
Herbivory Rather than Root Competition and Environmental Factors Determines Plant Establishment in Fragmented Forests



Abstract
:1. Introduction
2. Materials and Methods
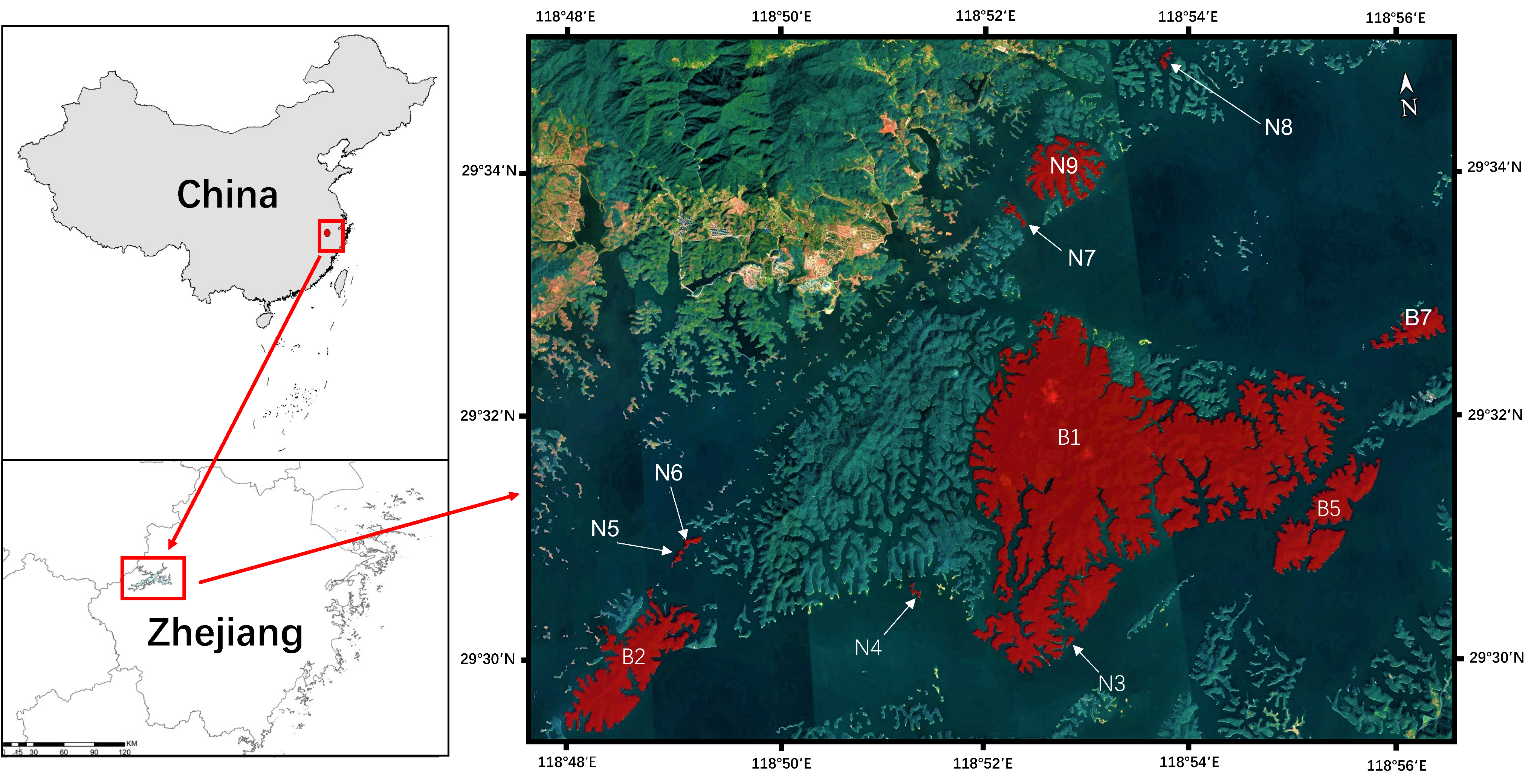
2.1. Study Site
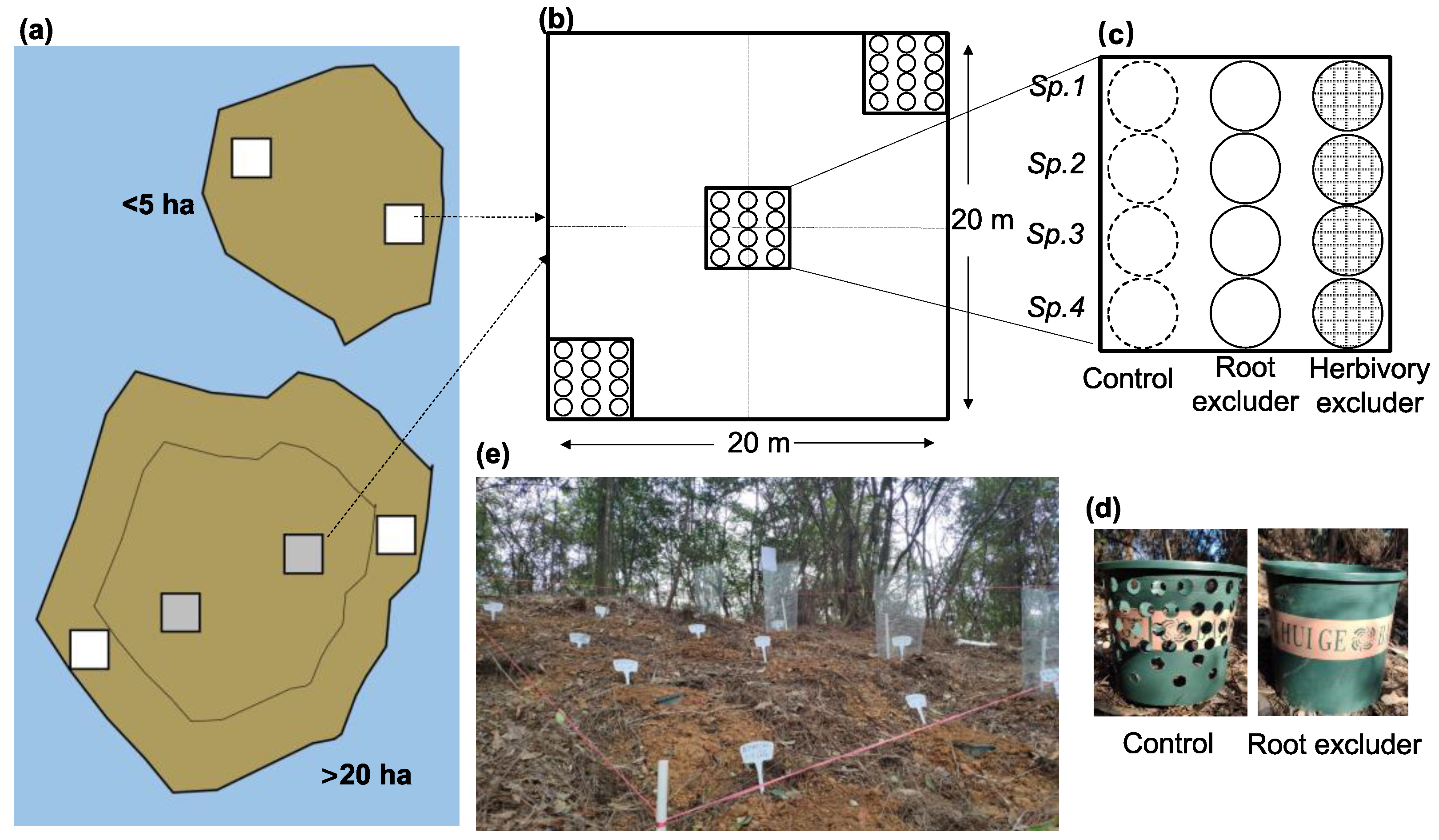
2.2. Experimental Design
2.3. Species Selection and Sampling
2.4. Measurement of Environmental Factors
2.5. Data Analysis
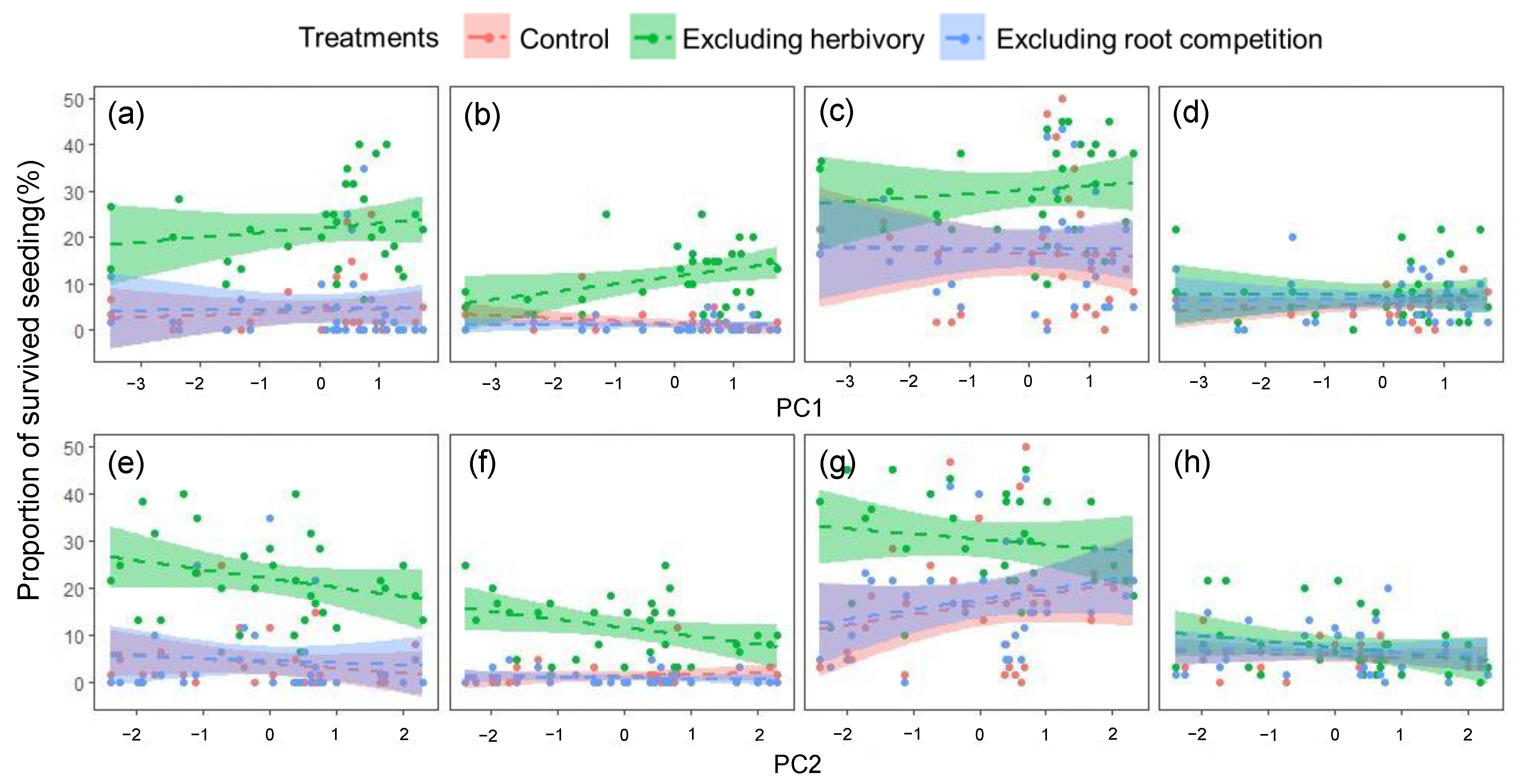
3. Results
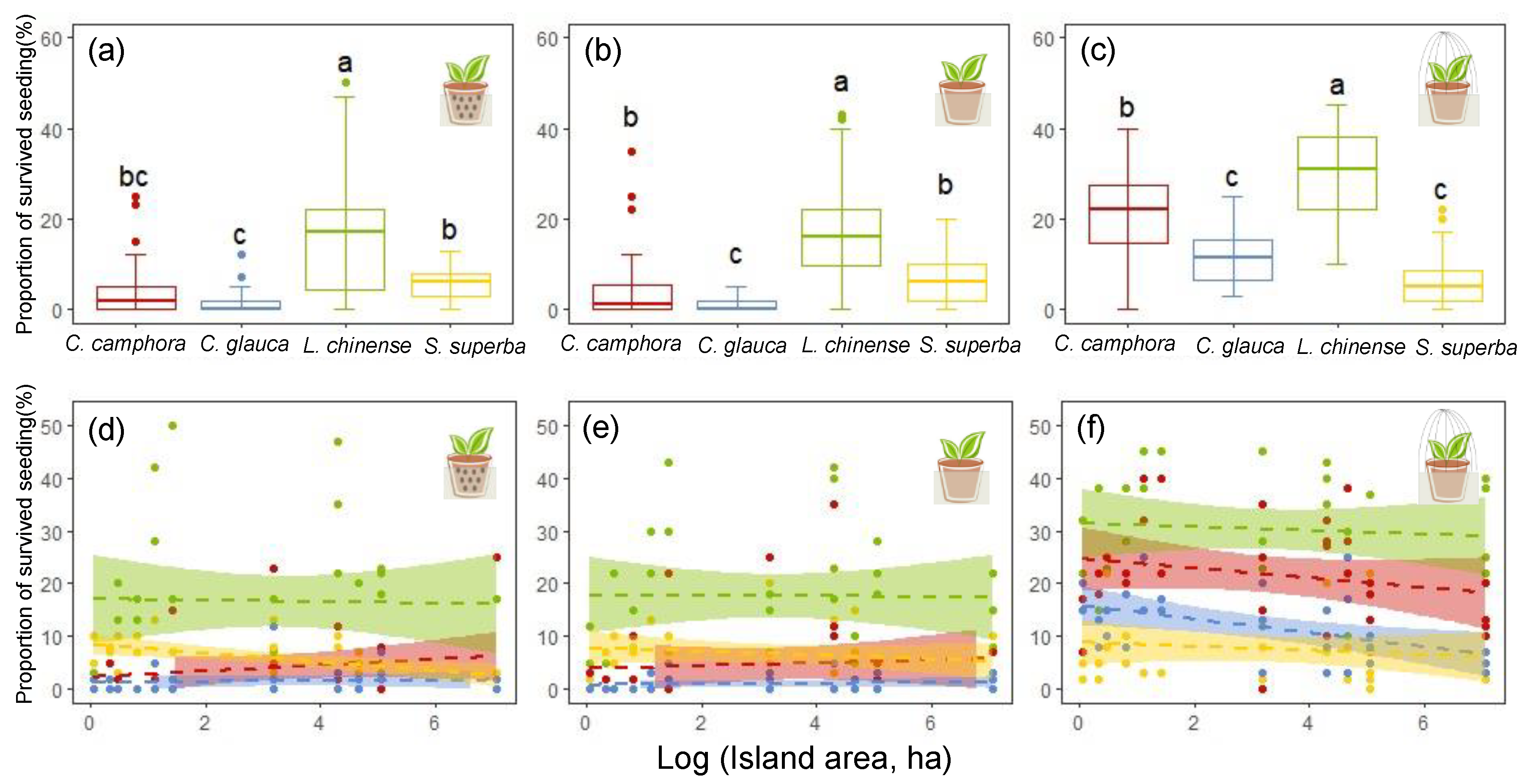
3.1. Differences in Seed Gemmation and Early Seedling Survival among Species
3.2. Differences among Treatments in Seed Gemmation and Early Seedling Survival
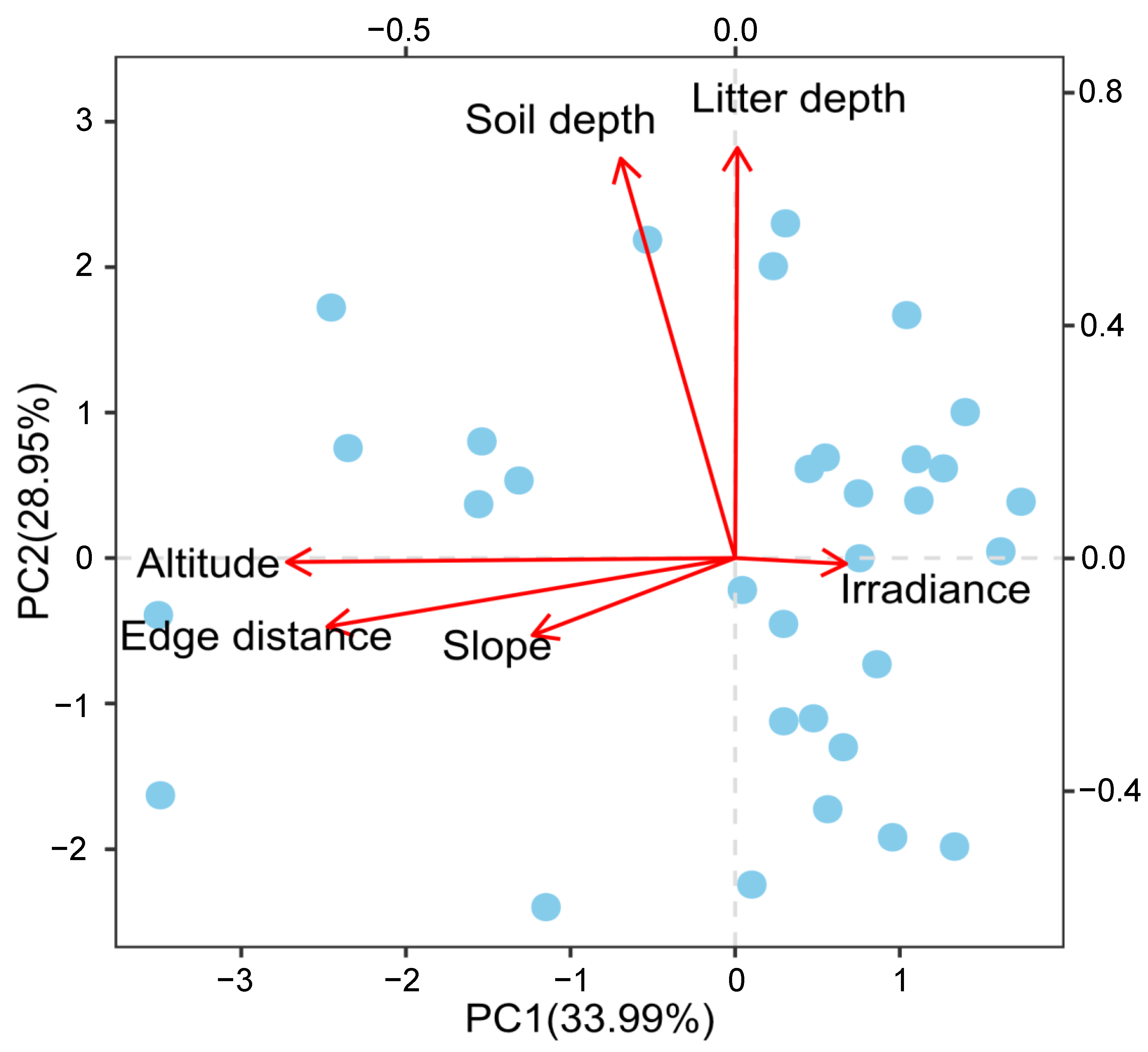
3.3. The Effect of Abiotic and Biotic Factors on Early Seedling Survival
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Excluding Herbivory | Excluding Root Competition | PC2 | R2adj | F92, 3 | p | |
|---|---|---|---|---|---|---|
| Cinnamomum camphora | 18.012 *** | 0.625 | −1.106 | 0.531 | 36.85 | <0.001 |
| Cyclobalanopsis glauca | 10.194 *** | −0.422 | −0.529 | 0.595 | 47.48 | <0.001 |
| Loropetalum chinense | 13.744 *** | 1.041 | 1.044 | 0.217 | 9.776 | <0.001 |
| Schima superba | 1.775 | 0.841 | −0.634 | 0.014 | 1.453 | 0.233 |
References
- Chase, J.M.; Blowes, S.A.; Knight, T.M.; Gerstner, K.; May, F. Ecosystem decay exacerbates biodiversity loss with habitat loss. Nature 2020, 584, 238–243. [Google Scholar] [CrossRef]
- Taubert, F.; Fischer, R.; Groeneveld, J.; Lehmann, S.; Muller, M.S.; Rodig, E.; Wiegand, T.; Huth, A. Global patterns of tropical forest fragmentation. Nature 2018, 554, 519–522. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef] [Green Version]
- Ewers, R.M.; Andrade, A.; Laurance, S.G.; Camargo, J.L.; Lovejoy, T.E.; Laurance, W.F. Predicted trajectories of tree community change in Amazonian rainforest fragments. Ecography 2017, 40, 26–35. [Google Scholar] [CrossRef]
