
Regulatory Control and the Effects of Condensation Water on Water Migration and Reverse Migration of Halostachys caspica (M.Bieb.) C.A.Mey. in Different Saline Habitats
Abstract
:1. Introduction
2. Materials and Methods
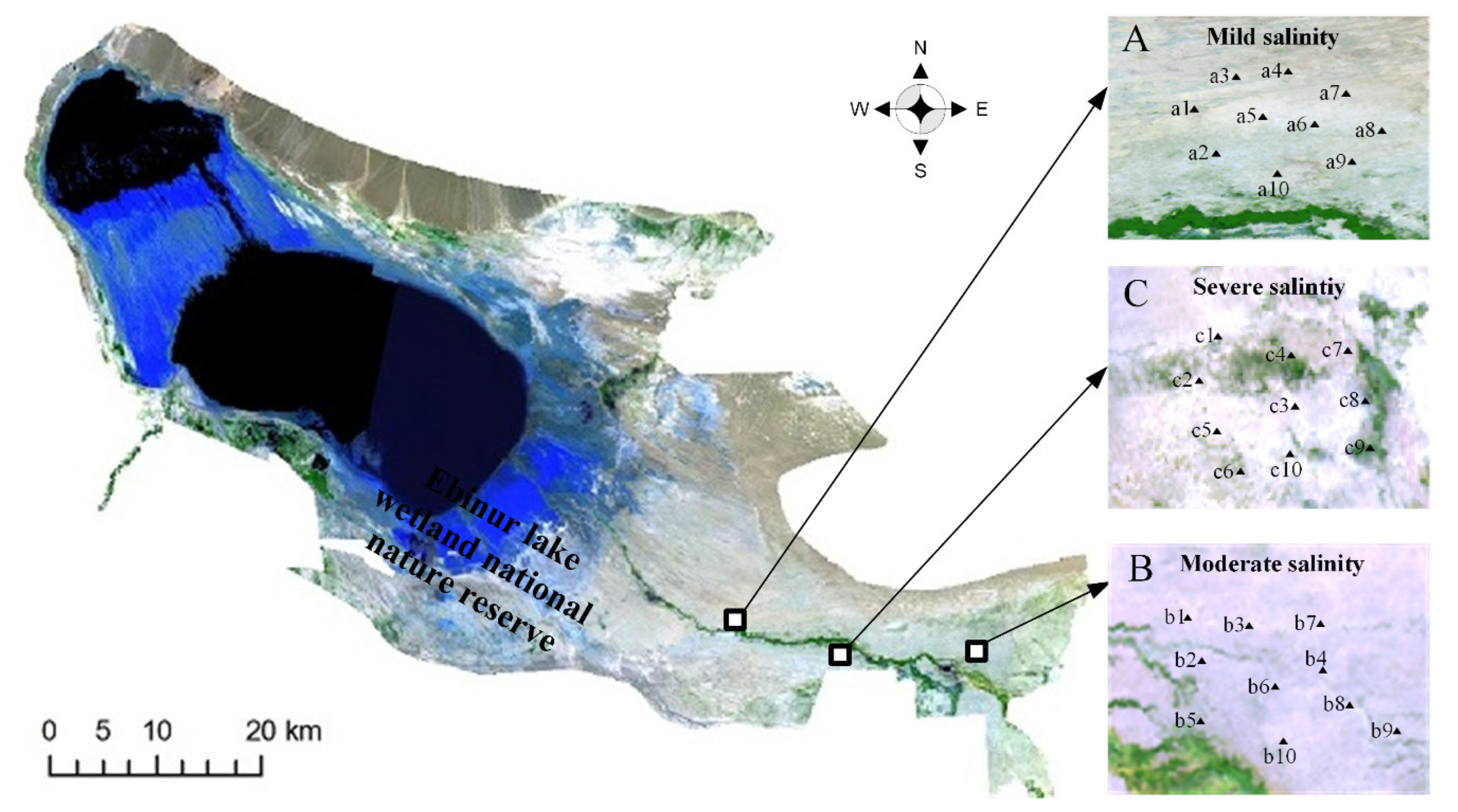
2.1. The Study Region
2.2. Experimental Method and Samples Collection
2.3. Plant Physiological Determination
2.3.1. Presence and Absence Condensation Water Treatment Setting
2.3.2. Water Potentials
