. Divers. Distrib. 2018, 24, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, L.; Betts, M.; Forbes, G.; Vernes, K. Effects of landscape composition and configuration on northern flying squirrels in a forest mosaic. For. Ecol. Manag. 2009, 257, 1920–1929. [Google Scholar] [CrossRef]
- Carey, A.B. Sciurids in Pacific Northwest managed and old-growth forests. Ecol. Appl. 1995, 5, 648–661. [Google Scholar] [CrossRef] [Green Version]
- Siepielski, A.M.; Benkman, C.W. A seed predator drives the evolution of a seed dispersal mutualism. Proc. R. Soc. B Biol. Sci. 2008, 275, 1917–1925. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Hanula, J.L.; Ulyshen, M.D.; Horn, S. Conserving pollinators in North American forests: A review. Nat. Areas J. 2016, 36, 427–439. [Google Scholar] [CrossRef]
- Boesing, A.L.; Nichols, E.; Metzger, J.P. Effects of landscape structure on avian-mediated insect pest control services: A review. Landsc. Ecol. 2017, 32, 931–944. [Google Scholar] [CrossRef]
- Blažek, J.; Konečný, A.; Bartonička, T. Bat aggregational response to pest caterpillar emergence. Sci. Rep. 2021, 11, 13634. [Google Scholar] [CrossRef]
- Charbonnier, Y.; Barbaro, L.; Theillout, A.; Jactel, H. Numerical and functional responses of forest bats to a major insect pest in pine plantations. PLoS ONE 2014, 9, e109488. [Google Scholar] [CrossRef] [PubMed]
- Reiskind, M.H.; Wund, M.A. Experimental assessment of the impacts of northern long-eared bats on ovipositing Culex (Diptera: Culicidae) mosquitoes. J. Med. Entomol. 2009, 46, 1037–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.; Gehrt, S. Bat response to woodland restoration within urban forest fragments. Restor. Ecol. 2010, 18, 914–923. [Google Scholar] [CrossRef]
- Lacki, M.J.; Hayes, J.P.; Kurta, A. Bats in Forests: Conservation and Management; Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 1–329. [Google Scholar]
- Lima, S.L.; O’Keefe, J.M. Do predators influence the behaviour of bats? Biol. Rev. Camb. Philos. Soc. 2013, 88, 626–644. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, E.V.; Laine, T.; Morgan, S.E.; Cole, K.R.; Speakman, J.R. Roost selection in the pipistrelle bat, Pipistrellus pipistrellus (Chiroptera: Vespertilionidae), in northeast Scotland. Anim. Behav. 1998, 56, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Avila-Flores, R.; Fenton, M.B. Use of spatial features by foraging insectivorous bats in a large urban landscape. J. Mammal. 2005, 86, 1193–1204. [Google Scholar] [CrossRef]
- Heim, O.; Treitler, J.T.; Tschapka, M.; Knörnschild, M.; Jung, K. The importance of landscape elements for bat activity and species richness in agricultural areas. PLoS ONE 2015, 10, e0134443. [Google Scholar] [CrossRef] [Green Version]
- Drake, E.C.; Gignoux-Wolfsohn, S.; Maslo, B. Systematic review of the roost-site characteristics of North American forest bats: Implications for conservation. Diversity 2020, 12, 76. [Google Scholar] [CrossRef] [Green Version]
- Kunz, T.H. Roosting ecology of bats. In Ecology of Bats; Kunz, T.H., Ed.; Springer US: Boston, MA, USA, 1982; pp. 1–55. [Google Scholar]
- Mager, K.J.; Nelson, T.A. Roost-Site Selection by Eastern Red Bats (Lasiurus borealis). Am. Midl. Nat. 2001, 145, 120–126. [Google Scholar] [CrossRef]
- Kerth, G.; Almasi, B.; Ribi, N.; Thiel, D.; Lüpold, S. Social interactions among wild female Bechstein’s bats (Myotis bechsteinii) living in a maternity colony. Acta Ethologica 2003, 5, 107–114. [Google Scholar] [CrossRef]
- Safi, K.; Kerth, G. Comparative analyses suggest that information transfer promoted sociality in male bats in the temperate zone. Am. Nat. 2007, 170, 465–472. [Google Scholar] [CrossRef]
- Wilkinson, G.S. Information transfer at evening bat colonies. Anim. Behav. 1992, 44, 501–518. [Google Scholar] [CrossRef] [Green Version]
- Sedgeley, J.A. Quality of cavity microclimate as a factor influencing selection of maternity roosts by a tree-dwelling bat, Chalinolobus tuberculatus, in New Zealand. J. Appl. Ecol. 2001, 38, 425–438. [Google Scholar] [CrossRef]