- Laurance, W.F.; Camargo, J.L.C.; Luizao, R.C.C.; Laurance, S.G.; Pimm, S.L.; Bruna, E.M.; Stouffer, P.C.; Williamson, G.B.; Benitez-Malvido, J.; Vasconcelos, H.L.; et al. The fate of Amazonian forest fragments: A 32-year investigation. Biol. Conserv. 2011, 144, 56–67. [Google Scholar] [CrossRef]
- Sorenson, Q.M.; Damschen, E.I.; Lee, T.M. The mechanisms affecting seedling establishment in restored savanna understories are seasonally dependent. J. Appl. Ecol. 2019, 56, 1140–1151. [Google Scholar] [CrossRef]
- Palma, A.C.; Goosem, M.; Stevenson, P.R.; Laurance, S.G.W. Enhancing plant diversity in secondary forests. Front. Forests Glob Chang. 2020, 3, 571352. [Google Scholar] [CrossRef]
- Burgos, A.; Grez, A.A.; Bustamante, R.O. Seed production, pre-dispersal seed predation and germination of Nothofagus glauca (Nothofagaceae) in a temperate fragmented forest in Chile. Forest Ecol. Manag. 2008, 255, 1226–1233. [Google Scholar] [CrossRef]
- Yates, C.J.; Elliott, C.; Byrne, M.; Coates, D.J.; Fairman, R. Seed production, germinability and seedling growth for a bird-pollinated shrub in fragments of kwongan in south-west Australia. Biol. Conserv. 2007, 136, 306–314. [Google Scholar] [CrossRef]
- Cordeiro, N.J.; Ndangalasi, H.J.; McEntee, J.P.; Howe, H.F. Disperser limitation and recruitment of an endemic African tree in a fragmented landscape. Ecology 2009, 90, 1030–1041. [Google Scholar] [CrossRef]
- Ruiz-Carbayo, H.; Bonal, R.; Pino, J.; Espelta, J.M. Zero-sum landscape effects on acorn predation associated with shifts in granivore insect community in new holm oak (Quercus ilex) forests. Divers. Distrib. 2018, 24, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Krishnadas, M.; Comita, L.S. Edge effects on seedling diversity are mediated by impacts of fungi and insects on seedling recruitment but not survival. Front. Forests Glob. Chang. 2019, 2, 76. [Google Scholar] [CrossRef]
