
The Edaphic and Vegetational Properties Controlling Soil Aggregate Stability Vary with Plant Communities in an Arid Desert Region of Northwest China


Abstract
:1. Introduction
2. Materials and Methods
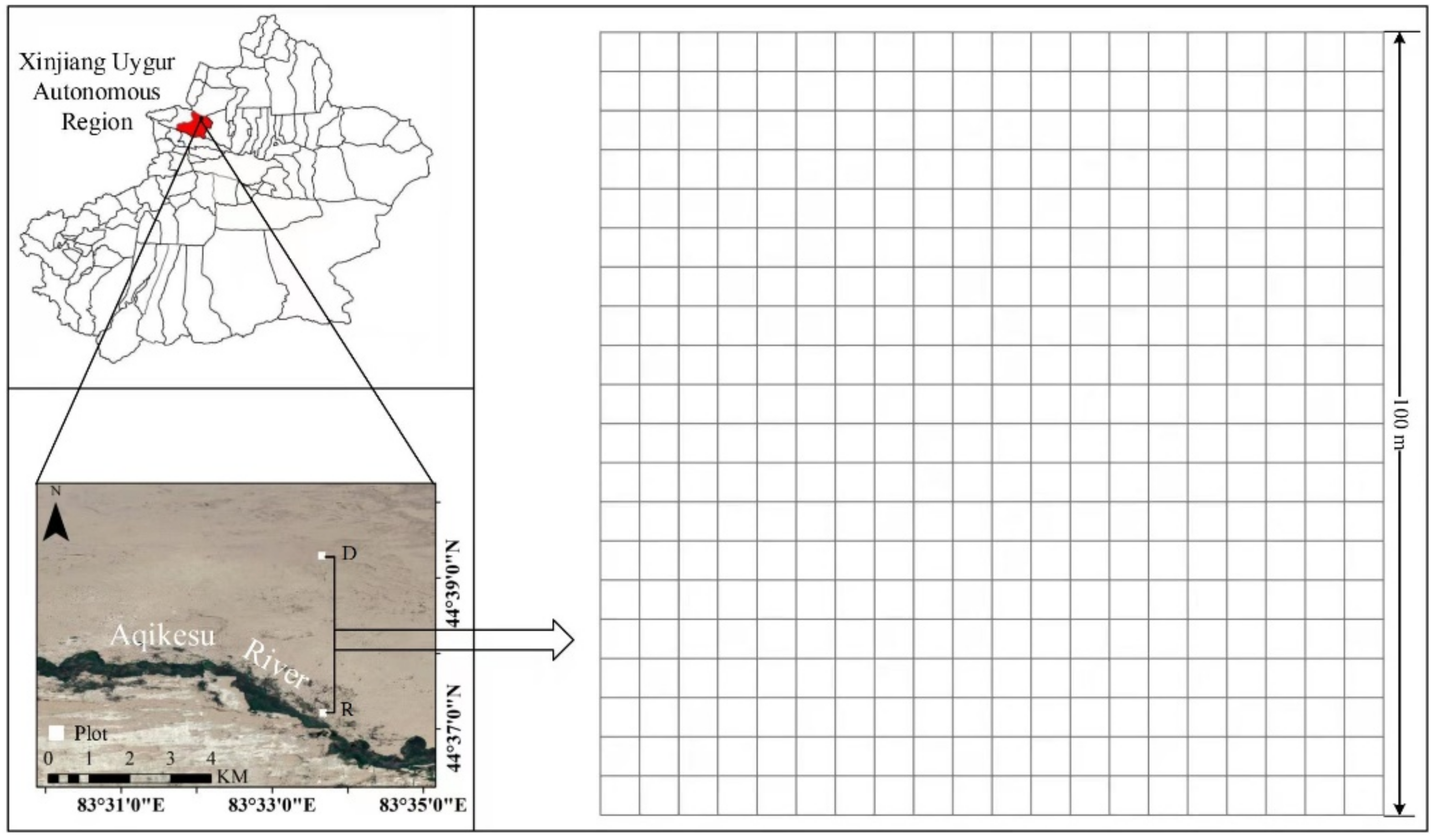
2.1. Study Area, Sample Layout, and Plant Sample Collection
2.2. Collection and Measurement of Soil Samples
2.3. Calculations for Soil Aggregates and Plant Diversity
2.3.1. Calculations for Soil Aggregates
2.3.2. Calculation of Plant Diversity
2.4. Data Analysis
3. Results
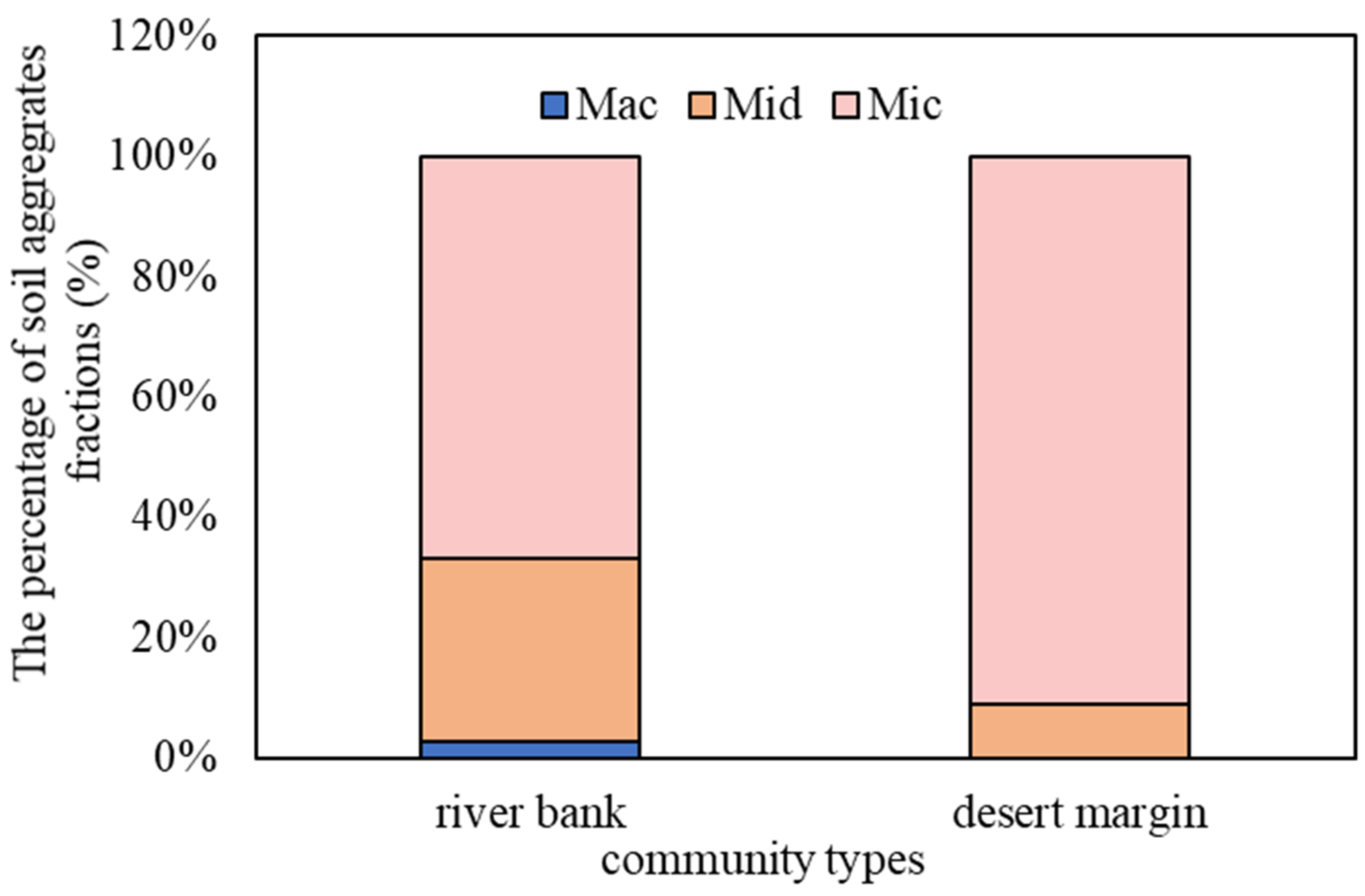
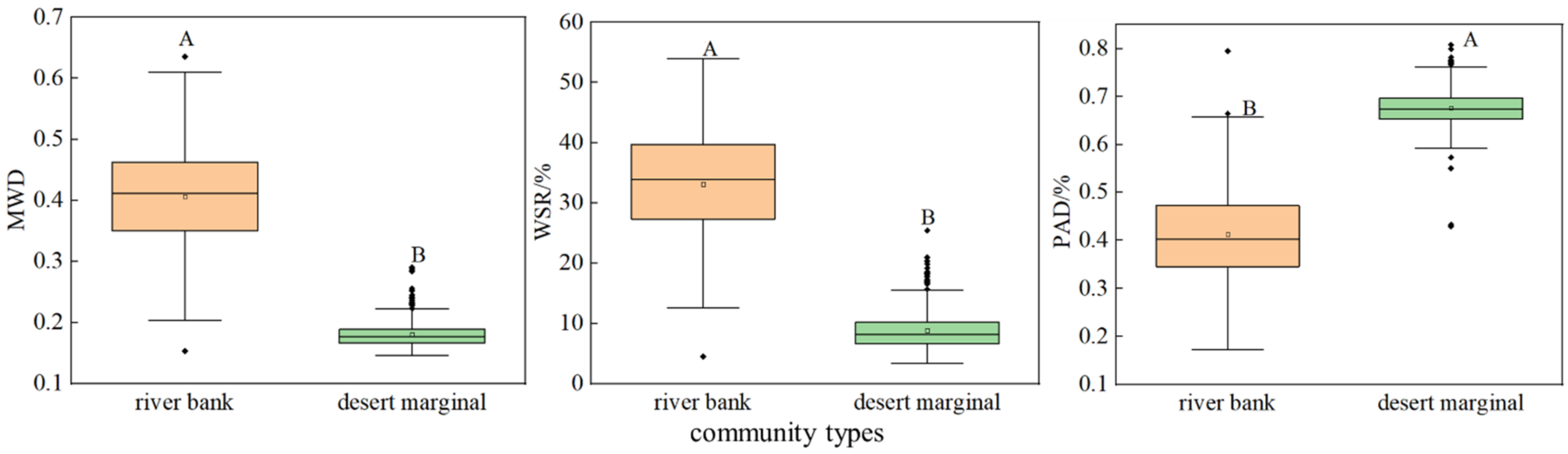
3.1. Differences in the Proportion and Stability of Soil Aggregates in Different Community Types
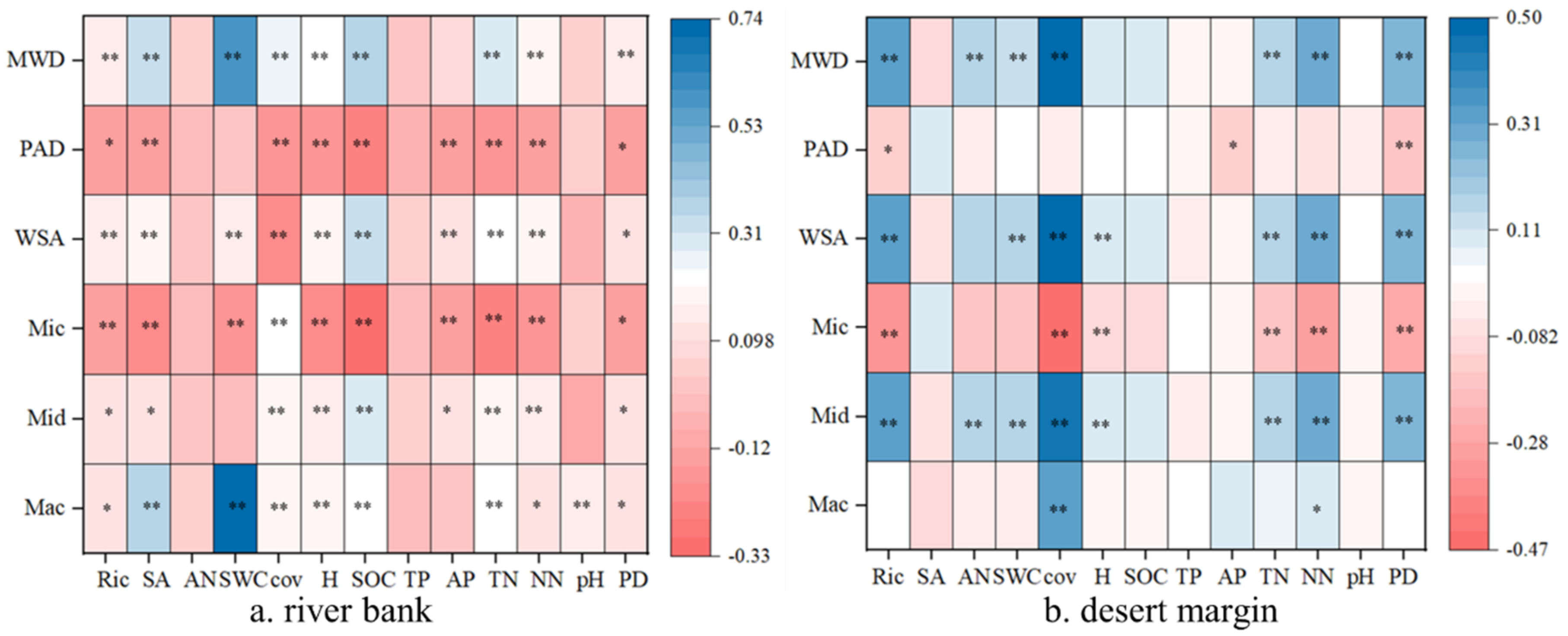
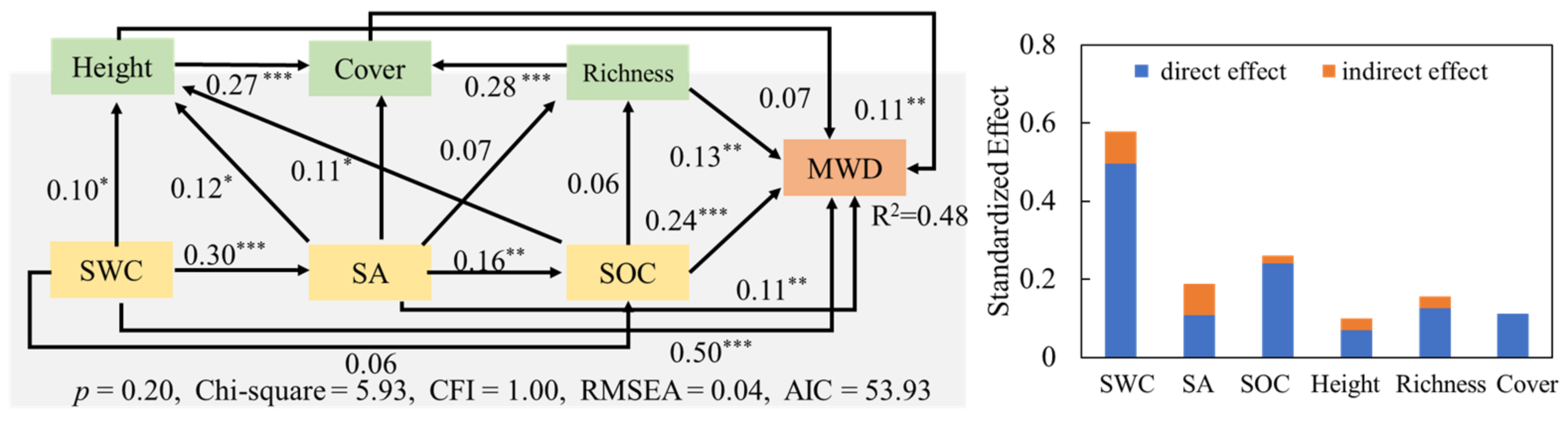
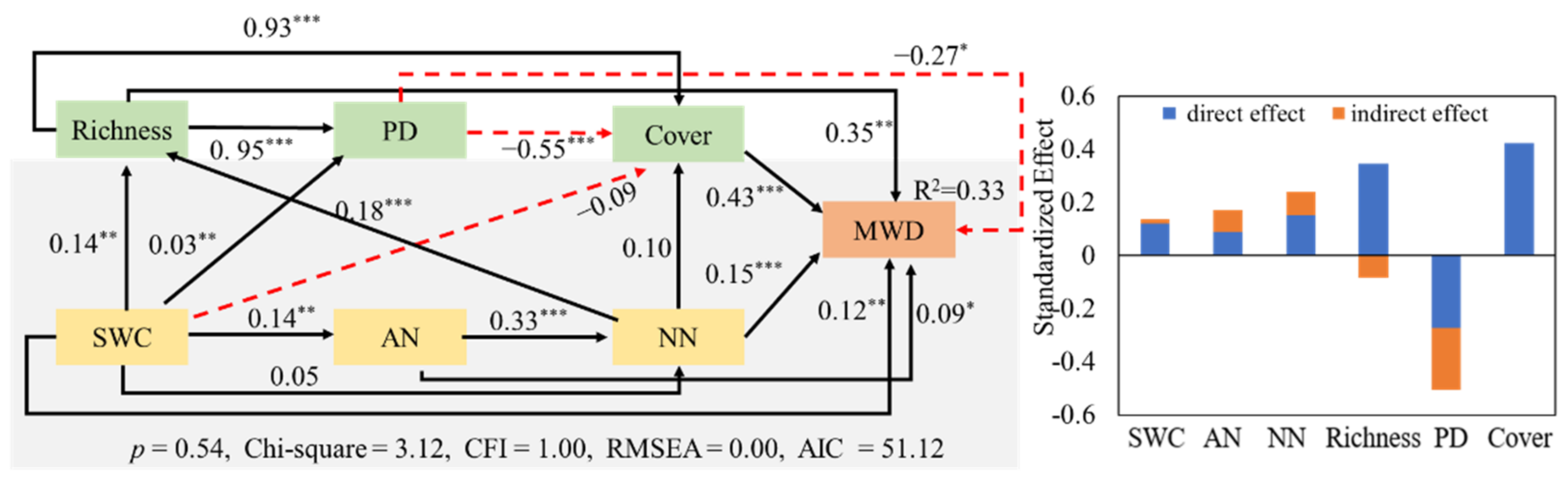
3.2. Influencing Factors of Soil Aggregates
3.3. Effects of Soil Factors and Plant Characteristics on the Stability of Soil Aggregates
4. Discussion
4.1. Differences in the Proportion and Stability of Soil Aggregates in Different Community Types
4.2. Influence of Soil Factors on the Stability of Soil Aggregates
4.3. Effects of Plant Characteristics on the Stability of Soil Aggregates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yılmaz, E.; Çanakcı, M.; Topakcı, M.; Sönmez, S. Effect of vineyard pruning residue application on soil aggregate formation, aggregate stability and carbon content in different aggregate sizes. Catena 2019, 183, 104219. [Google Scholar] [CrossRef]
- Fialho, R.C.; Zinn, Y.L. Changes in soil organic carbon under eucalyptus plantations in Brazil: A comparative analysis. Land Degrad. Dev. 2014, 25, 428–437. [Google Scholar] [CrossRef]
- Parras-Alcántara, L.; Díaz-Jaimes, L.; Lozano-García, B. Management effects on soil organic carbon stock in Mediterranean open rangelands-treeless grasslands. Land Degrad. Dev. 2015, 26, 22–34. [Google Scholar] [CrossRef]
- Elliott, E.T.; Coleman, D.C. Let the soil work for us. Ecol. Bull. 1988, 39, 23–32. [Google Scholar]
- Feeney, D.S.; Crawford, J.W.; Daniell, T.; Hallett, P.D.; Nunan, N.; Ritz, K.; Rivers, M.; Young, I.M. Three-dimensional microorganization of the soil-root-microbe system. Microbic. Ecol. 2006, 52, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Young, I.M.; Crawford, J.W.; Rappoldt, C. New methods and models for characterizing structural heterogeneity of soil. Soil Tillage Res. 2001, 61, 33–45. [Google Scholar] [CrossRef]