- Turbill, C. Roosting and thermoregulatory behaviour of male Gould’s long-eared bats, Nyctophilus gouldi: Energetic benefits of thermally unstable tree roosts. Aust. J. Zool. 2006, 54, 57–60. [Google Scholar] [CrossRef]
- Bergeson, S.B. Multi-Scale Analysis of Roost Characteristics and Behavior of the Endangered Indiana Bat (Myotis sodalis). Ph.D. Dissertation, Indiana State University, Terre Haute, IN, USA, 2017. [Google Scholar]
- Wilcox, A.; Willis, C.K.R. Energetic benefits of enhanced summer roosting habitat for little brown bats (Myotis lucifugus) recovering from white-nose syndrome. Conserv. Physiol. 2016, 4, cov070. [Google Scholar] [CrossRef] [Green Version]
- Besler, N.K.; Broders, H.G. Combinations of reproductive, individual, and weather effects best explain torpor patterns among female little brown bats (Myotis lucifugus). Ecol. Evol. 2019, 9, 5158–5171. [Google Scholar] [CrossRef] [Green Version]
- Wilde, C.J.; Knight, C.H.; Racey, P.A. Influence of torpor on milk protein composition and secretion in lactating bats. J. Exp. Zool. 1999, 284, 35–41. [Google Scholar] [CrossRef]
- Bergeson, S.M.; Brigham, R.M.; O’Keefe, J.M. Free-ranging bats alter thermoregulatory behavior in response to reproductive stage, roost type, and weather. J. Mammal. 2021, 102, 705–717. [Google Scholar] [CrossRef]
- Dzal, Y.A.; Brigham, R.M. The tradeoff between torpor use and reproduction in little brown bats (Myotis lucifugus). J. Comp. Physiol. B 2013, 183, 279–288. [Google Scholar] [CrossRef]
- Johnson, J.S.; Lacki, M.J. Effects of reproductive condition, roost microclimate, and weather patterns on summer torpor use by a vespertilionid bat. Ecol. Evol. 2014, 4, 157–166. [Google Scholar] [CrossRef]
- Frick, W.F.; Kingston, T.; Flanders, J. A review of the major threats and challenges to global bat conservation. Ann. New York Acad. Sci. 2020, 1469, 5–25. [Google Scholar] [CrossRef]
- Park, K.J. Mitigating the impacts of agriculture on biodiversity: Bats and their potential role as bioindicators. Mamm. Biol. 2015, 80, 191–204. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.L.; Reichard, J.D.; Coleman, J.T.H.; Weller, T.J.; Thogmartin, W.E.; Reichert, B.E.; Bennett, A.B.; Broders, H.G.; Campbell, J.; Etchison, K.; et al. The scope and severity of white-nose syndrome on hibernating bats in North America. Conserv. Biol. 2021, 35, 1586–1597. [Google Scholar] [CrossRef]
- Frick, W.F.; Pollock, J.F.; Hicks, A.C.; Langwig, K.E.; Reynolds, D.S.; Turner, G.G.; Butchkoski, C.M.; Kunz, T.H. An emerging disease causes regional population collapse of a common North American bat species. Science 2010, 329, 679–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, N.W.; McGuire, L.P.; Pannkuk, E.L.; Blute, T.; Haase, C.G.; Mayberry, H.W.; Risch, T.S.; Willis, C.K.R. Disease recovery in bats affected by white-nose syndrome. J. Exp. Biol. 2020, 223, jeb211912. [Google Scholar] [CrossRef]
- Hyzy, B.; Russell, R.; Silvis, A.; Ford, W.; Riddle, J.; Russell, K. Investigating maternity roost selection by northern long-eared bats at three sites in Wisconsin. Endanger. Species Res. 2020, 41, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Pettit, J.L.; O’Keefe, J.M. Impacts of white-nose syndrome observed during long-term monitoring of a Midwestern bat community. J. Fish Wildl. Manag. 2017, 8, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, R.J.; Powers, K.E.; Orndorff, W.; Ford, W.M.; Hobson, C.S. Changes in rates of capture and demographics of Myotis septentrionalis (northern long-eared bat) in western Virginia before and after onset of white-nose syndrome. Northeast. Nat. 2016, 23, 195–204. [Google Scholar] [CrossRef]
- Thalken, M.M.; Lacki, M.J. Tree roosts of northern long-eared bats following white-nose syndrome. J. Wildl. Manag. 2018, 82, 629–638. [Google Scholar] [CrossRef]
- Endangered and Threatened Wildlife and Plants; Endangered Species Status for Northern Long-Eared Bat; 87 Fed. Reg. 16442 (proposed March 23, 2022); Office of the Federal Register, National Archives and Records Administration: Washington, DC, USA, 2022.