- Horst, J.L.; Venable, D.L. Frequency-dependent seed predation by rodents on Sonoran Desert winter annual plants. Ecology 2018, 99, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Larios, L.; Pearson, D.E.; Maron, J.L.; Brody, A. Incorporating the effects of generalist seed predators into plant community theory. Funct. Ecol. 2017, 31, 1856–1867. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.; Swihart, R.K.; Zhao, Y.; Si, X.; Ding, P. Cascading effects of forested area and isolation on seed dispersal effectiveness of rodents on subtropical islands. J. Ecol. 2019, 107, 1506–1517. [Google Scholar] [CrossRef]
- Young, T.P.; Petersen, D.A.; Clary, J.J. The ecology of restoration: Historical links, emerging issues and unexplored realms. Ecol. Lett. 2005, 8, 662–673. [Google Scholar] [CrossRef]
- Negoita, L.; Fridley, J.D.; Lomolino, M.; Mittelhauser, G.; Craine, J.; Weiher, E. Isolation-driven functional assembly of plant communities on islands. Ecography 2016, 39, 1066–1077. [Google Scholar] [CrossRef]
- Rominger, A.J.; Goodman, K.R.; Lim, J.Y.; Armstrong, E.E.; Becking, L.E.; Bennett, G.M.; Brewer, M.S.; Cotoras, D.D.; Ewing, C.P.; Harte, J.; et al. Community assembly on isolated islands: Macroecology meets evolution. Glob. Ecol. Biogeogr. 2016, 25, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Burns, K.C.; Berg, J.; Bialynicka-Birula, A.; Kratchmer, S.; Shortt, K. Tree diversity on islands: Assembly rules, passive sampling and the theory of island biogeography. J. Biogeogr. 2010, 37, 1876–1883. [Google Scholar] [CrossRef]
