
Natural Regeneration of Scots Pine Requires the Application of Silvicultural Treatments such as Overstorey Density Regulation and Soil Preparation


Abstract
:1. Introduction
- (1)
- The most important density and growth parameters in Scots pine seedlings can be controlled by overstorey tree density or the basal area of pine stands;
- (2)
- Soil scarification increases pine seedling densities by reducing the competition pressure exerted by unfavourable ground vegetation types;
- (3)
- The small-scale patterns of varying densities of pine seedling distribution are strongly affected by the spatial distributions and interaction radii of the overstorey pine trees in combination with the prevailing ground cover species and microsite types.
2. Materials and Methods
2.1. Study Region, Sample Design and Tree Characteristics
2.2. Statistical Analyses
2.2.1. Point Pattern Analysis
2.2.2. Modelling
3. Results
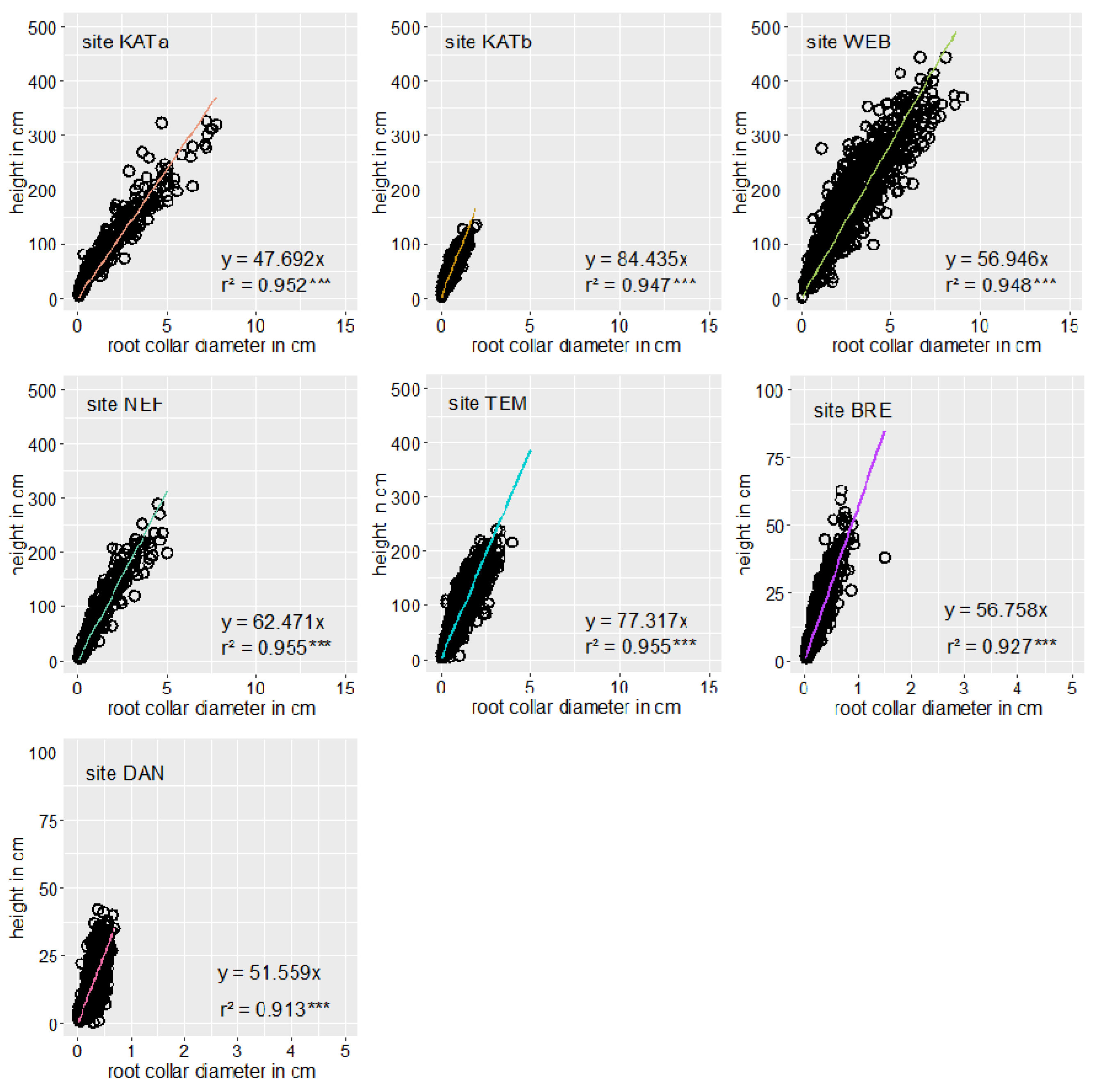
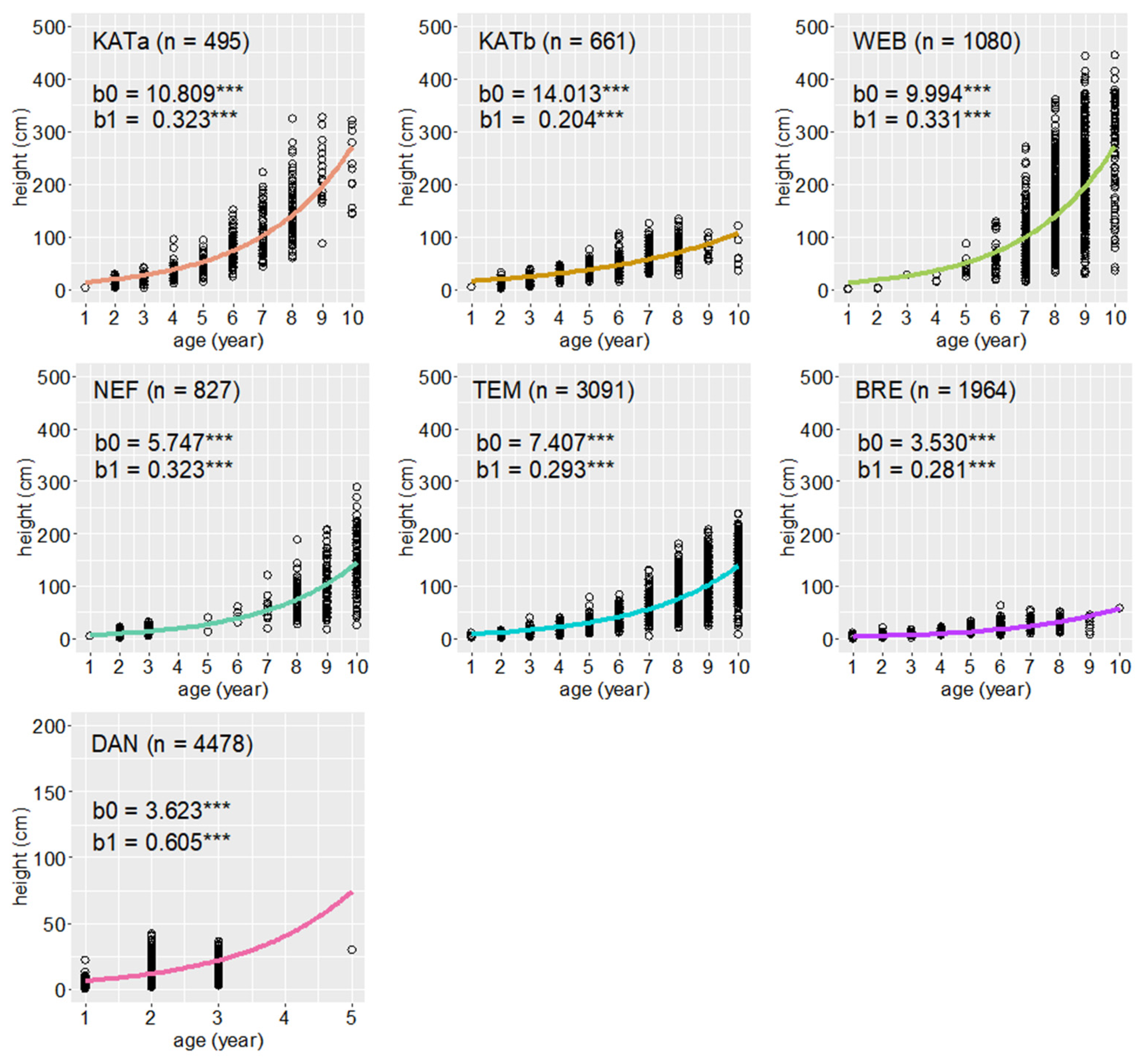
3.1. Densities and Growth Characteristics of Pine Regeneration
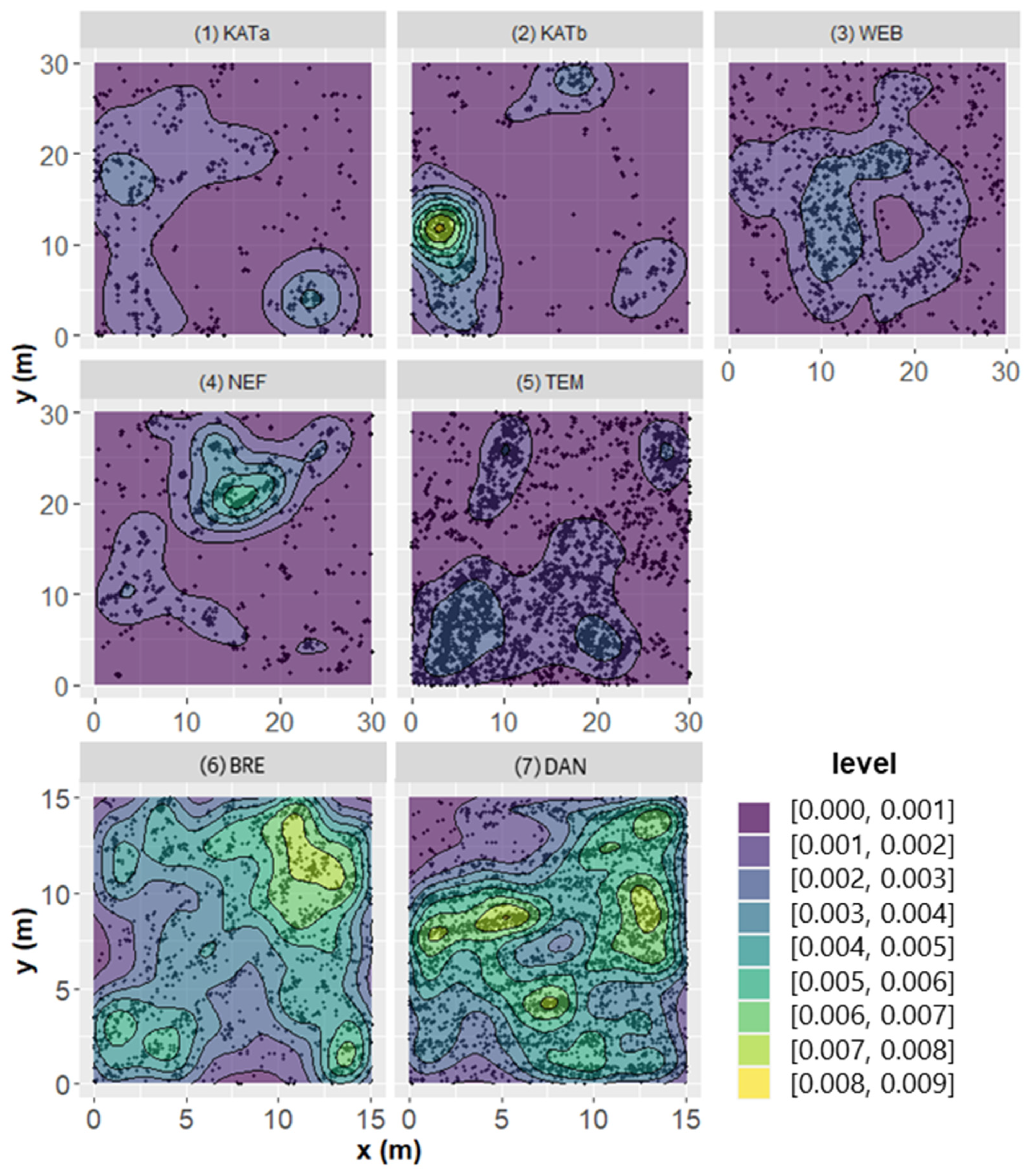
3.2. Density, Spatial Distribution and Growth of Pine Seedlings Relative to Ground Vegetation and Microsite Type
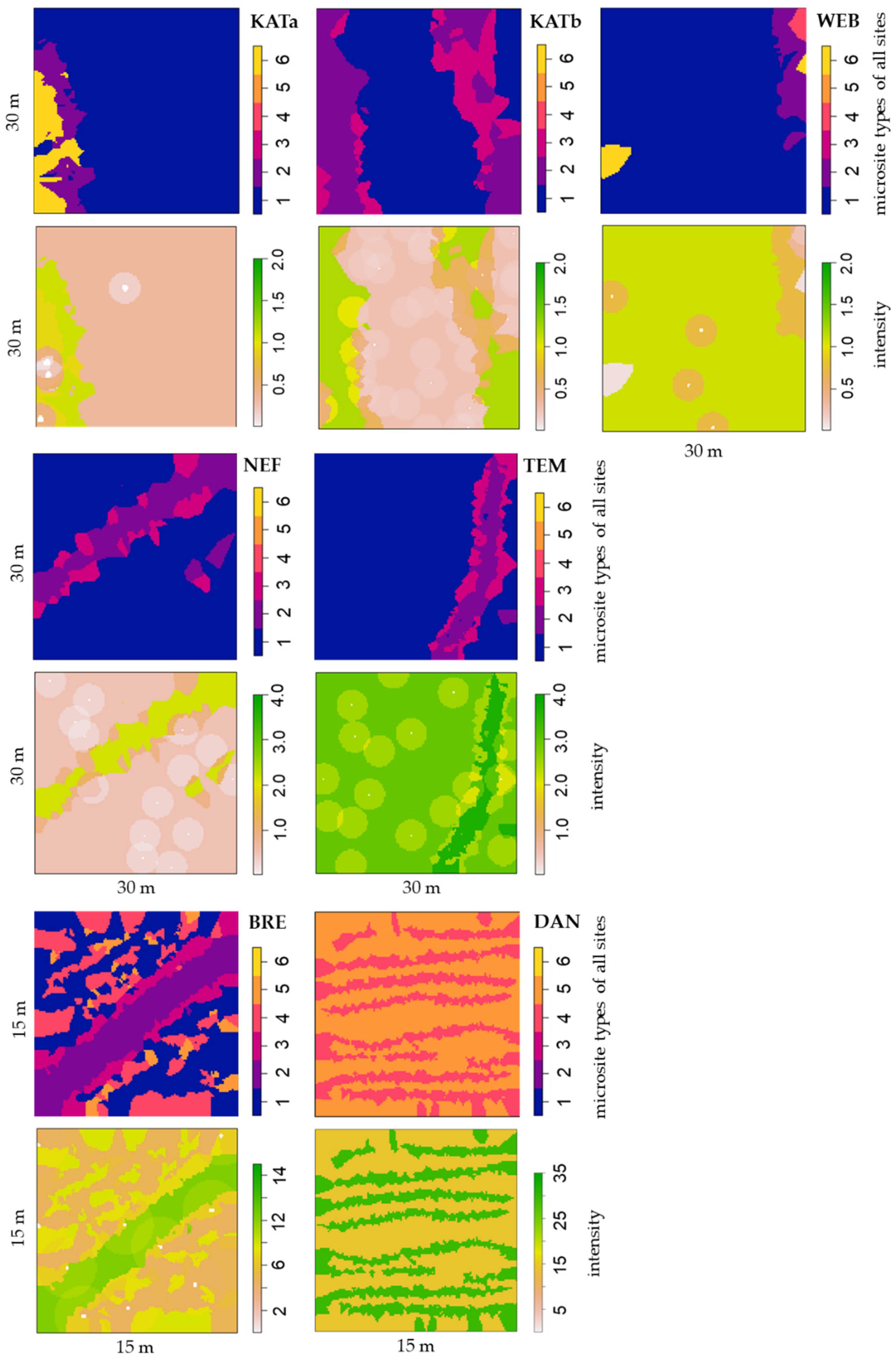
3.3. Effects of Overstorey Trees and Microsite Conditions on Regeneration Density
4. Discussion
4.1. Pine Seedling Density and Growth Affected by Overstorey Trees and Microsites
4.2. Supporting Spatially Heterogenous Pine Seedling Patterns
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spathelf, P.; Ammer, C. Forest management of Scots pine (Pinus sylvestris L.) in northern Germany a brief review of the history and current trends. Forstarchiv 2015, 86, 59–66. [Google Scholar]
- Lust, N.; Geudens, G.; Nachtergale, L. Aspects of biodiversity of Scots pine forests in Europe. Silva Gandav. 2001, 66, 16–39. [Google Scholar] [CrossRef] [Green Version]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.-J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Lundqvist, L.; Ahlström, M.A.; Petter Axelsson, E.; Mörling, T.; Valinger, E. Multi-layered Scots pine forests in boreal Sweden result from mass regeneration and size stratification. For. Ecol. Manag. 2019, 441, 176–181. [Google Scholar] [CrossRef]
- Zawadzka, A.; Słupska, A. Under-Canopy Regeneration of Scots Pine (Pinus sylvestris L.) as Adaptive Potential in Building a Diverse Stand Structure. Sustainability 2022, 14, 1044. [Google Scholar] [CrossRef]
- Miina, J.; Saksa, T. Predicting establishment of tree seedlings for evaluating methods of regeneration for Pinus sylvestris. Scand. J. For. Res. 2008, 23, 12–27. [Google Scholar] [CrossRef]