- Westrich, B.J.; Indiana Department of Natural Resources, Indianapolis, IN, USA. Personal communication, 2022.
- Barton, A.P.; Miller, E.L.; Bergeson, S.M. Bat diversity survey of a nature preserve complex near Fort Wayne, Indiana. In Proceedings of the Indiana Academy of Science; Ruch, D.G., Rothrock, P., Eds.; IAS Publications: Indianapolis, IN, USA, 2020; Volume 129, pp. 115–123. [Google Scholar]
- Carter, T.C.; Feldhamer, G.A. Roost tree use by maternity colonies of Indiana bats and northern long-eared bats in southern Illinois. For. Ecol. Manag. 2005, 219, 259–268. [Google Scholar] [CrossRef]
- Bergeson, S.M.; Holmes, J.B.; O’Keefe, J.M. Indiana bat roosting behavior differs between urban and rural landscapes. Urban Ecosyst. 2020, 23, 79–91. [Google Scholar] [CrossRef]
- Garroway, C.J.; Broders, H.G. Day roost characteristics of northern long-eared bats (Myotis septentrionalis) in relation to female reproductive status. Écoscience 2008, 15, 89–93. [Google Scholar] [CrossRef]
- Silvis, A.; Ford, W.M.; Britzke, E.R.; Beane, N.R.; Johnson, J.B. Forest succession and maternity day roost selection by Myotis septentrionalis in a mesophytic hardwood forest. Int. J. For. Res. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Kaupas, L. Roosting Behaviour and Thermoregulation of the Northern Long-Eared Bat (Myotis septentrionalis) near the Northern Extent of Its Range. M.S. Thesis, University of Calgary, Calgary, AB, Canada, 2016. [Google Scholar]
- Sichmeller, T.J. Determining energy conservation during torpor for three Myotis species and response of Myotis species to human disturbance while day roosting. M.S. Thesis, Ball State University, Muncie, IN, USA, 2010. [Google Scholar]
- Owen, S.F.; Menzel, M.A.; Ford, W.M.; Chapman, B.R.; Miller, K.V.; Edwards, J.W.; Wood, P.B. Home-range size and habitat used by the northern myotis (Myotis septentrionalis). Am. Midl. Nat. 2003, 150, 352–359. [Google Scholar] [CrossRef]
- Johnson, J.; Gates, J.; Ford, W. Distribution and activity of bats at local and landscape scales within a rural–urban gradient. Urban Ecosyst. 2008, 11, 227–242. [Google Scholar] [CrossRef]
- Thorne, T.J.; Matczak, E.; Donnelly, M.; Franke, M.C.; Kerr, K.C.R. Occurrence of a forest-dwelling bat, northern myotis (Myotis septentrionalis), within Canada’s largest conurbation. J. Urban Ecol. 2021, 7, juab029. [Google Scholar] [CrossRef]
- Gorman, K.M.; Barr, E.L.; Nocera, T.; Ford, W.M. Characteristics of day-roosts used by northern long-eared bats (Myotis septentrionalis) in Coastal New York. Northeast. Nat. 2022, 29, 153–170. [Google Scholar] [CrossRef]
- Bergeson, S.M.; Confortin, K.A.; Carter, T.C.; Karsk, J.R.; Haulton, S.; Burnett, H. Northern long-eared bats roosting in a managed forest in south-central Indiana. For. Ecol. Manag. 2021, 483, 118928. [Google Scholar] [CrossRef]