- Bottinelli, N.; Jouquet, P.; Capowiez, Y.; Podwojewski, P.; Grimaldi, M.; Peng, X. Why is the influence of soil macrofauna on soil structure only considered by soil ecologists? Soil Tillage Res. 2015, 146, 118–124. [Google Scholar] [CrossRef]
- An, S.; Mentler, A.; Mayer, H.; Blum, W.E.H. Soil aggregation, aggregate stability, organic carbon and nitrogen in different soil aggregate fractions under forest and shrub vegetation on the Loess Plateau, China. Catena 2010, 81, 226–233. [Google Scholar] [CrossRef]
- Deviren, S.S.; Cornelis, W.M.; Erpul, G.; Gabriels, D. Comparison of different aggregate stability approaches for loamy sand soils. Appl. Soil Ecol. 2012, 54, 1–6. [Google Scholar] [CrossRef]
- Blankinship, J.C.; Fonte, S.J.; Six, J.; Schimela, J.D. Plant versus microbial controls on soil aggregate stability in a seasonally dry ecosystem. Geoderma 2016, 272, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Bedel, L.; Legout, A.; Poszwa, A.; Heijden, G.V.D.; Court, M.; Goutal-Pousse, N.; Montarges-Pelletier, E.; Ranger, J. Soil aggregation may be a relevant indicator of nutrient cation availability. Ann. For. Sci. 2018, 75, 103. [Google Scholar] [CrossRef] [Green Version]
- Egan, G.; Crawley, M.J.; Fornara, D.A. Effects of long-term grassland management on the carbon and nitrogen pools of different soil aggregate fractions. Sci. Total Environ. 2018, 613, 810–819. [Google Scholar] [CrossRef]
- Six, J. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Dimoyiannis, D. Wet aggregate stability as affected by excess carbonate and other soil properties. Land Degrad. Dev. 2012, 23, 450–455. [Google Scholar] [CrossRef]
- Regelink, I.C.; Stoof, C.R.; Rousseva, S.; Weng, L.P.; Lairef, G.J.; Kramg, P.; Nikolaos, P.; Nikolaidish, N.P.; Kerchevaf, M.; Banwartgi, S.; et al. Linkages between aggregate formation, porosity and soil chemical properties. Geoderma 2015, 247, 24–37. [Google Scholar] [CrossRef]
- Duchicela, J.; Sullivan, T.; Bontti, E.; Bever, J.D. Soil aggregate stability increase is strongly related to fungal community succession along an abandoned agricultural field chronosequence in the Bolivian Altiplano. J. Appl. Ecol. 2013, 50, 1266–1273. [Google Scholar] [CrossRef]