- Löf, M.; Ammer, C.; Coll, L.; Drössler, L.; Huth, F.; Madsen, P.; Wagner, S. Regeneration Patterns in Mixed-Species Stands; Springer: Cham, Switzerland, 2018; p. 420. [Google Scholar]
- Edwards, C.; Mason, W.L. Stand structure and dynamics of four native Scots pine (Pinus sylvestris L.) woodlands in northern Scotland. For. Int. J. For. Res. 2006, 79, 261–277. [Google Scholar] [CrossRef] [Green Version]
- Vacek, S.; Vacek, Z.; Bílek, L.; Simon, J.; Remeš, J.; Hůnová, I.; Král, J.; Putalová, T.; Mikeska, M. Structure, regeneration and growth of Scots pine (Pinus sylvestris L.) stands with respect to changing climate and environmental pollution. Silva Fenn. 2016, 50, 1564. [Google Scholar] [CrossRef] [Green Version]
- Nyland, R.D. Silviculture: Concepts and Applications, 2nd ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2007; p. 682. [Google Scholar]
- Çoban, S.; Çolak, A.H.; Rotherham, I.D.; Özalp, G.; Çalışkan, A. Effects of canopy gap size on the regeneration of Scots pine (Pinus sylvestris L.) in Eskişehir-Çatacık region of Turkey. Austrian J. For. Sci. 2018, 135, 183–212. [Google Scholar]
- Brichta, J.; Bílek, L.; Linda, R.; Vítámvás, J. Does shelterwood regeneration on natural Scots pine sites under changing environmental conditions represent a viable alternative to traditional clear-cut management? Cent. Eur. For. J. 2020, 66, 104–115. [Google Scholar] [CrossRef]
- Zhou, W. Optimal natural regeneration of Scots pine with seed trees. J. Environ. Manag. 1998, 53, 263–271. [Google Scholar] [CrossRef]
- Béland, M.; Agestam, E.; Ekö, P.M.; Gemmel, P.; Nilsson, U. Scarification and Seedfall affects Natural Regeneration of Scots Pine under Two Shelterwood Densities and a Clear-cut in Southern Sweden. Scand. J. For. Res. 2000, 15, 247–255. [Google Scholar] [CrossRef]
- Barbeito, I.; Fortin, M.-J.; Montes, F.; Cañellas, I. Response of Pine Natural Regeneration to Small-Scale Spatial Variation in a Managed Mediterranean Mountain Forest. Appl. Veg. Sci. 2009, 12, 488–503. [Google Scholar] [CrossRef]
- Huth, F.; Wagner, S. Ökosystemleistungen von Dauerwäldern–eine aktuelle Analyse des Waldbaus. Schweiz. Z. Forstwes. 2013, 164, 27–36. [Google Scholar] [CrossRef]
- Ackzell, L. A comparison of planting, sowing and natural regeneration for Pinus sylvestris (L.) in boreal Sweden. For. Ecol. Manag. 1993, 61, 229–245. [Google Scholar] [CrossRef]
- Hyppönen, M.; Alenius, V.; Valkonen, S. Models for the establishment and height development of naturally regenerated Pinus sylvestris in Finnish Lapland. Scand. J. For. Res. 2005, 20, 347–357. [Google Scholar] [CrossRef]