2.3.3. Fresh Leaf Water Absorption per Unit Area
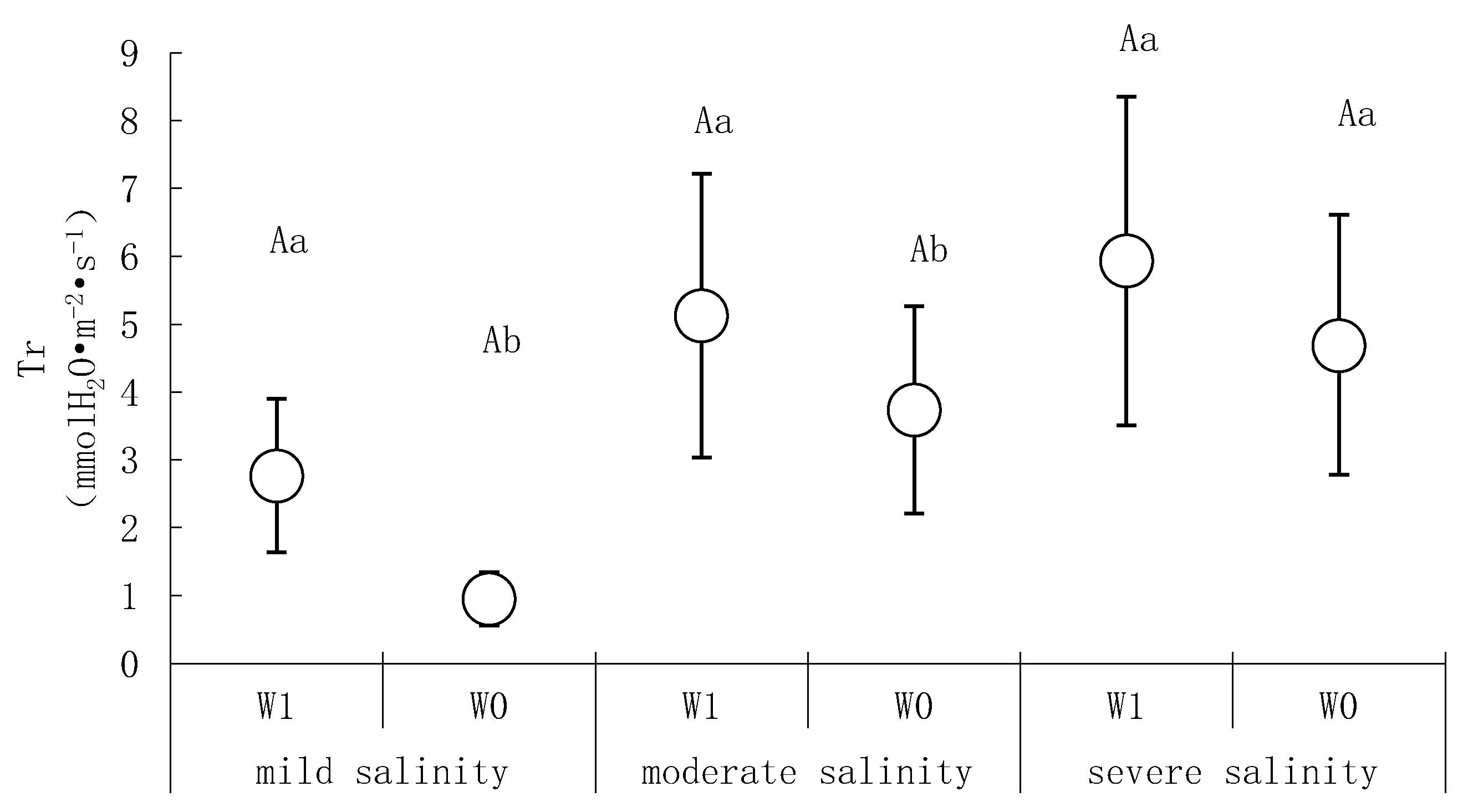
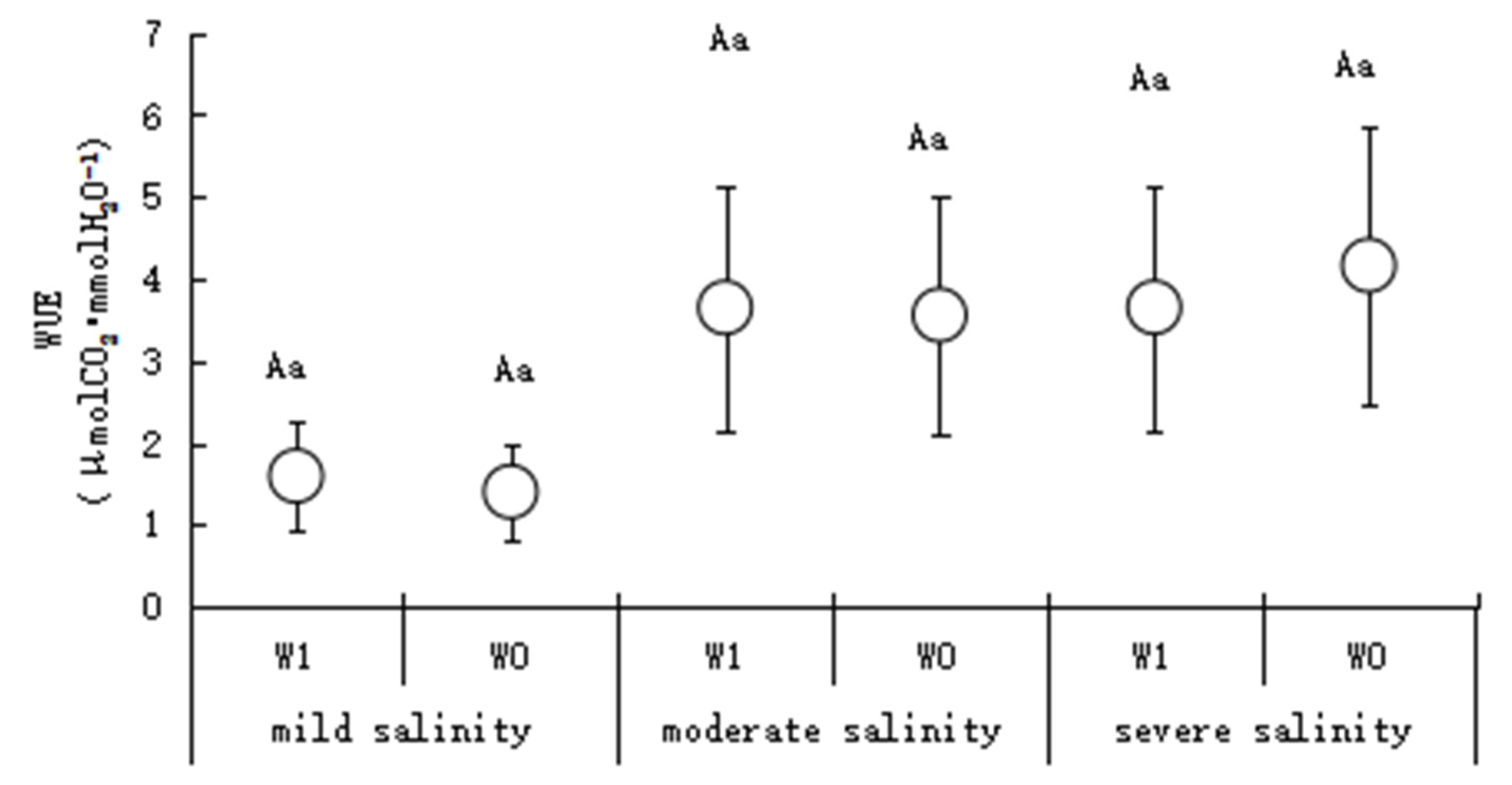
2.3.4. Transpiration Rate (Tr) and Water Use Efficiency (WUE)
2.4. Water Extraction and Isotope Determination
2.5. Data Analysis
3. Result and Analysis
3.1. Plant Physiological Characteristics in Different Salinity Habitats
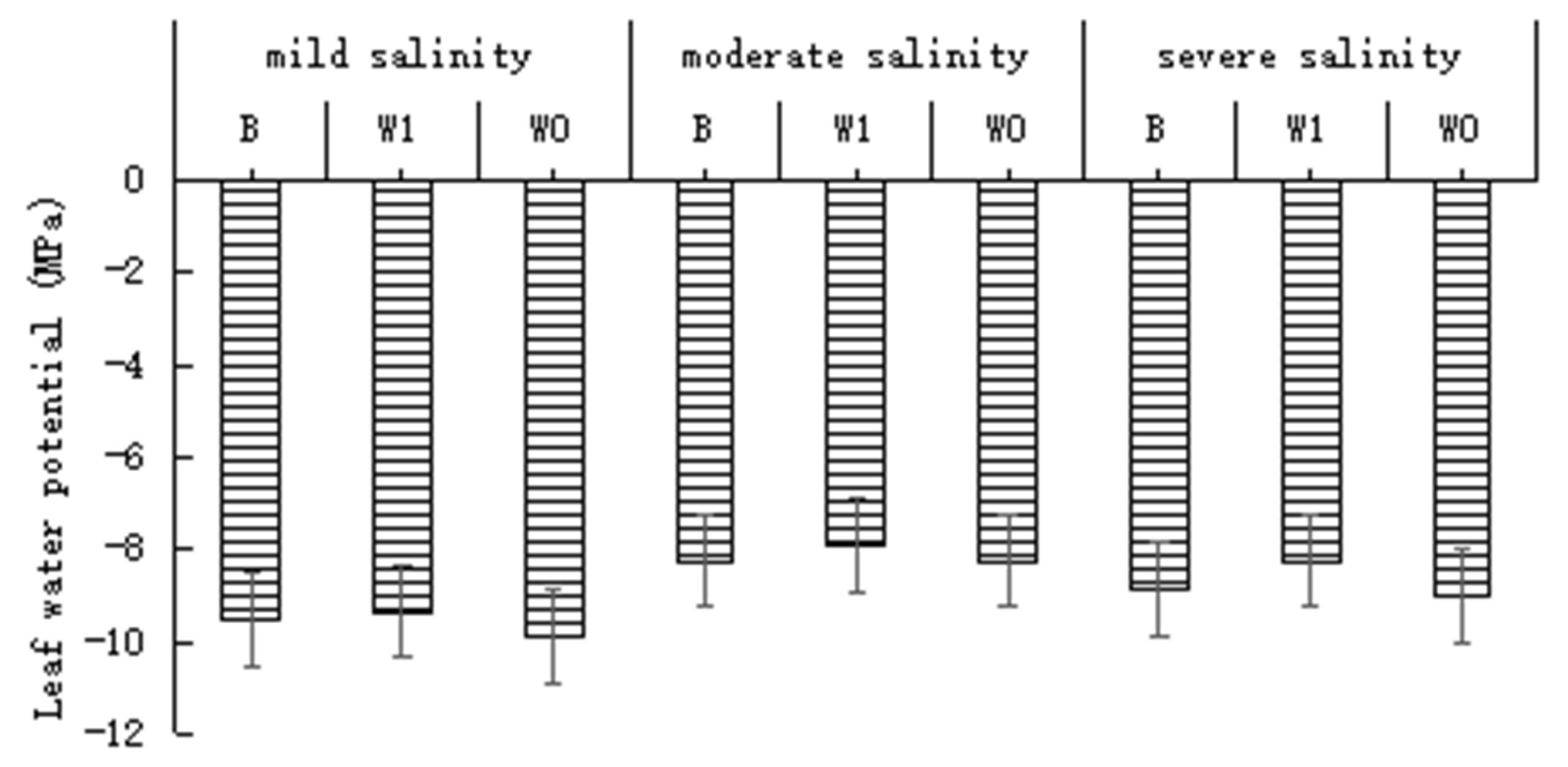
3.1.1. Water Potentials
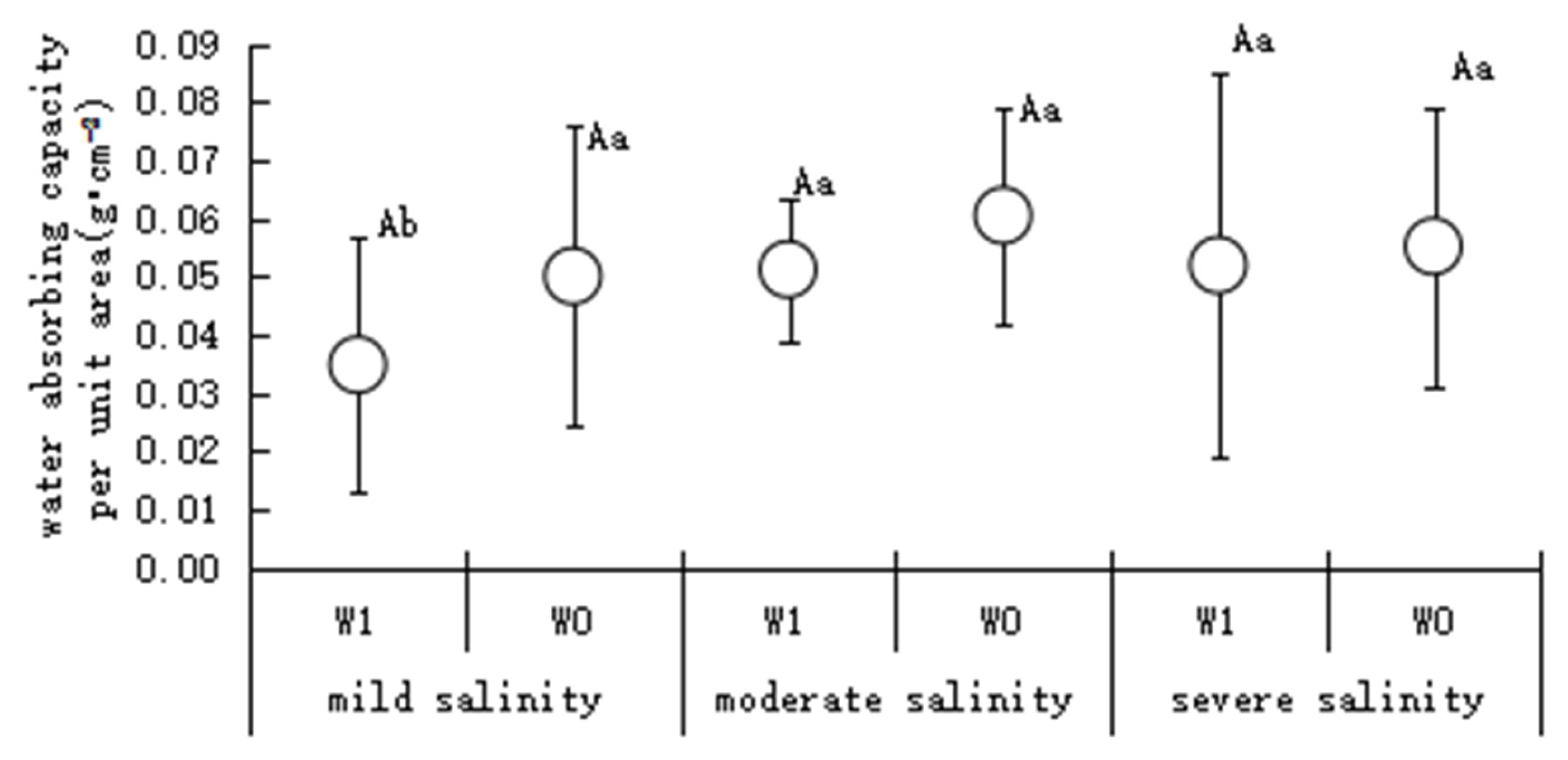