- Paul, B.K.; Vanlauwe, B.; Ayuke, F.; Gassner, A.; Hoogmoed, M.; Hurisso, T.T.; Koala, S.; Lelei, D.; Ndabamenye, T.; Six, J.; et al. Medium-term impact of tillage and residue management on soil aggregate stability, soil carbon and crop productivity. Agric. Ecosyst. Environ. 2013, 164, 14–22. [Google Scholar] [CrossRef]
- Onweremadu, E.U.; Onyia, V.N.; Anikwe, M. Carbon and nitrogen distribution in water-stable aggregates under two tillage techniques in Fluvisols of Owerri area, southeastern Nigeria. Soil Tillage Res. 2007, 97, 195–206. [Google Scholar] [CrossRef]
- Reich, P.B.; Tilman, D.; Isbell, F.; Mueller, K.; Hobbie, S.E.; Flynn, D.F.B.; Eisenhauer, N. Impacts of biodiversity loss escalate through time as redundancy fades. Science 2012, 336, 589–592. [Google Scholar] [CrossRef] [Green Version]
- Peres, G.; Cluzeau, D.; Menasseri, S.; Soussana, J.F.; Bessler, H.; Engels, C.; Habekost, M.; Gleixner, G.; Weigelt, A.; Weisser, W.W.; et al. Mechanisms linking plant community properties to soil aggregate stability in an experimental grassland plant diversity gradient. Plant Soil 2013, 373, 285–299. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Lanoue, A.; Strecker, T.; Scheu, S.; Steinauer, K.; Thakur, M.P.; Mommer, L. Root biomass and exudates link plant diversity with soil bacterial and fungal biomass. Sci. Rep. 2017, 7, 44641. [Google Scholar] [CrossRef] [Green Version]
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Sher, Y.; Baker, N.R.; Herman, D.; Fossum, C.; Hale, L.; Zhang, X.X.; Nuccio, E.; Saha, M.; Zhou, J.H.; Pett-Ridge, J. Microbial extracellular polysaccharide production and aggregate stability controlled by switchgrass (Panicum virgatum) root biomass and soil water potential. Soil Biol. Biochem. 2020, 143, 107742. [Google Scholar] [CrossRef]
- Zhao, D.; Xu, M.; Liu, G.; Ma, L.; Zhang, S.; Xiao, T.; Peng, G. Effect of vegetation type on microstructure of soil aggregates on the Loess Plateau, China. Agric. Ecosyst. Environ. 2017, 242, 1–8. [Google Scholar] [CrossRef]
- Zeng, Q.; Darboux, F.; Man, C.; Zhu, Z.; An, S. Soil aggregate stability under different rain conditions for three vegetation types on the Loess Plateau, China. Catena 2018, 167, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Bronick, C.J.; Lal, R. Soil Structure and Management: A Review. Geoderma. 2005, 124, 3–22. [Google Scholar] [CrossRef]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef] [PubMed]