- Lehto, J. Tutkimuksia männyn uudistamisesta Pohjois-Suomessa siemenpuu-ja suojuspuumenetelmällä. Summary: Studies conducted in northern Finland on the regeneration of Scots pine by means of the seed tree and shelterwood method. Commun. Inst. For. Fenn. 1969, 67, 1–140, (In Finnish with English summary). [Google Scholar]
- Kint, V.; Geudens, G.; Mohren, G.M.J.; Lust, N. Silvicultural interpretation of natural vegetation dynamics in ageing Scots pine stands for their conversion into mixed broadleaved stands. For. Ecol. Manag. 2006, 223, 363–370. [Google Scholar] [CrossRef]
- Hallikainen, V.; Hökkä, H.; Hyppönen, M.; Rautio, P.; Valkonen, S. Natural regeneration after gap cutting in Scots pine stands in northern Finland. Scand. J. For. Res. 2019, 34, 115–125. [Google Scholar] [CrossRef]
- Heinsdorf, M. Kiefernnaturverjüngung–ein historischer Abriss. Beitr. Forstwirtsch. Landsch. 1994, 28, 62–65. [Google Scholar]
- Karlsson, C.; Örlander, G. Soil Scarification Shortly before a Rich Seed Fall Improves Seedling Establishment in Seed Tree Stands of Pinus sylvestris. Scand. J. For. Res. 2000, 15, 256–266. [Google Scholar] [CrossRef]
- Hallingbäck, H.R.; Burton, V.; Vizcaíno-Palomar, N.; Trotter, F.; Liziniewicz, M.; Marchi, M.; Berlin, M.; Ray, D.; Benito Garzón, M. Managing Uncertainty in Scots Pine Range-Wide Adaptation under Climate Change. Front. Ecol. Evol. 2021, 9, 724051. [Google Scholar] [CrossRef]
- Muffler, L.; Schmeddes, J.; Weigel, R.; Barbeta, A.; Beil, I.; Bolte, A.; Buhk, C.; Holm, S.; Klein, G.; Klisz, M.; et al. High plasticity in germination and establishment success in the dominant forest tree Fagus sylvatica across Europe. Glob. Ecol. Biogeogr. J. Macroecol. 2021, 30, 1583–1596. [Google Scholar] [CrossRef]
- Szmyt, J. Spatial statistics in ecological analysis: From indices to functions. Silva Fenn. 2014, 48, 1008. [Google Scholar] [CrossRef] [Green Version]
- Wehnert, A.; Wagner, S.; Huth, F. Effects of Pure and Mixed Pine and Oak Forest Stands on Carabid Beetles. Diversity 2021, 13, 127. [Google Scholar] [CrossRef]
- Dovčiak, M.; Reich, P.B.; Frelich, L.E. Seed rain, safe sites, competing vegetation, and soil resources spatially structure white pine regeneration and recruitment. Can. J. For. Res. 2003, 33, 1892–1904. [Google Scholar] [CrossRef]
- Karlsson, C. Effects of Release Cutting and Soil Scarification on Natural Regeneration in Pinus sylvestris Shelterwoods. Ph.D. Thesis, Swedish University of Agricultural Sciences Alnarp, Alnarp, Sweden, 2000; p. 102. [Google Scholar]
- Mukassabi, T.A.; Polwart, A.; Coleshaw, T.; Thomas, P.A. Scots pine seed dynamics on a waterlogged site. Trees 2012, 26, 1305–1315. [Google Scholar] [CrossRef]
- Hannerz, M.; Almqvist, C.; Hörnfeldt, R. Timing of seed dispersal in Pinus sylvestris stands in central Sweden. Silva Fenn. 2002, 36, 518. [Google Scholar] [CrossRef] [Green Version]