- Silvis, A.; Ford, W.M.; Britzke, E.R. Day-roost tree selection by northern long-eared bats—What do non-roost tree comparisons and one year of data really tell us? Glob. Ecol. Conserv. 2015, 3, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Solick, D.; Barclay, R. Thermoregulation and roosting behaviour of reproductive and nonreproductive female western long-eared bats (Myotis evotis) in the Rocky Mountains of Alberta. Can. J. Zool. 2006, 84, 589–599. [Google Scholar] [CrossRef]
- Ruch, D.G.; Nelsen, J.; Carlson, R.; Fisher, B.; Fleming, A.H.; Gorney, D.; Holland, J.D.; Jordan, M.; Kingsbury, B.; Murphy, B.; et al. Results of the 2014 Eagle Marsh biodiversity survey, Allen County, Indiana. In Proceedings of the Indiana Academy of Science; Ruch, D.G., Rothrock, P., Eds.; IAS Publications: Indianapolis, IN, USA, 2016; Volume 125, pp. 40–49. [Google Scholar]
- Cli-MATE Daily Data between Two Dates. Available online: https://mrcc.purdue.edu/CLIMATE/Station/Daily/StnDyBTD.jsp (accessed on 18 December 2021).
- Kunz, T.H.; Fenton, M.B. Bat Ecology; The University of Chicago Press: Chicago, IL, USA, 2003. [Google Scholar]
- Kunz, T.H.; Anthony, E.L.P. Age estimation and post-natal growth in the bat Myotis lucifugus. J. Mammal. 1982, 63, 23–32. [Google Scholar] [CrossRef]
- Aldridge, H.D.J.N.; Brigham, R.M. Load carrying and maneuverability in an insectivorous bat: A test of the 5% “rule” of radio-telemetry. J. Mammal. 1988, 69, 379–382. [Google Scholar] [CrossRef]
- Sikes, R.S.; Bryan, J.A., II; Byman, D.; Danielson, B.J.; Eggleston, J.; Gannon, M.R.; Gannon, W.L.; Hale, D.W.; Jesmer, B.R.; Odell, D.K.; et al. 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef] [PubMed]
- Kunz, T.H.; Anthony, E.L.P. Variation in nightly emergence behavior in the little brown bat, Myotis lucifugus (Chiroptera: Vespertilionidae). In Contributions in Mammalogy: A Memorial Volume Honoring J. Knox Jones, Jr.; Genoways, H.H., Baker, R.J., Eds.; Texas Tech University Press: Lubbock, TX, USA, 1996; pp. 225–236. [Google Scholar]
- O’Keefe, J.M.; Loeb, S.C. Indiana bats roost in ephemeral, fire-dependent pine snags in the southern Appalachian Mountains, USA. For. Ecol. Manag. 2017, 391, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Bergeson, S.M.; O’Keefe, J.M.; Haulton, G.S. Managed forests provide roosting opportunities for Indiana bats in south-central Indiana. For. Ecol. Manag. 2018, 427, 305–316. [Google Scholar] [CrossRef]
- Duguay, S.M.; Arii, K.; Hooper, M.; Lechowicz, M.J. Ice storm damage and early recovery in an old-growth forest. Environ. Monit. Assess. 2001, 67, 97–108. [Google Scholar] [CrossRef]
- Barclay, R.M.R.; Kalcounis, M.C.; Crampton, L.H.; Stefan, C.I.; Vonhof, M.J.; Wilkinson, L.; Brigham, R.M. Can external radiotransmitters be used to assess body temperature and torpor in bats? J. Mammal. 1996, 77, 1102–1106. [Google Scholar] [CrossRef] [Green Version]
- Willis, C.K. An energy-based body temperature threshold between torpor and normothermia for small mammals. Physiol. Biochem. Zool. 2007, 80, 643–651. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 9 June 2022).