- Sarker, T.C.; Incerti, G.; Spaccini, R.O.; Piccolo, A.; Mazzoleni, S.; Bonanomi, G. Linking organic matter chemistry with soil aggregate stability: Insight from 13 C NMR spectroscopy. Soil Biol. Biochem. 2018, 117, 175–184. [Google Scholar] [CrossRef]
- Demenois, J.; Carriconde, F.; Rey, F.; Stokes, A. Tropical plant communities modify soil aggregate stability along a successional vegetation gradient on a Ferralsol. Ecol. Eng. 2017, 109, 161–168. [Google Scholar] [CrossRef]
- Dou, Y.; Yang, Y.; An, S.; Zhu, Z. Effects of different vegetation restoration measures on soil aggregate stability and erodibility on the Loess Plateau, China. Catena 2019, 185, 104294. [Google Scholar] [CrossRef]
- Whitford, W.G. Ecology Desert Systems; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Yang, H.; Wang, J.; Zhang, F. Soil aggregation and aggregate-associated carbon under four typical halophyte communities in an arid area. Environ. Sci. Pollut. Res. 2016, 23, 1–10. [Google Scholar] [CrossRef]
- Abdi, E.; Saleh, H.R.; Majnonian, B.; Deljouei, A. Soil fixation and erosion control by Haloxylon persicum roots in arid lands, Iran. J. Arid. Land 2019, 11, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Lv, G.H.; Guo, Z.J.; Chen, Y. Influence of aridity and salinity on plant nutrient s scales up from species to community level in a desert ecosystem. Sci. Rep. 2017, 7, 6811. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.D.; Zhang, X.N.; Lv, G.H.; Arshad, A. Linking Populus euphratica hydraulic redistribution to diversity assembly in the arid desert zone of Xin jiang, China. PLoS ONE 2014, 9, e109071. [Google Scholar]
- Zhang, Z.S.; Dong, X.J.; Xu, B.X.; Dong, X.J.; Zhang, Z.S.; Gao, Y.H.; Hu, Y.G.; Huang, L. Soil respiration sensitivities to water and temperature in a revegetated desert. J. Geophys. Res. Biogeosci. 2015, 120, 773–787. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Norman, R.; Edberg, J.; Stucki, J. Determination of Nitrate in Soil Extracts by Dual-wavelength Ultraviolet Spectrophotometry. Soil Sci. Soc. Am. J. 1985, 49, 1182–1185. [Google Scholar] [CrossRef]
- Dalai, R.C.; Sahrawat, K.L.; Myers, R.J.K. Inclusion of nitrate and nitrite in the Kjeldahl nitrogen determination of soils and plant materials using sodium thiosulphate. Commun. Soil Sci. Plant Anal. 1984, 15, 1453–1461. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Song, X.; Chang, S.X.; Peng, C.G.; Xiao, W.F.; Zhang, G.B.; Xiang, W.H.; Li, Y.; Wang, W.F. Nitrogen depositions increase soil respiration and decrease temperature sensitivity in a Moso bamboo forest. Agric. For. Meteorol. 2019, 268, 48–54. [Google Scholar] [CrossRef]