- Benítez-Malvido, J.; Lázaro, A.; Ferraz, I.D.K. Effect of distance to edge and edge interaction on seedling regeneration and biotic damage in tropical rainforest fragments: A long-term experiment. J. Ecol. 2018, 106, 2204–2217. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Continuous response functions for quantifying the strength of edge effects. J. Appl. Ecol. 2006, 43, 527–536. [Google Scholar] [CrossRef]
- Fischer, R.; Taubert, F.; Muller, M.S.; Groeneveld, J.; Lehmann, S.; Wiegand, T.; Huth, A. Accelerated forest fragmentation leads to critical increase in tropical forest edge area. Sci. Adv. 2021, 7, eabg7012. [Google Scholar] [CrossRef]
- Liu, J.; Matthews, T.J.; Zhong, L.; Liu, J.; Wu, D.; Yu, M. Environmental filtering underpins the island species—Area relationship in a subtropical anthropogenic archipelago. J. Ecol. 2020, 108, 424–432. [Google Scholar] [CrossRef]
- Huebner, C.D.; Randolph, J.C.; Parker, G.R. Environmental factors affecting understory diversity in second-growth deciduous forests. Am. Midl. Nat. 1995, 134, 155–165. [Google Scholar] [CrossRef]
- Jin, Y.; Russo, S.E.; Yu, M.J. Effects of light and topography on regeneration and coexistence of evergreen and deciduous tree species in a Chinese subtropical forest. J. Ecol. 2018, 106, 1634–1645. [Google Scholar] [CrossRef]
- Krishnadas, M.; Bagchi, R.; Sridhara, S.; Comita, L.S. Weaker plant-enemy interactions decrease tree seedling diversity with edge-effects in a fragmented tropical forest. Nat. Commun. 2018, 9, 4523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Coomes, D.A.; Hu, G.; Liu, J.; Yu, J.; Luo, Y.; Yu, M. Larger fragments have more late-successional species of woody plants than smaller fragments after 50 years of secondary succession. J. Ecol. 2019, 107, 582–594. [Google Scholar] [CrossRef]
- Sugiyama, A.; Peterson, C.J. Edge effects act differentially on multiple early regeneration stages of a shade-tolerant tree Tapirira mexicana. Biotropica 2013, 45, 37–44. [Google Scholar] [CrossRef]