- Kellomäki, S.; Hänninen, H.; Kolström, T.; Kotisaari, A.; Pukkala, T. A tentative model for describing the effects of some regenerative processes on the properties of natural seedlings stands. Silva Fenn. 1987, 21, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Suchockas, V. Distribution of Scots Pine (Pinus sylvestris) Naturally Regenerating Seedlings on Abandoned Agricultural Land at Forest Edges. Balt. For. 2001, 7, 79–83. [Google Scholar]
- Wagner, S.; Herrmann, I.; Dempe, S. Spatial optimization for dispersion of remnant trees in seed-tree cuttings and retention-tree stands of Scots pine. Scand. J. For. Res. 2010, 25, 432–445. [Google Scholar] [CrossRef]
- Valkonen, S.; Ruuska, J.; Siipilehto, J. Effect of retained trees on the development of young Scots pine stands in Southern Finnland. For. Ecol. Manag. 2002, 166, 227–243. [Google Scholar] [CrossRef]
- Dohrenbusch, A. Die Natürliche Verjüngung der Kiefer (Pinus silvestris L.) in Nordwestdeutschen Pleistozän; Schriften aus der Forstlichen Fakultät der Universität Göttingen und der Niedersächsischen Forstlichen Versuchsanstalt Bd. 123; J.D. Sauerländer’s Verlag: Frankfurt am Main, Germany, 1997; p. 269. [Google Scholar]
- Sonohat, G.; Balandier, P.; Ruchaud, F. Predicting solar radiation transmittance in the understory of even-aged coniferous stands in temperate forests. Ann. For. Sci. 2004, 61, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Gaudio, N.; Balandier, P.; Perret, S.; Ginisty, C. Growth of understorey Scots pine (Pinus sylvestris L.) saplings in response to light in mixed temperate forest. Forestry 2011, 84, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Kuuluvainen, T.; Linkosalo, T. Estimation of a spatial tree-influence model using iterative optimization. Ecol. Model. 1998, 106, 63–75. [Google Scholar] [CrossRef]
- Valkonen, S. Effect of Retained Scots Pine Trees on Regeneration, Growth, Form, and Yield of Forest Stands. For. Syst. 2000, 9, 121–145. [Google Scholar] [CrossRef]
- Siipilehto, J. Height distributions of Scots pine sapling stands affected by retained tree and edge stand competition. Silva Fenn. 2006, 40, 473–486. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Beck, W.; Bolte, A.; Hofmann, G.; Jenssen, M.; Krakau, U.; Müller, J. Ökologie und Vegetation der Wälder Nordostdeutschlands: Einfluß von Niederschlagsarmut und Erhöhtem Stickstoffeintrag auf Kiefern-, Eichen- und Buchen-Wald- und Forstökosysteme des Nordostdeutschen Tieflandes, 1st ed.; Dr. Kessel: Oberwinter, Germany, 2002; p. 283. [Google Scholar]
- Örlander, G.; Gemmel, P.; Hunt, J.A. Site Preparation: A Swedish Overview; FRDA Report; BC Ministry of Forests: Victoria, BC, Canada, 1990; Volume 105, p. 61.