3.1.2. Fresh Leaf Water Absorption per Unit Area
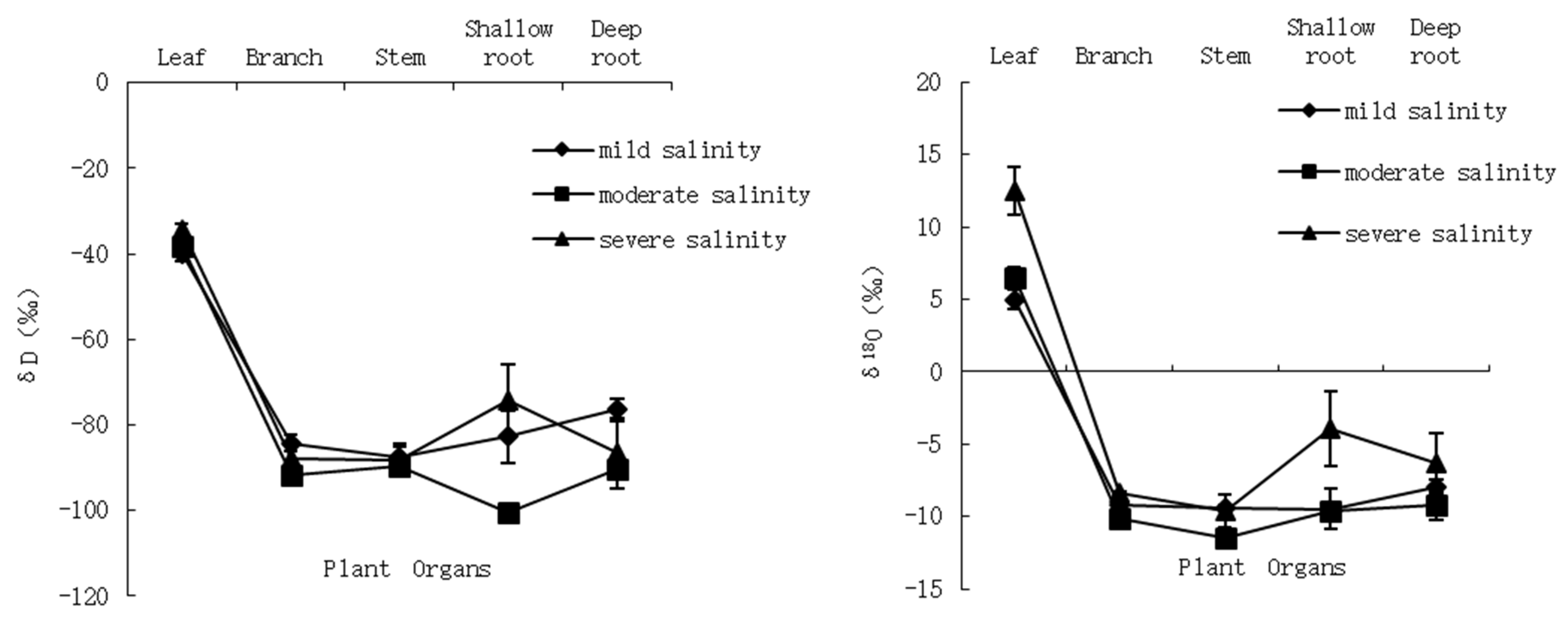
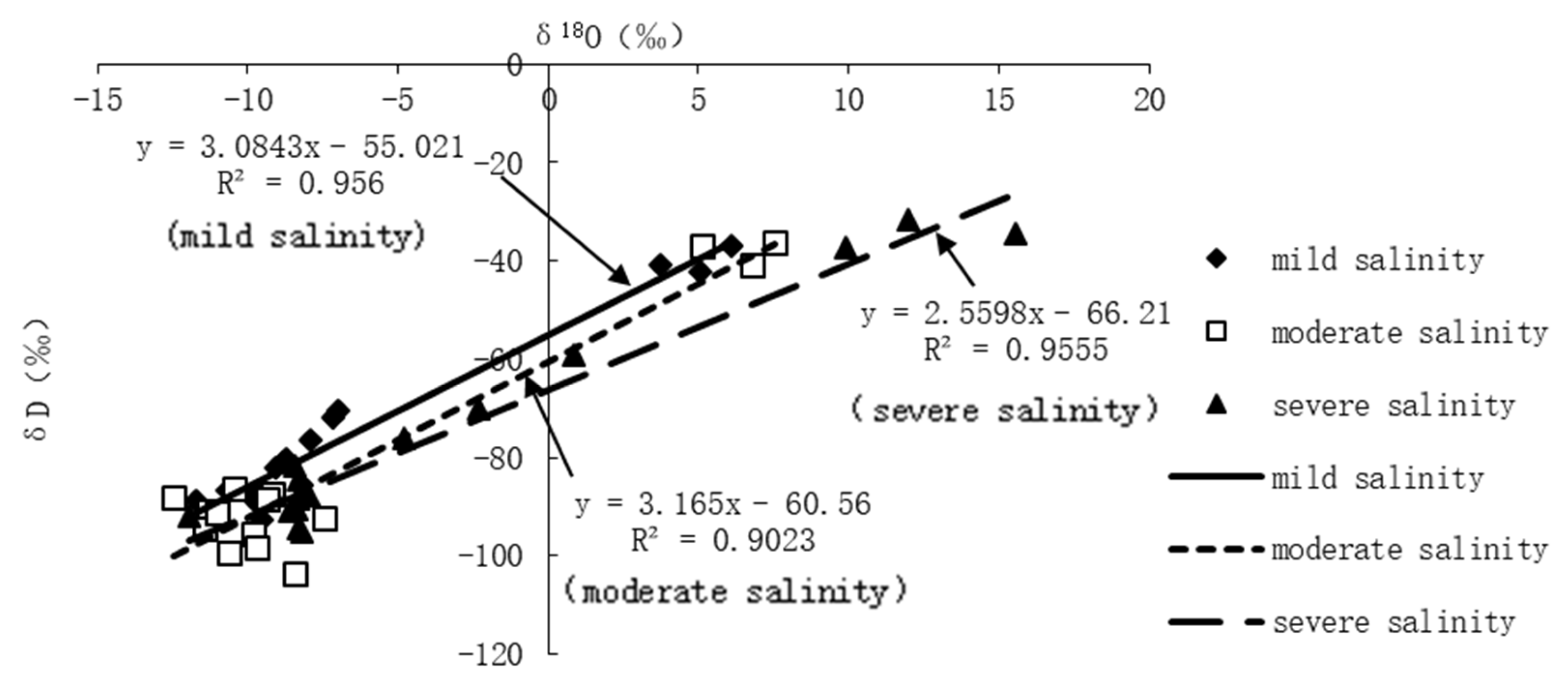
3.1.3. δD and δ18O Composition in Plant Structures and the δD-δ18O Relationship in Different Salinity Habitats
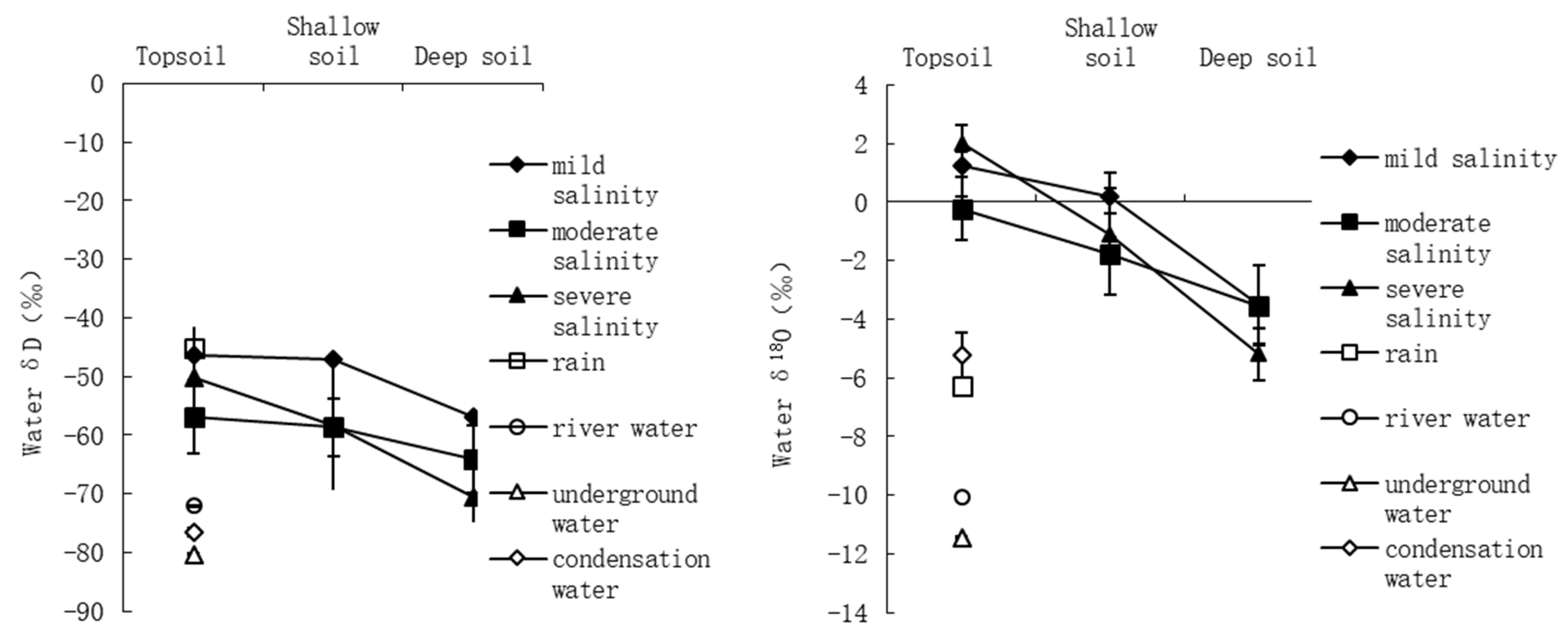
3.2. Hydrogen and Oxygen Isotope Composition and Soil Profiles of Halostachys Caspica Communities
3.3. The Contribution of Condensation Water to Plant Growth
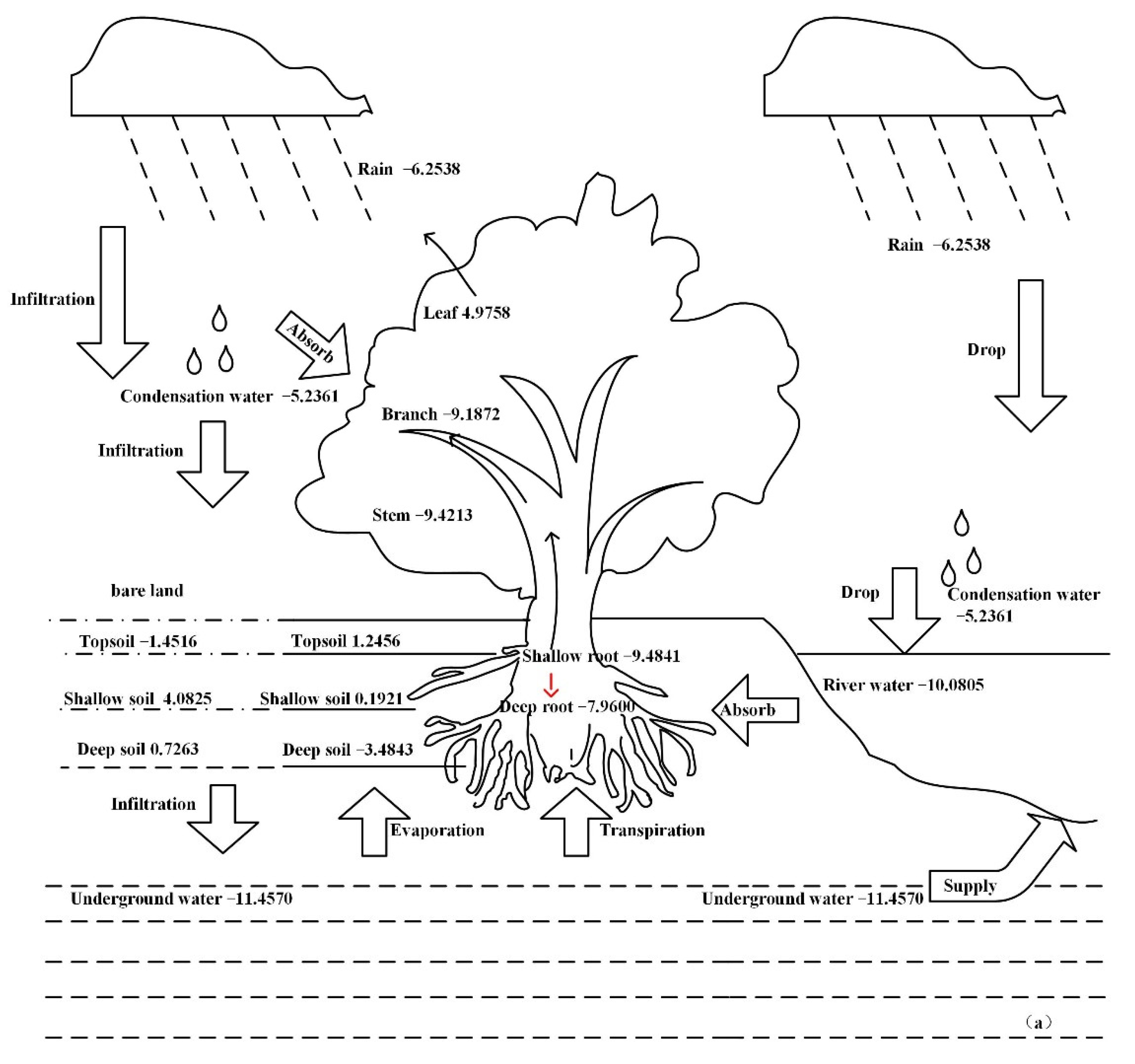