- Kiaer, L.P.; Weisbach, A.N.; Weiner, J. Root and shoot competition: A meta-analysis. J. Ecol. 2013, 101, 1298–1312. [Google Scholar] [CrossRef]
- Zheng, Z.; Bai, W.; Zhang, W.-H. Root trait-mediated belowground competition and community composition of a temperate steppe under nitrogen enrichment. Plant Soil 2019, 437, 341–354. [Google Scholar] [CrossRef]
- Janzen, D.H. Seed predation by animals. Annu. Rev. Ecol. Syst. 1971, 2, 465–492. [Google Scholar] [CrossRef]
- Crawley, M.J. Seed Predators and Plant Population Dynamics. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CABI International: Wallingford, UK, 1992; pp. 157–191. [Google Scholar]
- Godó, L.; Valkó, O.; Borza, S.; Deák, B. A global review on the role of small rodents and lagomorphs (clade Glires) in seed dispersal and plant establishment. Glob. Ecol. Conser. 2022, 33, e01982. [Google Scholar] [CrossRef]
- Terborgh, J.; Lopez, L.; Nuñez, P.; Rao, M.; Shahabuddin, G.; Orihuela, G.; Riveros, M.; Ascanio, R.; Adler, G.H.; Lambert, T.D.; et al. Ecological meltdown in predator-free forest fragments. Science 2001, 294, 1923–1926. [Google Scholar] [CrossRef] [Green Version]
- Terborgh, J.; Feeley, K.; Silman, M.; Nunez, P.; Balukjian, N. Vegetation dynamics of predator-free land-bridge islands. J. Ecol. 2006, 94, 253–263. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, X.; Shi, Y.S.; Li, Y.Y.; Wu, J.; He, F.; Chen, X.Y. Habitat fragmentation changes top-down and bottom-up controls of food webs. Ecology 2020, 101, e03062. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Slik, F.; Coomes, D.A.; Corlett, R.T.; Wang, Y.P.; Wilson, M.; Hu, G.; Ding, P.; Yu, M.J. The distribution of plants and seed dispersers in response to habitat fragmentation in an artificial island archipelago. J. Biogeogr. 2019, 46, 1152–1162. [Google Scholar] [CrossRef]