- Jäärats, A.; Sims, A.; Seemen, H. The Effect of Soil Scarification on Natural Regeneration in Forest Microsites in Estonia. Balt. For. 2012, 18, 133–143. [Google Scholar]
- Oleskog, G.; Sahlén, K. Effects of Seedbed Substrate on Moisture Conditions and Germination of Pinus sylvestris Seeds in a Clearcut. Scand. J. For. Res. 2000, 15, 225–236. [Google Scholar] [CrossRef]
- Hafemann, E.; Stähr, F. Zur Verjüngung der Kiefer. Eberswalder Forstl. Schr. 2007, 32, 414–420. [Google Scholar]
- Posselt, K.; Huth, F.; Wagner, S.; Müller, M. Kulla-Kultivator fördert Kiefern-Naturverjüngung. AFZ-DerWald 2011, 20, 27–32. [Google Scholar]
- Aleksandrowicz-Trzcińska, M.; Drozdowski, S.; Studnicki, M.; Żybura, H. Effects of Site Preparation Methods on the Establishment and Natural-Regeneration Traits of Scots Pines (Pinus sylvestris L.) in Northeastern Poland. Forests 2018, 9, 717. [Google Scholar] [CrossRef] [Green Version]
- Mäkitalo, K. Effect of Site Preparation and Reforestation Method on Survival and Height Growth of Scots Pine. Scand. J. For. Res. 1999, 14, 512–525. [Google Scholar] [CrossRef]
- Nohrstedt, H.-Ö. Effects of soil scarification and previous N fertilisation on pools of inorganic N in soil after clear-felling of a Pinus sylvestris (L.) stand. Silva Fenn. 2000, 34, 625. [Google Scholar] [CrossRef] [Green Version]
- Forest Inventory Brandenburg. 2013. Available online: https://bwi.info/start.aspx (accessed on 21 November 2021).
- Climate Data Center (CDC). Available online: https://cdc.dwd.de/portal (accessed on 8 January 2022).
- MLuR (Ministerium für Landwirtschaft, Umweltschutz und Raumordnung des Landes Brandenburg). Waldbau-Richtlinie 2004 „Grüner Ordner“ der Landesforstverwaltung Brandenburg; vierC digitalprint—Mediafabrik GmbH: Potsdam, Germany, 2004; Available online: https://forst.brandenburg.de/lfb/de/service/publikationen/detail/~01-05-2004-waldbaurichtlinie-gruener-ordner (accessed on 17 March 2022).
- Ammer, C.; Brang, P.; Knoke, T.; Wagner, S. Methoden zur waldbaulichen Untersuchung von Jungwüchsen. Forstarchiv 2004, 75, 83–110. [Google Scholar]
- Hedderich, J.; Sachs, L. Angewandte Statistik-Methodensammlung mit R-; Springer Spektrum: Berlin/Heidelberg, Germany, 2020; p. 1053. [Google Scholar] [CrossRef]
- Ameztegui, A.; Coll, L. Tree dynamics and co-existence in the montane—Sub-alpine ecotone: The role of different light-induced strategies. J. Veg. Sci. 2011, 22, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Clark, P.J.; Evans, F.C. Distance to nearest neighbor as a measure of spatial relationships in populations. Ecology 1954, 35, 445–453. [Google Scholar] [CrossRef]
- Krebs, C.J. Ecological Methodology, 2nd ed.; Addison Wesley Longman Inc.: Menlo Park, CA, USA, 1999; p. 620. [Google Scholar]
- Horvitz, C.C.; Le Corff, J. Spatial scale and dispersion pattern of ant- and bird-dispersed herbs in two tropical lowland rain forests. Vegetatio 1993, 15, 351–362. [Google Scholar] [CrossRef]
- Pommerening, A.; Grabarnik, P. Individual-Based Methods in Forest Ecology and Management; Springer Nature: Cham, Switzerland, 2019; p. 411. [Google Scholar]
- Dale, M.R.T.; Dixon, P.; Fortin, M.-J.; Legendre, P.; Myers, D.E.; Rosenberg, M.S. Conceptual and mathematical relationships among methods for spatial analysis. Ecography 2002, 25, 558–577. [Google Scholar] [CrossRef]
- Illian, J.; Penttinen, A.; Stoyan, H.; Stoyan, D. Statistical Analysis and Modelling of Spatial Point Patterns; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2009; p. 534. [Google Scholar]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; Taylor & Francis Group: London, UK; Chapmann & Hall: New York, NY, USA; CRC Press: Boca Raton, FL, USA, 2014; p. 510. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns. Methodology and Applications with R; Chapman & Hall/CRC Interdisciplinary Statistics Series; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: London, UK; A Chapman & Hall Book: New York, NY, USA, 2016; p. 810. [Google Scholar]
- Pukkala, T.; Kolström, T. A stochastic spatial regeneration model for Pinus sylvestris. Scand. J. For. Res. 1992, 7, 377–385. [Google Scholar] [CrossRef]
- Karlsson, C. Fertilization and release cutting increase seed production and stem diameter growth in Pinus sylvestris seed trees. Scand. J. For. Res. 2006, 21, 317–326. [Google Scholar] [CrossRef]
- Croker, T.C.; Boyer, W.D. Regenerating Longleaf Pine Naturally; U.S. Department of Agriculture Forest Service Research Paper SO-105; US Department of Agriculture, Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1975; 26p.
- Ayari, A.; Zubizarreta-Gerendiain, A.; Tome, M.; Tome, J.; Garchi, S.; Henchi, B. Stand, tree and crown variables affecting cone crop and seed yield of Aleppo pine forests in different bioclimatic regions of Tunisia. For. Syst. 2012, 21, 128–140. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Fernández, D.; Cañellas, I.; Calama, R.; Gordo, J.; Sánchez-González, M. Thinning increases cone production of stone pine (Pinus pinea L.) stands in the Northern Plateau (Spain). Ann. For. Sci. 2013, 70, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Kuuluvainen, T.; Pukkala, T. Effect of Scots pine seed trees on the density of ground vegetation and tree seedlings. Silva Fenn. 1989, 23, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Eisenhauer, D.-R.; Irrgang, S.; Behrens, S.; Ritter, K. Integration der Kiefern-Naturverjüngung in einen ökologisch orientierten Waldbau—Geotope, Bodenvegetation, Bodenbearbeitung, Konkurrenz des Oberbestandes. Eberswalder Forstl. Schr. 2004, 21, 20–30. [Google Scholar]