- Johnson, J.B.; Edwards, J.W.; Ford, W.M.; Gates, J.E. Roost tree selection by northern myotis (Myotis septentrionalis) maternity colonies following prescribed fire in a Central Appalachian Mountains hardwood forest. For. Ecol. Manag. 2009, 258, 233–242. [Google Scholar] [CrossRef]
- Audet, D.; Thomas, D.W. Evaluation of the accuracy of body temperature measurement using external radio transmitters. Can. J. Zool. 1996, 74, 1778–1781. [Google Scholar] [CrossRef]
- Foster, R.W.; Kurta, A. Roosting ecology of the northern bat (Myotis septentrionalis) and comparisons with the endangered Indiana bat (Myotis sodalis). J. Mammal. 1999, 80, 659–672. [Google Scholar] [CrossRef]
- Krynak, T.J. Bat Habitat Use and Roost Tree Selection for Northern Long-Eared Myotis (Myotis septentrionalis) in North-Central Ohio. M.S. Thesis, John Carroll University, University Heights, OH, USA, 2010. [Google Scholar]
- Ford, W.M.; Owen, S.F.; Edwards, J.W.; Jane, L.R. Robinia pseudoacacia (Black Locust) as day-roosts of male Myotis septentrionalis (Northern Bats) on the Fernow Experimental Forest, West Virginia. Northeast. Nat. 2006, 13, 15–24. [Google Scholar] [CrossRef]
- Perry, R.W.; Thill, R.E. Roost selection by male and female northern long-eared bats in a pine-dominated landscape. For. Ecol. Manag. 2007, 247, 220–226. [Google Scholar] [CrossRef]
- Henderson, L.E.; Broders, H.G. Movements and resource selection of the northern long-eared myotis (Myotis septentrionalis) in a forest—Agriculture landscape. J. Mammal. 2008, 89, 952–963. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.E. Roost fidelity of bats: A review. J. Mammal. 1995, 76, 481–496. [Google Scholar] [CrossRef]
- Fenton, M.B.; Rautenbach, I.L.; Smith, S.E.; Swanepoel, C.M.; Grosell, J.; van Jaarsveld, J. Raptors and bats: Threats and opportunities. Anim. Behav. 1994, 48, 9–18. [Google Scholar] [CrossRef]
- Kerth, G.; Konig, B. Fission, fusion and nonrandom associations in female Bechstein’s bats (Myotis bechsteinii). Behaviour 1999, 136, 1187–1202. [Google Scholar] [CrossRef]
- Ruczyński, I.; Bartoń, K.A. Seasonal changes and the influence of tree species and ambient temperature on the fission-fusion dynamics of tree-roosting bats. Behav. Ecol. Sociobiol. 2020, 74, 63. [Google Scholar] [CrossRef]
- Broders, H.; Forbes, G.; Woodley, S.; Thompson, I. Range extent and stand selection for roosting and foraging in forest-dwelling northern long-eared bats and little brown bats in the Greater Fundy Ecosystem, New Brunswick. J. Wildl. Manag. 2006, 70, 1174–1184. [Google Scholar] [CrossRef]
- Gorman, K.M.; Deeley, S.M.; Barr, E.L.; Freeze, S.R.; Kalen, N.; Muthersbaugh, M.S.; Ford, W.M. Broad-scale geographic and temporal assessment of northern long-eared bat (Myotis septentrionalis) maternity colony-landscape association. Endanger. Species Res. 2022, 47, 119–130. [Google Scholar] [CrossRef]
- Racey, P.A.; Swift, S.M. Variations in gestation length in a colony of pipistrelle bats (Pipistrellus pipistrellus) from year to year. Reproduction 1981, 61, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, I.; Barclay, R. Patterns of daily torpor and day-roost selection by male and female big brown bats (Eptesicus fuscus). Can. J. Zool.-Rev. Can. De Zool. 1994, 72, 744–749. [Google Scholar] [CrossRef]
- Menzel, M.A.; Owen, S.F.; Ford, W.M.; Edwards, J.W.; Wood, P.B.; Chapman, B.R.; Miller, K.V. Roost tree selection by northern long-eared bat (Myotis septentrionalis) maternity colonies in an industrial forest of the central Appalachian mountains. For. Ecol. Manag. 2002, 155, 107–114. [Google Scholar] [CrossRef]
- Lacki, M.J.; Schwierjohann, J.H. Day-roost characteristics of northern bats in mixed mesophytic forest. J. Wildl. Manag. 2001, 65, 482. [Google Scholar] [CrossRef]