- Hu, G.; Wilson, M.; Zhou, B.B.; Shang, C.; Yu, M.; Wu, J. Spatiotemporal patterns and ecological consequences of a fragmented landscape created by damming. PeerJ 2021, 9, e11416. [Google Scholar] [CrossRef]
- Liu, J.; Zhong, Y.; Zhong, L.; Wei, B.; Zheng, S.; Xie, Y.; Jin, Y.; Yu, M. The asymmetric relationships of the distribution of conspecific saplings and adults in forest fragments. J. Plant Ecol. 2020, 13, 398–404. [Google Scholar] [CrossRef]
- Liu, J.; Vellend, M.; Wang, Z.; Yu, M. High beta diversity among small islands is due to environmental heterogeneity rather than ecological drift. J. Biogeogr. 2018, 45, 2252–2261. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Bagchi, R.; Philipson, C.D.; Slade, E.M.; Hector, A.; Phillips, S.; Villanueva, J.F.; Lewis, O.T.; Lyal, C.H.C.; Nilus, R.; Madran, A.; et al. Impacts of logging on density-dependent predation of dipterocarp seeds in a South East Asian rainforest. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 3246–3255. [Google Scholar] [CrossRef]
- Kelly, D. The evolutionary ecology of mast seeding. Trends Ecol. Evol. 1994, 9, 465–470. [Google Scholar] [CrossRef]
- Thompson, K. Seeds and seed banks. New Phytol. 1987, 106, 23–34. [Google Scholar] [CrossRef]
- Maron, J.L.; Hajek, K.L.; Hahn, P.G.; Pearson, D.E. Seedling recruitment correlates with seed input across seed sizes: Implications for coexistence. Ecology 2019, 100, e02848. [Google Scholar] [CrossRef] [PubMed]
- Brenes-Arguedas, T.; Roddy, A.B.; Kursar, T.A.; Tjoelker, M. Plant traits in relation to the performance and distribution of woody species in wet and dry tropical forest types in Panama. Funct. Ecol. 2013, 27, 392–402. [Google Scholar] [CrossRef]