- Wagner, S. About a spatial optimization technique in seed-tree cuttings to optimize for dispersion of remnant trees. Forst. Holz. 2008, 63, 29–33, (In German with English Abstract). [Google Scholar]
- Rosenvald, R.; Rosenvald, K.; Kaart, T.; Soolmann, E. Effects of stand parameters on conifer regeneration success in pine shelterwood stands in Estonia. Eur. J. For. Res. 2020, 139, 29–40. [Google Scholar] [CrossRef]
- Karlsson, M. Natural Regeneration of Broadleaved Tree Species in Southern Sweden-Effects of Silvicultural Treatments and Seed Dispersal from Surrounding Stands. Ph.D. Thesis, Swedish University of Agricultural Sciences, Alnarp, Sweden, 2001; p. 124. [Google Scholar]
- Çömez, A.; Tolunay, D.; Güner, Ş.T. Litterfall and the effects of thinning and seed cutting on carbon input into the soil in Scots pine stands in Turkey. Eur. J. For. Res. 2019, 138, 1–14. [Google Scholar] [CrossRef]
- Kuper, J.H. Sustainable Development of Scots Pine Forest; Wageningen Agricultural University Papers Nr.; Wageningen University and Research: Wageningen, The Netherlands, 1994; Volume 94-2. [Google Scholar]
- Hille, M.; den Ouden, J. Improved recruitment and early growth of Scots pine (Pinus sylvestris L.) seedlings after fire and soil scarification. Eur. J. For. Res. 2004, 123, 213–218. [Google Scholar] [CrossRef]
- Köhler, F. Einflussfaktoren auf das Ankommen von Kiefern-Naturverjüngung–vorläufige Ergebnisse von Untersuchungen im Land Brandenburg. Eberswalder Forstl. Schr. 2004, 21, 48–58. [Google Scholar]
- Castro, J.; Leverkus, A.B. Effect of Herbaceous Layer Interference on the Post-Fire Regeneration of a Serotinous Pine (Pinus pinaster Aiton) across Two Seedling Ages. Forests 2019, 10, 74. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK; New York, NY, USA; San Francisco, CA, USA, 1977; p. 892. [Google Scholar]
- Lamont, B.B.; Witkowski, E.T.F.; Enright, N.J. Post-Fire Litter Microsites: Safe for Seeds, Unsafe for Seedlings. Ecology 1993, 74, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Schupp, E.W. Seed-Seedling Conflicts, Habitat Choice, and Patterns of Plant Recruitment. Am. J. Bot. 1995, 82, 399–409. [Google Scholar] [CrossRef]
- Yazdani, R.; Lindgren, D. Gene Dispersion after Natural Regeneration under a Widely-Spaced Seed-Tree Stand of Pinus sylvestris (L.). Silvae Genet. 1992, 41, 1–5. [Google Scholar]
- Aspe, P. Dynamique de la Régénération Naturelle chez le pin Sylvestre, Pinus sylvestris L.: Impacts de la Structure des Peuplements et des Flux de Gènes à Différentes Échelles Spatiales sur la Diversité Génétique. Ph.D. Thesis, Université d’Orléans, Orléans, France, 2009. [Google Scholar]
- Vacek, Z.; Vacek, S.; Bílek, L.; Král, J.; Ulbrichová, I.; Simon, J.; Bulušek, D. Impact of applied silvicultural systems on spatial pattern of hornbeam-oak forests. Cent. Eur. For. J. 2018, 64, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Adámek, M.; Hadincová, V.; Wild, J. Long-term effect of wildfires on temperate Pinus sylvestris forests: Vegetation dynamics and ecosystem resilience. For. Ecol. Manag. 2016, 380, 285–295. [Google Scholar] [CrossRef]
- Wiebecke, E. Der Dauerwald in 16 Fragen und Antworten; der Landwirtschaftskammer für die Provinz Pommern: Stettin, Poland, 1920; p. 51. [Google Scholar]
- Weiß, J. Regeneration of Pine on small areas or under shelter. Allg. Forstzeitschr. 1959, 14, 460–464. [Google Scholar]
- Saursaunet, M.; Mathisen, K.M.; Skarpe, C. Effects of Increased Soil Scarification Intensity on Natural Regeneration of Scots Pine Pinus sylvestris L. and Birch Betula spp. L. Forests 2018, 9, 262. [Google Scholar] [CrossRef] [Green Version]
- González-Martínez, S.C.; Bravo, F. Density and population structure of the natural regeneration of Scots pine (Pinus sylvestris L.) in the High Ebro Basin (Northern Spain). Ann. For. Sci. 2001, 58, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Gerelbaatar, S.; Baatarbileg, N.; Batsaikhan, G. Density and growth performance of Scots pine (Pinus sylvestris L.) understory saplings in logged forests of Tujiin Nars National Park in the Western Khentii Mountains, Case study from Northern Mongolia. Glob. Adv. Res. J. Agric. Sci. 2015, 4, 17–25. [Google Scholar]
- Coban, S.; Colak, A.H.; Rotherham, I.D. Interactions between canopy cover density and regeneration cores of older saplings in Scots pine (Pinus sylvestris L.) stands. For. Syst. 2016, 25, e073. [Google Scholar] [CrossRef] [Green Version]
- Socha, J.; Tymińska-Czabańska, L.; Bronisz, K.; Zięba, S.; Hawryło, P. Regional height growth models for Scots pine in Poland. Sci. Rep. 2021, 11, 10330. [Google Scholar] [CrossRef] [PubMed]
- Lembcke, G.; Knapp, E.; Dittmar, O. Ertragstafel für die Kiefer (Pinus sylvestris L.) im Nordostdeutschen Tiefland; Vieth Verlag GmbH: Eberswalde, Germany, 2000; p. 107. [Google Scholar]
- Ara, M.; Barbeito, I.; Kalén, C.; Nilsson, U. Regeneration failure of Scots pine changes the species composition of young forests. Scand. J. For. Res. 2021, 37, 14–22. [Google Scholar] [CrossRef]
- Behrens, L. Past and Present Scots Pine (Pinus sylvestris L.) Regeneration along Site Type Gradients in Bialowieza Forest, Poland. Ph.D. Thesis, Swedish University of Agricultural Sciences, SLU, Southern Swedish Forest Research Centre, Alnarp, Sweden, 2011. [Google Scholar]
- Eriksson, A.-M.; Olsson, J.; Jonsson, B.G.; Toivanen, S.; Edman, M. Effects of restoration fire on dead wood heterogeneity and availability in three Pinus sylvestris forests in Sweden. Silva Fenn. 2013, 47, 954. [Google Scholar] [CrossRef] [Green Version]