- Callahan, E.V.; Drobney, R.D.; Clawson, R.L. Selection of summer roosting sites by Indiana bats (Myotis sodalis) in Missouri. J. Mammal. 1997, 78, 818–825. [Google Scholar] [CrossRef]
- Gardner, J.E.; Garner, J.D.; Hofmann, J.E. Summer Roost Selection and Roosting Behavior of Myotis Sodalis (Indiana bat) in Illinois; Illinois Natural History Survey Technical Reports; University of Illinois Press: Champaign, IL, USA, 1991; p. 56. [Google Scholar]
- Lacki, M.J.; Cox, D.R.; Dickinson, M.B. Meta-analysis of summer roosting characteristics of two species of Myotis bats. Am. Midl. Nat. 2009, 162, 318–326. [Google Scholar] [CrossRef]
- Timpone, J.C.; Boyles, J.G.; Murray, K.L.; Aubrey, D.P.; Robbins, L.W. Overlap in Roosting Habits of Indiana Bats (Myotis sodalis) and Northern Bats (Myotis septentrionalis). Am. Midl. Nat. 2010, 163, 115–123. [Google Scholar] [CrossRef]
- Salinas-Ramos, V.B.; Ancillotto, L.; Bosso, L.; Sánchez-Cordero, V.; Russo, D. Interspecific competition in bats: State of knowledge and research challenges. Mammal Rev. 2020, 50, 68–81. [Google Scholar] [CrossRef]
- Starbuck, C.A.; Amelon, S.K.; Thompson, F.R. Relationships between bat occupancy and habitat and landscape structure along a savanna, woodland, forest gradient in the Missouri Ozarks. Wildl. Soc. Bull. 2014, 39, 20–30. [Google Scholar] [CrossRef]
- Haddock, J.K.; Threlfall, C.G.; Law, B.; Hochuli, D.F. Light pollution at the urban forest edge negatively impacts insectivorous bats. Biol. Conserv. 2019, 236, 17–28. [Google Scholar] [CrossRef]
- Moretto, L.; Francis, C.M. What factors limit bat abundance and diversity in temperate, North American urban environments? J. Urban Ecol. 2017, 3, jux016. [Google Scholar] [CrossRef] [Green Version]
- Oberle, B.; Ogle, K.; Zanne, A.E.; Woodall, C.W. When a tree falls: Controls on wood decay predict standing dead tree fall and new risks in changing forests. PLoS ONE 2018, 13, e0196712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, M.; Leak, W.B. Snag longevity in managed northern hardwoods. North. J. Appl. For. 2006, 23, 215–217. [Google Scholar] [CrossRef]
Figure 1.
Map of Fox Island County Park and Eagle Marsh Nature Preserve in Fort Wayne, Indiana, USA (star). Adult female and juvenile northern long-eared bats (Myotis septentrionalis) were captured at three out of eight total net sites and tracked back to their day roosts between May and August 2019–2021. To study roost selection in this colony, characteristics of roosts were compared to characteristics of available trees, located randomly within 0.9 ± 0.4 km buffers (average distance between roosts and capture sites) around each roost.
Figure 1.
Map of Fox Island County Park and Eagle Marsh Nature Preserve in Fort Wayne, Indiana, USA (star). Adult female and juvenile northern long-eared bats (Myotis septentrionalis) were captured at three out of eight total net sites and tracked back to their day roosts between May and August 2019–2021. To study roost selection in this colony, characteristics of roosts were compared to characteristics of available trees, located randomly within 0.9 ± 0.4 km buffers (average distance between roosts and capture sites) around each roost.

Figure 2.
Tree species used by a population of northern long-eared bats (Myotis septentrionalis) as roosts (black) and available trees within foraging range (white) of capture locations in Fox Island County Park in Fort Wayne, IN. Data were collected between May and August 2019–2021. Bats preferentially selected for silver maples (Acer saccharinum; p < 0.01).
Figure 2.
Tree species used by a population of northern long-eared bats (Myotis septentrionalis) as roosts (black) and available trees within foraging range (white) of capture locations in Fox Island County Park in Fort Wayne, IN. Data were collected between May and August 2019–2021. Bats preferentially selected for silver maples (Acer saccharinum; p < 0.01).