- Aguiar, M.R.; Sala, A. Competition and facilitation in the recruitment of seedlings in Patagonian steppe. Funct. Ecol. 1992, 6, 66–70. [Google Scholar] [CrossRef]





| Species | Seed Mass (g) 1 | Shade Tolerance | Dominance 2 |
|---|---|---|---|
| Cinnamomum camphora | 12.75 | Shade intolerant | Rare |
| Cyclobalanopsis glauca | 180.34 | Shade tolerant | Rare |
| Loropetalum chinense | 2.43 | Shade intolerant | Dominant |
| Schima superba | 0.66 | Shade tolerant | Common |
| Excluding Herbivory | Excluding Root Competition | PC1 | R2adj | F3, 92 | p | |
|---|---|---|---|---|---|---|
| C. camphora | 18.012 *** | 0.625 | 0.528 | 0.519 | 35.19 | <0.001 |
| C. glauca | 10.194 *** | −0.422 | 0.363 | 0.589 | 46.46 | <0.001 |
| L. chinense | 13.744 *** | 1.041 | 0.143 | 0.206 | 9.218 | <0.001 |
| S. superba | 1.755 | 0.841 | 0.161 | −0.010 | 0.681 | 0.566 |
| Estimate | Std. Error | t Value | Pr (>|t|) | |
|---|---|---|---|---|
| Cinnamomum camphora | ||||
| (Intercept) | 4.806 | 15.795 | 0.304 | 0.764 |
| Soil depth | −0.627 | 5.867 | −0.107 | 0.916 |
| Litter depth | −4.784 | 6.913 | −0.692 | 0.495 |
| Slope | 4.010 | 2.754 | 1.456 | 0.157 |
| Irradiance | −9.724 | 7.423 | −1.310 | 0.201 |
| Edge distance | −3.510 | 2.020 | −1.738 | 0.094 |
| Island area | 0.259 | 0.528 | 0.491 | 0.628 |
| Excluding herbivory | 17.939 | 1.681 | 10.670 | <0.001 |
| Excluding root competetion | 0.5518 | 1.681 | 0.328 | 0.744 |
| Cyclobalanopsis glauca | ||||
| (Intercept) | 6.063 | 6.548 | 0.926 | 0.357 |
| Soil depth | −0.006 | 2.505 | −0.002 | 0.998 |
| Litter depth | −4.450 | 2.893 | −1.555 | 0.124 |
| Slope | 0.416 | 1.158 | 0.360 | 0.720 |
| Irradiance | −3.013 | 3.109 | −0.969 | 0.335 |
| Edge distance | −0.478 | 0.840 | −0.569 | 0.571 |
| Island area | −0.432 | 0.219 | −1.976 | 0.051 |
| Excluding herbivory | 10.194 | 1.002 | 10.175 | <0.001 |
| Excluding root competetion | −0.422 | 1.002 | −0.421 | 0.675 |