- Aleksandrowicz-Trzcińska, M.; Drozdowski, S.; Brzeziecki, B.; Rutkowska, P.; Jabłońska, B. Effects of different methods of site preparation on natural regeneration of Pinus sylvestris in Eastern Poland. Dendrobiology 2014, 71, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Kuuluvainen, T.; Juntunen, P. Seedling Establishment in Relation to Microhabitat Variation in a Windthrow Gap in a Boreal Pinus sylvestris Forest. J. Veg. Sci. 1998, 9, 551–562. [Google Scholar] [CrossRef]
- Juntunen, V.; Neuvonen, S. Natural regeneration of Scots pine and Norway spruce close to the timberline in northern Finland. Silva Fenn. 2006, 40, 329. [Google Scholar] [CrossRef] [Green Version]
- Booth, M.G.; Hoeksema, J.D. Mycorrhizal networks counteract competitive effects of canopy trees on seedling survival. Ecology 2010, 91, 2294–2302. [Google Scholar] [CrossRef]
- Brockway, D.G.; Outcalt, K.W. Gap-phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manag. 1998, 106, 125–139. [Google Scholar] [CrossRef]
- Niemistö, P.; Lappalainen, E.; Isomäki, A. Growth of Scots pine seed bearers and the development of seedlings during a protracted regeneration period. Folia For. 1993, 826, 1–26. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | KATa | KATb | WEB | NEF | TEM | BRE | DAN |
|---|---|---|---|---|---|---|---|
| stand area (ha) | 4.57 | 8.41 | 1.89 | 6.39 | 3.08 | 3.77 | 13.8 |
| number of trees (n/ha) | 44 | 233 | 44 | 144 | 156 | 311 | 89 |
| age (years) | 120 | 120 | 120 | 125 | 139 | 128 | 106 |
| basal area (m2/ha) | 3.9 | 21.1 | 4.1 | 17.5 | 16.9 | 22.3 | 12.9 |
| mean height (m) | 20.5 (0.047) | 20.2 (0.712) | 22.1 (0.108) | 24.9 (0.056) | 24.8 (0.067) | 22.0 (0.096) | 28.0 (0.110) |
| CV of height | |||||||
| mean dbh (cm) | 36.0 (0.124) | 33.3 (0.874) | 41.4 (0.285) | 38.9 (0.123) | 38.2 (0.168) | 30.0 (0.197) | 46.0 (0.226) |
| CV of dbh | |||||||
| mean crown length (m) | 8.6 (0.184) | 5.2 (0.697) | 8.3 (0.329) | 8.6 (0.317) | 13.4 (0.431) | 7.8 (0.221) | 11.0 (0.282) |
| CV of crown length | |||||||
| mean crown diameter (m) | 5.1 (0.242) | 3.3 (0.635) | 2.9 (0.338) | 5.1 (0.218) | 6.0 (0.204) | 3.5 (0.318) | 6.6 (0.332) |
| CV of crown diameter | |||||||
| RCE | 1.03 | 0.89 | 0.77 | 1.03 | 1.08 | 1.20 | 1.14 |
| Parameters | KATa | KATb | WEB | NEF | TEM | BRE | DAN |
|---|---|---|---|---|---|---|---|
| total seedling density (n/ha) | 5900 | 7844 | 12,089 | 10,589 | 35,856 | 87,511 | 199,067 |
| proportion of pine seedlings with an age up to 10 years (%) | 93.2 | 93.6 | 99.3 | 86.8 | 96.6 | 100.0 | 100.0 |
| m1 seedling density (n/ha) | 4222 | 1967 | 11,489 | 5789 | 29,244 | 24,000 | - |
| m2 seedling density (n/ha) | 1033 | 4500 | 533 | 3967 | 4300 | 27,378 | - |
| m3 seedling density (n/ha) | - | 1378 | 33 | 833 | 2300 | 9911 | - |
| m4 seedling density (n/ha) | - | - | - | - | - | 22,622 | 105,422 |
| m5 seedling density (n/ha) | - | - | - | - | - | 3600 | 93,644 |
| m6 seedling density (n/ha) | 644 | - | 33 | - | - | - | - |
| Parameters | KATa | KATb | WEB | NEF | TEM | BRE | DAN |
|---|---|---|---|---|---|---|---|
| mean age (years) | 6.14 (0.443) | 6.85 (0.983) | 8.16 (0.178) | 5.39 (0.714) | 7.32 (0.347) | 2.02 (0.983) | 1.51 (0.349) |
| CV of age | |||||||
| mean height (cm) | 97.40 (0.925) | 59.59 (1.321) | 158.16 (0.582) | 63.72 (1.206) | 77.25 (0.668) | 7.44 (1.177) | 10.37 (0.761) |
| CV of height | |||||||
| mean rcd (cm) | 1.892 (1.134) | 0.705 (1.553) | 2.575 (0.695) | 1.000 (1.327) | 0.97 (0.726) | 0.13 (1.088) | 0.20 (0.847) |
| CV of rcd | |||||||
| mean crown length (cm) | 85.13 (0.994) | 37.76 (1.186) | 115.48 (0.703) | 73.84 (0.923) | 52.18 (0.718) | 14.19 (0.639) | 13.38 (0.498) |
| CV of crown length | |||||||
| mean crown diameter (cm) | 55.891 (0.987) | 26.34 (1.210) | 75.79 (0.624) | 53.92 (0.849) | 41.21 (0.659) | 10.18 (0.710) | 5.95 (0.476) |
| CV of crown diameter | |||||||
| mean slenderness | 7.07 (0.444) | 10.47 (0.361) | 6.83 (0.353) | 7.51 (0.348) | 8.20 (0.300) | 5.64 (0.341) | 6.21 (0.558) |
| CV of slenderness | |||||||
| mean live crown ratio | 64.44 (0.413) | 49.60 (0.488) | 67.38 (0.285) | 39.50 (0.879) | 54.22 (0.668) | 54.86 (0.348) | 37.29 (1.100) |
| CV of live crown ratio | |||||||
| mean crown profile | 2.30 (1.588) | 2.63 (2.007) | 1.58 (0.524) | 1.65 (0.983) | 1.40 (0.438) | 2.01 (1.069) | 2.69 (0.632) |
| CV of crown profile |
| BA~Height | BA~rcd | BA~Crown Length | BA~Crown Diameter | BA~Slenderness | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Age | r | p-Value | r | p-Value | r | p-Value | r | p-Value | r | p-Value |
| 1 | −0.582 | 0.171 | −0.585 | 0.167 | NA | NA | NA | NA | 0.386 | 0.392 |
| 2 | −0.607 | 0.202 | −0.052 | 0.912 | −0.150 | 0.810 | −0.953 | 0.196 | 0.269 | 0.559 |
| 3 | −0.707 | 0.075 | −0.039 | 0.934 | −0.465 | 0.353 | −0.125 | 0.875 | −0.083 | 0.876 |
| 4 | −0.443 | 0.455 | −0.449 | 0.448 | −0.329 | 0.589 | −0.357 | 0.555 | −0.258 | 0.674 |
| 5 | −0.833 | 0.039 | −0.920 | 0.009 | −0.895 | 0.016 | −0.947 | 0.004 | 0.172 | 0.712 |
| 6 | −0.869 | 0.025 | −0.916 | 0.010 | −0.863 | 0.027 | −0.887 | 0.018 | 0.416 | 0.412 |
| 7 | −0.904 | 0.013 | −0.901 | 0.014 | −0.877 | 0.022 | −0.884 | 0.020 | 0.342 | 0.507 |
| 8 | −0.969 | 0.001 | −0.954 | 0.003 | −0.955 | 0.003 | −0.949 | 0.004 | 0.504 | 0.308 |
| 9 | −0.980 | 0.001 | −0.970 | 0.001 | −0.971 | 0.001 | −0.957 | 0.003 | 0.547 | 0.261 |
| 10 | −0.975 | 0.001 | −0.994 | 0.000 | −0.991 | 0.000 | −0.979 | 0.001 | 0.872 | 0.023 |
| Microsite Types | Indices | KATa | KATb | WEB | NEF | TEM | BRE | DAN |
|---|---|---|---|---|---|---|---|---|