- Zhao, Q.G. Nanjing Institute of Soil, Chinese Academy of Sciences Soil Physical and Chemical Analysis; Shanghai Science and Technology Press: Shanghai, China, 1983; pp. 62–126. [Google Scholar]
- Zhao, J.; Chen, S.; Hu, R.; Li, Y. Aggregate stability and size distribution of red soils under different land uses integrally regulated by soil organic matter, and iron and aluminum oxides. Soil Tillage Res. 2017, 167, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Mengke, Z.; Siqian, Y.; Shenghao, A.; Xiaoyan, A.; Xue, J.; Jiao, C.; Ruirui, L.; Yingwei, A. Artificial soil nutrient, aggregate stability and soil quality index of restored cut slopes along altitude gradient in southwest China. Chemosphere 2020, 246, 125687. [Google Scholar] [CrossRef]
- Mulumba, L.N.; Lal, R. Mulching effects on selected soil physical properties. Soil Tillage Res. 2008, 98, 106–111. [Google Scholar] [CrossRef]
- Zhao, Y.P.; Meng, M.J.; Zhang, J.C.; Ma, J.Y.; Liu, S.L. Study on the composition and stability of soil aggregates of the main forest stands in Fengyang Mountain, Zhejiang Province. J. Nanjing For. Univ. 2018, 42, 84–90. [Google Scholar]
- Wang, X.Y.; Zhou, C.; Feng, W.H.; Zhang, Y.; Cheng, J.L.; Jiang, X.H. Changes of soil aggregates and organic carbon in Chinese Fir Plantation with different forest ages. J. Soil Water Conserv. 2019, 33, 126–131. [Google Scholar]
- Chen, H.; Ma, W.M.; Zhou, Q.P.; Yang, Y.; Liu, C.W.; Liu, J.Q.; Du, Z.M. Shrub encroachement effects on the stability of soil aggregates and the differeneiation of Fe and Al oxides in Qinghai-Tibet alpine grassland. Acta Pratacult. Sin. 2020, 29, 73–84. [Google Scholar]
- Dai, W.H.; Huang, Y.; Wu, L.; Yu, J. Relationship between soil organic matter content and pH in topsoil of zone soils in China. Acta Pedol. Sin. 2009, 46, 851–860. [Google Scholar]
- Wang, X.X.; Dong, S.K.; Gao, Q.Z.; Zhang, Y.; Hu, G.Z.; Luo, W.R. The rate of soil nitrogen transformation decreased by the degradation of alpine grassland in Qinghai Tibet Plateau. Acta Pratacult. Sin. 2018, 27, 1–9. [Google Scholar]