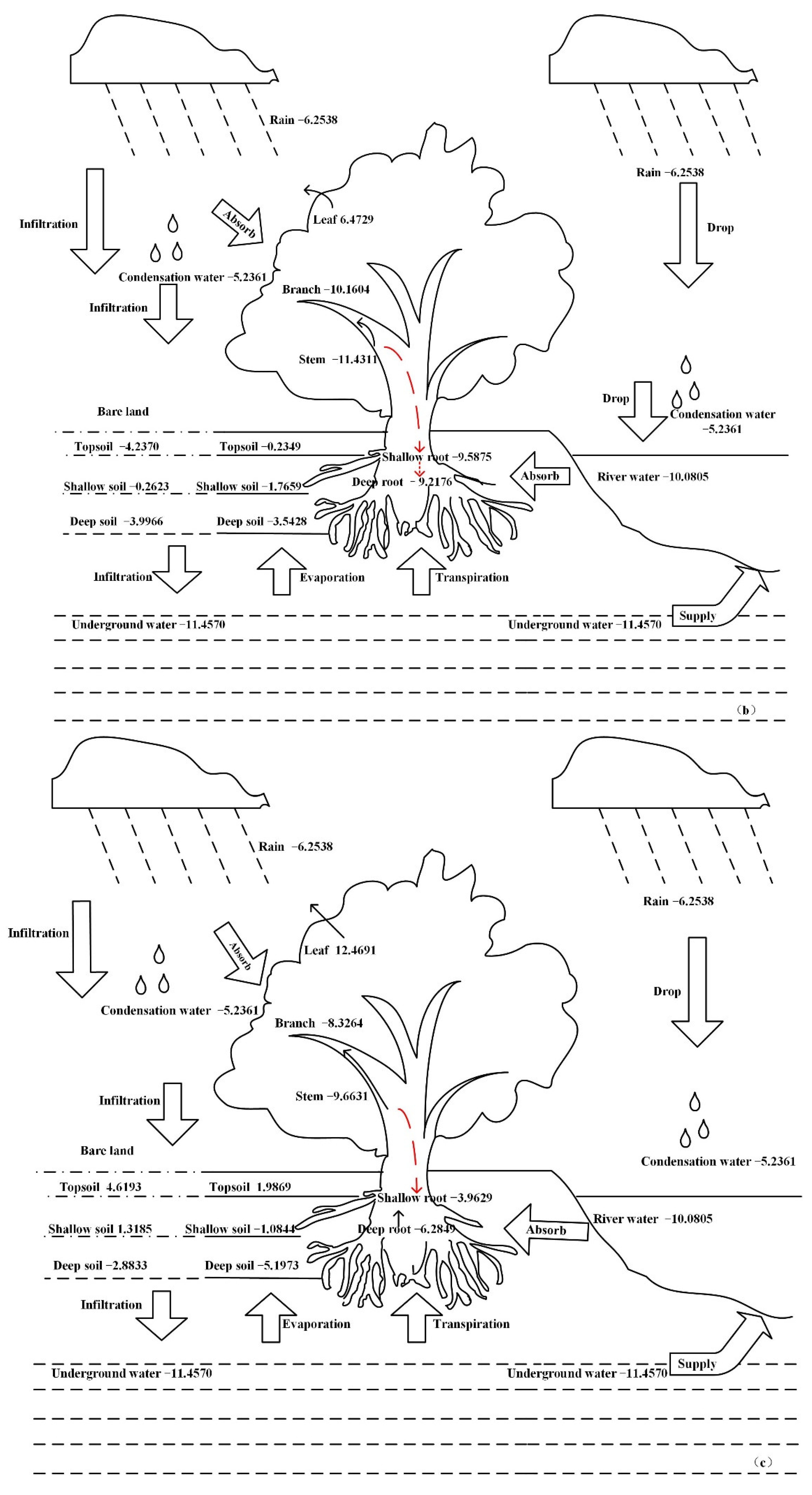
3.4. Water Migration Path in the Plant Body
4. Discussions
4.1. The Contribution of Condensation Water to the Water Needed for the Growth Halostachys Caspica
4.2. The Water Migration Path in Halostachys Caspica
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uclés, O.; Villagarcía, L.; Moro, M.J.; Canton, Y.; Domingo, F. Role of dewfall in the water balance of a semiarid coastal steppe ecosystem. Hydrol. Process. 2014, 28, 2271–2280. [Google Scholar] [CrossRef]