| m1 | RCE | 0.581 | 0.537 | 0.746 | 0.578 | 0.702 | 0.526 | - |
| TESS | 2.746 | 3.659 | 0.982 | 1.810 | 0.403 | 0.239 | - | |
| m2 | RCE | 0.236 | 0.346 | 0.237 | 0.228 | 0.265 | 0.357 | - |
| TESS | 1.063 | 1.015 | 1.719 | 0.521 | 0.283 | 0.108 | - | |
| m3 | RCE | - | 0.351 | 0.033 | 0.258 | 0.204 | 0.326 | - |
| TESS | - | 1.551 | 12.806 | 0.905 | 0.447 | 0.170 | - | |
| m4 | RCE | - | - | - | - | - | 0.302 | 0.517 |
| TESS | - | - | - | - | - | 0.139 | 0.045 | |
| m5 | RCE | - | - | - | - | - | 0.316 | 0.742 |
| TESS | - | - | - | - | - | 0.191 | 0.101 | |
| m6 | RCE | 0.236 | - | 0.035 | - | - | - | - |
| TESS | 2.270 | - | 9.073 | - | - | - | - | |
| total | RCE | 0.598 | 0.595 | 0.770 | 0.573 | 0.736 | 0.684 | 0.810 |
| TESS | 2.401 | 1.777 | 1.065 | 1.271 | 0.392 | 0.163 | 0.071 |
| Functions | Estimate | Std. Error | t-Value | Adj. r2 | p-Value |
|---|---|---|---|---|---|
| lm(age~tess.area-1) | 1.072 | 0.253 | 4.23 | 0.424 | 0.000 |
| lm(height~tess.area-1) | 13.323 | 3.188 | 4.18 | 0.417 | 0.000 |
| lm(rcd~tess.area-1) | 2.487 | 0.521 | 4.77 | 0.486 | 0.000 |
| lm(crown.diameter~tess.area-1) | 6.811 | 1.553 | 4.39 | 0.442 | 0.000 |
| Function | Estimate | Std. Error | t-Value | Adj. r2 | p-Value |
|---|---|---|---|---|---|
| lm(pine.litter~age-1) | 6.016 | 0.434 | 13.880 | 0.771 | 0.000 |
| lm(moss~age-1) | 7.770 | 0.803 | 9.680 | 0.623 | 0.000 |
| lm(vaccinium.vitis~age-1) | 2.638 | 0.294 | 8.965 | 0.582 | 0.000 |
| lm(vaccinium.myrt~age-1) | 2.901 | 0.267 | 10.850 | 0.672 | 0.000 |
| Site | MS | MH | Overstorey Pine Trees | Pine Tree Regeneration | ||||
|---|---|---|---|---|---|---|---|---|
| Radii (m) | Radii (m) | Freq. | Proportion | Intensity | Freq. | Proportion | Intensity | |
| KATa | 2.5 | 0.50 | 4 | 0.00739 | 0.00394 | 501 | 0.99208 | 0.49342 |
| KATb | 2.5 | 0.11 | 32 | 0.04261 | 0.03167 | 674 | 0.95467 | 0.66694 |
| WEB | 2.5 | 0.26 | 5 | 0.00419 | 0.00499 | 1080 | 0.99539 | 1.07853 |
| NEF | 2.5 | 0.10 | 14 | 0.01665 | 0.01421 | 827 | 0.98335 | 0.83937 |
| TEM | 2.5 | 0.10 | 17 | 0.00543 | 0.01661 | 3113 | 0.99457 | 3.04099 |
| BRE | 2.5 | 0.17 | 10 | 0.00505 | 0.03444 | 1969 | 0.99495 | 6.78124 |
| DAN | - | - | 2 | 0.00044 | 0.00693 | 4579 | 0.99956 | 15.87229 |
| Site | Microsite Type Parameter | Estimate | Std. Error | CI95.lo | CI95.hi | Ztest | Zval |
|---|---|---|---|---|---|---|---|
| KATa | m1 | −1.0416456 | 0.05252133 | −1.1445855 | −0.9387056 | *** | −19.832808 |
| m2 | 0.0863855 | 0.11610683 | −0.2441196 | 0.4168907 | *** | 9.715459 | |
| m6 | −0.0725547 | 0.15514918 | −0.4795814 | 0.3344714 | *** | 6.246188 | |
| markX1xX2 | −0.6660055 | 0.04577624 | −0.7557253 | −0.5762857 | *** | −14.549154 | |
| KATb | m1 | −1.2918200 | 0.07544377 | −1.4396871 | −1.1439530 | *** | −17.122951 |
| m2 | 0.1799492 | 0.09434293 | −0.1528831 | 0.5127250 | *** | 15.600207 | |
| m3 | −0.4782191 | 0.11767162 | −0.8567093 | −0.0997110 | *** | 6.914241 | |
| markX1xX2 | −0.1748097 | 0.00615304 | −0.1868695 | −0.1627500 | *** | −28.410310 | |
| WEB | m1 | 0.0842216 | 0.02998521 | 0.0254516 | 0.1429915 | ** | 2.808770 |
| m2 | −0.3185780 | 0.14891511 | −0.6692162 | 0.0320601 | ** | −2.704894 | |
| m3 | −0.9400014 | 0.55854394 | −2.0934977 | 0.2134942 | n.s. | −1.833738 | |
| m6 | −2.3639542 | 0.57812840 | −3.5558351 | −1.1726635 | *** | −4.234658 | |
| markX1xX2 | −0.3952673 | 0.01691281 | −0.4284158 | −0.3621188 | *** | −1.833738 | |
| NEF | m1 | −0.6336548 | 0.04722066 | −0.7262056 | −0.5411040 | *** | −13.419017 |
| m2 | 0.7599928 | 0.07192198 | 0.5264775 | 0.9935081 | *** | 19.377215 | |
| m3 | −0.1041160 | 0.14754053 | −0.4858410 | 0.2776089 | *** | 3.589107 | |
| markX1xX2 | −0.5588286 | 0.02523363 | −0.5093716 | −0.5093716 | *** | −22.146181 | |
| TEM | m1 | 1.0929196 | 0.01859953 | 1.0564652 | 1.1293740 | *** | 58.760602 |
| m2 | 1.3021970 | 0.05467421 | 1.1585831 | 1.4458109 | *** | 3.827718 | |
| m3 | 0.9125870 | 0.07405674 | 0.7309841 | 1.0941900 | * | −2.435060 | |
| markX1xX2 | −0.1713702 | 0.00277762 | −0.1768142 | −0.1659262 | *** | −61.696738 | |
| BRE | m1 | 1.5765652 | 0.04217726 | 1.4938993 | 1.6592311 | *** | 37.379510 |
| m2 | 2.3397087 | 0.05890207 | 2.1415969 | 2.5378205 | *** | 12.956141 | |
| m3 | 1.9592090 | 0.07960589 | 1.7205184 | 2.1978996 | *** | 4.806727 | |
| m4 | 2.0476753 | 0.06157934 | 1.8443161 | 2.2510345 | *** | 7.650457 | |
| m5 | 1.7668650 | 0.12001474 | 1.4489745 | 2.0847554 | n.s. | 1.585637 | |
| markX1xX2 | −0.0662259 | 0.00246044 | −0.0710482 | −0.0614035 | *** | −26.916310 | |
| DAN | m4 | 3.3911820 | 0.02022783 | 3.3515360 | 3.4308280 | *** | 167.64930 |
| m5 | 2.6463600 | 0.02164218 | 2.6039420 | 2.6887780 | *** | 122.27790 | |
| markX1xX2 | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huth, F.; Wehnert, A.; Wagner, S. Natural Regeneration of Scots Pine Requires the Application of Silvicultural Treatments such as Overstorey Density Regulation and Soil Preparation. Forests 2022, 13, 817. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060817
Huth F, Wehnert A, Wagner S. Natural Regeneration of Scots Pine Requires the Application of Silvicultural Treatments such as Overstorey Density Regulation and Soil Preparation. Forests. 2022; 13(6):817. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060817
Chicago/Turabian StyleHuth, Franka, Alexandra Wehnert, and Sven Wagner. 2022. "Natural Regeneration of Scots Pine Requires the Application of Silvicultural Treatments such as Overstorey Density Regulation and Soil Preparation" Forests 13, no. 6: 817. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060817