| Loropetalum chinense | ||||
| (Intercept) | −4.915 | 26.364 | −0.186 | 0.854 |
| Soil depth | 9.131 | 9.465 | 0.965 | 0.342 |
| Litter depth | −1.790 | 11.499 | −0.156 | 0.878 |
| Slope | 2.772 | 4.302 | 0.644 | 0.525 |
| Irradiance | −17.321 | 11.995 | −1.444 | 0.161 |
| Edge distance | −4.158 | 3.103 | −1.340 | 0.196 |
| Island area | 0.251 | 1.118 | 0.225 | 0.827 |
| Excluding herbivory | 13.544 | 1.687 | 8.031 | <0.001 |
| Excluding root competetion | 0.841 | 1.687 | 0.499 | 0.620 |
| Schima superba | ||||
| (Intercept) | 19.113 | 11.460 | 1.668 | 0.108 |
| Soil depth | −5.494 | 4.229 | −1.299 | 0.203 |
| Litter depth | 3.684 | 5.010 | 0.735 | 0.469 |
| Slope | −1.144 | 1.939 | −0.590 | 0.560 |
| Irradiance | 3.856 | 5.338 | 0.722 | 0.476 |
| Edge distance | 1.172 | 1.340 | 0.838 | 0.413 |
| Island area | −0.559 | 0.425 | −1.317 | 0.231 |
| Excluding herbivory | 1.687 | 1.015 | 1.662 | 0.101 |
| Excluding root competetion | 0.752 | 1.015 | 0.741 | 0.461 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Li, X.; Wu, C.; Zhong, Y.; Yu, M.; Liu, J. Herbivory Rather than Root Competition and Environmental Factors Determines Plant Establishment in Fragmented Forests. Forests 2022, 13, 767. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050767
Li T, Li X, Wu C, Zhong Y, Yu M, Liu J. Herbivory Rather than Root Competition and Environmental Factors Determines Plant Establishment in Fragmented Forests. Forests. 2022; 13(5):767. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050767
Chicago/Turabian StyleLi, Tianxiang, Xue Li, Chuping Wu, Yuping Zhong, Mingjian Yu, and Jinliang Liu. 2022. "Herbivory Rather than Root Competition and Environmental Factors Determines Plant Establishment in Fragmented Forests" Forests 13, no. 5: 767. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050767