- Liao, J.J.; Huang, B.; Sun, W.X.; Zou, Z.; Su, J.P.; Ding, F.; Huang, Y. Spatio-Temporal variation of soil available phosphorus and its influencing factors—Acasestudy of rugao county, Jiangsu province. Acta Pedol. Sin. 2007, 4, 620–628. [Google Scholar]
- Pohl, M.; Stroude, R.; Buttler, A.; Rixen, C. Functional traits and root morphology of alpine plants. Ann. Bot. 2011, 108, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Leffelaar, P.A. Water movement, oxygen supply and biological processes on the aggregate scale. Geoderma 1993, 57, 143–165. [Google Scholar] [CrossRef] [Green Version]
- He, X.L.; Gao, L.; Zhao, L. Effects of AM fungi on the growth and drought resistance of Seriphidium minchünense under water stress. Acta Ecol. Sin. 2011, 31, 1029–1037. [Google Scholar]
- Pulleman, M.M.; Marinissen, J. Physical protection of mineralizable C in aggregates from long-term pasture and arable soil. Geoderma 2004, 120, 273–282. [Google Scholar] [CrossRef]
- Liu, W.T.; Wang, T.L.; Zhang, S.; Ding, L.J.; Lv, S.J.; Wei, Z.J. Effects of grazing on edificators and soil aggregate characteristics in Stipa breviflora desert steppe. Ecol. Environ. Sci. 2017, 26, 978–984. [Google Scholar]
- Chen, F.S.; Yu, K.; Gan, L.; Liu, Y.; Hu, X.F.; Ge, G. Effects of temperature moisture and forests succession on nitrogen mineralization in hillside red soils in mid-subtropical region China. Chin. J. Appl. Ecol. 2009, 20, 1529–1535. [Google Scholar]
- Totsche, K.U.; Amelung, W.; Gerzabek, M.H.; Guggenberger, G.; Klumpp, E.; Knief, C.; Lehndorff, E.; Mikutta, R.; Peth, S.; Prechtel, A. Microaggregates in soils. J. Plant Nutr. Soil Sci. 2018, 181, 104–136. [Google Scholar] [CrossRef] [Green Version]
- Chambers, L.G.; Osborne, T.Z.; Reddy, K.R. Effect of salinity-altering pulsing events on soil organic carbon loss along an intertidal wetland gradient: A laboratory experiment. Biogeochemistry 2013, 115, 363–383. [Google Scholar] [CrossRef]
- Yu, H.Y.; Li, T.X.; Zhou, J.M. Secondary Salinization of Greenhouse Soil and Its Effects on Soil Properties. Soils 2005, 6, 581–586. [Google Scholar]
- Erktan, A.L.; Cécillon, L.; Graf, F.; Roumet, C.; Legout, C.; Rey, F. Increase in soil aggregate stability along a Mediterranean successional gradient in severely eroded gully bed ecosystems: Combined effects of soil, root traits and plant community characteristics. Plant Soil 2016, 398, 121–137. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; Vries, F. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; Deyn, G.B.D.; Bardgett, R.D.; Seabloom, E. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef]