Figure 3.
Measurements of ambient temperature (white) and roost tree temperature (black) for roosts containing temperature-sensitive transmitters dropped by tagged northern long-eared bats (Myotis septentrionalis). Temperature averages are reported for measurements taken during the day (6:54–21:54), at night (21:54–6:54), and overall. Data were collected in Fox Island County Park in Fort Wayne, IN, between May and August 2019–2021.
Figure 3.
Measurements of ambient temperature (white) and roost tree temperature (black) for roosts containing temperature-sensitive transmitters dropped by tagged northern long-eared bats (Myotis septentrionalis). Temperature averages are reported for measurements taken during the day (6:54–21:54), at night (21:54–6:54), and overall. Data were collected in Fox Island County Park in Fort Wayne, IN, between May and August 2019–2021.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of roost and available tree characteristics for a population of northern long-eared bats (Myotis septentrionalis) in Fort Wayne, Indiana, studied from 2019–2021. Bold text designates statistical significance. Data are reported as mean ± SE.
Table 1.
Comparison of roost and available tree characteristics for a population of northern long-eared bats (Myotis septentrionalis) in Fort Wayne, Indiana, studied from 2019–2021. Bold text designates statistical significance. Data are reported as mean ± SE.
| Roost Characteristics | Roost Trees (n = 12) | Available Trees (n = 12) | t- or V-Value | p-Value | Adjusted p-Value |
|---|---|---|---|---|---|
| Tree-Level | |||||
| Tree Height (m) | 20.7 ± 2.6 | 15.9 ± 2.2 | t = −1.53 | 0.15 | 0.28 |
| Tree Diameter at Breast Height (cm) | 51.4 ± 6.8 | 48.7 ± 6.8 | t = −0.24 | 0.81 | 1.00 |
| Bark Remaining (%) | 75.8 ± 7.6 | 61.7 ± 10.4 | t = −1.13 | 0.28 | 0.44 |
| Roost Canopy Closure (%) | 28.2 ± 8.6 | 27.6 ± 7.7 | t = −0.06 | 0.96 | 1.00 |
| Plot-Level | |||||
| Plot Canopy Closure (%) | 20.9 ± 9.0 | 21.9 ± 7.0 | V = 27.5 | 1.00 | 1.00 |
| Number of Snags | 22 ± 3 | 6 ± 2 | t = −4.17 | <0.01 | 0.02 |
| Number of Live Trees | 4 ± 2 | 16 ± 4 | V = 49.0 | 0.03 | 0.09 |
| Landscape-Level | |||||
| Distance to Water (m) | 30.4 ± 18.1 | 113.7 ± 37.3 | V = 42.0 | 0.02 | 0.09 |
| Distance to Forest Edge (m) | 105.3 ± 13.3 | 106.2 ± 31.2 | t = −0.70 | 0.50 | 0.69 |
| Distance to Fox Island Forest (m) | 87.7 ± 17.7 | 430.7 ± 135.2 | V = 55.0 | 0.06 | 0.12 |
| Distance to Snag Forest (m) | 27.3 ± 18.4 | 444.8 ± 118.4 | V = 55.0 | 0.01 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Burrell, G.E.; Bergeson, S.M. Roosting Behavior of Northern Long-Eared Bats (Myotis septentrionalis) in an Urban-Adjacent Forest Fragment. Forests 2022, 13, 1972. https://0-doi-org.brum.beds.ac.uk/10.3390/f13121972
AMA Style
Burrell GE, Bergeson SM. Roosting Behavior of Northern Long-Eared Bats (Myotis septentrionalis) in an Urban-Adjacent Forest Fragment. Forests. 2022; 13(12):1972. https://0-doi-org.brum.beds.ac.uk/10.3390/f13121972
Chicago/Turabian StyleBurrell, Galen E., and Scott M. Bergeson. 2022. "Roosting Behavior of Northern Long-Eared Bats (Myotis septentrionalis) in an Urban-Adjacent Forest Fragment" Forests 13, no. 12: 1972. https://0-doi-org.brum.beds.ac.uk/10.3390/f13121972
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.