- Liu, R.; Zhou, X.; Wang, J.; Shao, J.; Fu, Y.; Liang, C.; Yan, E.; Chen, X.; Wang, X.; Bai, S.H. Differential magnitude of rhizosphere effects on soil aggregation at three stages of subtropical secondary forest successions. Plant Soil 2019, 436, 365–380. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [Green Version]
- Winter, M.; Devictor, V.; Schweiger, O. Phylogenetic diversity and nature conservation: Where are we? Trends Ecol. Evol. 2013, 28, 199–204. [Google Scholar] [CrossRef]
- Haynes, R.J.; Beare, M.H. Influence of six crop species on aggregate stability and some labile organic matter fractions. Soil Biol. Biochem. 1997, 29, 1647–1653. [Google Scholar] [CrossRef]
- Wright, S.F.; Upadhyaya, A. Extraction of an abundant and unusual protein from soil and comparison with hyphal protein of Arbuscular Mycorrhizal Fungi. Soil Sci. 1996, 161, 575–586. [Google Scholar] [CrossRef]
- Pohl, M.; Alig, D.; Krner, C.; Rixen, C. Higher plant diversity enhances soil stability in disturbed alpine ecosystems. Plant Soil 2009, 324, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.Y.; Zhou, Z.C.; Su, X.M. Review of the mechanism of root systems on the formation of soil aggregate. J. Soil Water Conserv. 2020, 34, 267–273, 298. [Google Scholar]
- Bird, S.B.; Herrick, J.E.; Wander, M.M.; Murray, L. Multi-scale variability in soil aggregate stability: Implications for understanding and predicting semi-arid grassland degradation. Geoderma 2007, 140, 106–118. [Google Scholar] [CrossRef]
- Zuazo, V.H.D.; Pleguezuelo, C.R.R. Soil-erosion and run off prevention by plant covers. A review. Agron. Sustain. Dev. 2008, 28, 65–86. [Google Scholar] [CrossRef] [Green Version]
- Agata, K.; Renée, M.B.; Rudy, V.D.; Kotowski, W. Species trait shifts in vegetation and soil seed bank during fen degradation. Plant Ecol. 2010, 206, 59–82. [Google Scholar]
- Lu, Q.; Ma, H.; Yu, H.; Wang, L.; Shen, Y.; Xu, D.M.; Xie, Y.Z. Effects of rotational grazing methods on soil aggregates and organic carbon characteristics in desert steppe. Chin. J. Appl. Ecol. 2019, 30, 3028–3038. [Google Scholar]
- Zhang, S.J.; He, X.B.; Bao, Y.H.; Tang, Q. Change characteristics of soil aggregates at different water levels in the water-level fluctuation zone of the three Gorges reservoir. Res. Soil Water Conserv. 2021, 28, 25–30. [Google Scholar]
- Du, H.D.; Jiao, J.Y.; Jia, Y.F.; Wang, N.; Wang, D.L. Phytogenic mounds of four typical shoot architecture species at different slope gradients on the Loess Plateau of China. Geomorphology 2013, 193, 57–64. [Google Scholar] [CrossRef]
- Pastrán, G.; Carretero, E.M. Phytogenic Mounds (Nebkhas): Effect of Tricomaria usillo on Sand Entrapment in Central-West of Argentina. J. Geogr. Inf. Syst. 2016, 8, 429–437. [Google Scholar]
- Xiao, H.L.; Zhang, J.X.; Li, J.H. Dustfall Particle size and sedimentation rate at the Souther edge of Tengger Desert. J. Desert Res. 1997, 17, 127–132. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River Bank | Desert Margin | |
|---|---|---|
| Vegetation | ||
| Plant height/cm | 4–1350 | 3.5–420 |
| Species number | 14 | 10 |
| Simpson | 1.52 ± 0.03 a | 1.54 ± 0.03 a |
| Maglef | 0.59 ± 0.02 a | 0.53 ± 0.02 b |
| Shannon–Weiner | 0.52 ± 0.02 a | 0.50 ± 0.02 a |
| Phylogenetic richness | 384.24 ± 0.09 a | 187.82 ± 0.11 b |
| Soil | ||
| pH | 8.07 ± 0.02 a | 7.38 ± 0.02 b |
| SWC % | 13.12 ± 0.19 a | 1.04 ± 0.02 b |
| SA (g/kg) | 5.58 ± 0.12 a | 1 ± 0.02 b |
| SOC (g/kg) | 9.57 ± 0.28 a | 1.35 ± 0.03 b |
| TN (g/kg) | 1.31 ± 0.01 a | 0.59 ± 0.01 b |
| AP (g/kg) | 38.19 ± 0.75 a | 7.96 ± 0.18 b |
| TP (mg/kg) | 2.05 ± 0.04 a | 0.63 ± 0.01 b |
| AN (mg/kg) | 2.47 ± 0.06 a | 1.39 ± 0.03 b |
| NN (mg/kg) | 12.51 ± 0.36 a | 2.76 ± 0.06 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, L.; Hu, D.; Lv, G. The Edaphic and Vegetational Properties Controlling Soil Aggregate Stability Vary with Plant Communities in an Arid Desert Region of Northwest China. Forests 2022, 13, 368. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030368
Jiang L, Hu D, Lv G. The Edaphic and Vegetational Properties Controlling Soil Aggregate Stability Vary with Plant Communities in an Arid Desert Region of Northwest China. Forests. 2022; 13(3):368. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030368
Chicago/Turabian StyleJiang, Lamei, Dong Hu, and Guanghui Lv. 2022. "The Edaphic and Vegetational Properties Controlling Soil Aggregate Stability Vary with Plant Communities in an Arid Desert Region of Northwest China" Forests 13, no. 3: 368. https://0-doi-org.brum.beds.ac.uk/10.3390/f13030368