- Fang, J.; Ding, Y.J. An experiental observation of the relationship between sandy soil condensation water and micrometerorogical factors in the arid desert region. J. Desert Res. 2015, 35, 1200–1205. [Google Scholar]
- Pina, A.L.C.B.; Zandavalli, R.B.; Oliveira, R.S.; Martins, F.R. Dew absorption by the leaf trichomes of Combretum leprosum in the Brazilian semiarid region. Funct. Plant Biol. 2016, 43, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Agam, N.; Berliner, P.R. Dew formation and water vapor adsorption in semi-arid environments—A review. J. Arid. Environ. 2006, 65, 572–590. [Google Scholar] [CrossRef]
- Hill, A.J.; Dawson, T.E.; Shelef, O.; Rachmilevitch, S. The role of dew in Negev Desert plants. Oecologia 2015, 178, 317–327. [Google Scholar] [CrossRef]
- Wang, X.H.; Xiao, H.L.; Ren, J.; Cheng, Y.B.; Yang, Q. An ultrasonic humidification fluorescent tracing method for detecting unsaturated atmospheric water absorption by the aerial parts of desert plants. J. Arid. Land 2016, 8, 272–283. [Google Scholar] [CrossRef]
- Kalthoff, N.; Fiebig-Wittmaack, M.; Meiβner, C.; Kohler, M.; Uriarte, M.; Bischoff-Gauß, I.; Gonzales, E. The energy balance, evapo-transpiration and nocturnal dew deposition of an arid valley in the Andes. J. Arid. Environ. 2006, 65, 420–443. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, K.F.; Zhang, W.J. A study on the Cogulation water in the Sandy soil and its role in water balance. Arid. Land Res. 1993, 10, 1–9. [Google Scholar]
- Agam, N. Diurnal Water Content Changes in the Bare Soil of a Coastal Desert. J. Hydrol. 2004, 5, 922–933. [Google Scholar]
- Zhang, Q.; Wang, S.; Yang, F.L.; Yue, P.; Yao, T.; Wang, W.Y. Characteristics of Dew Formation and Distribution, and Its Contribution to the Surface Water Budget in a Semi-arid Region in China. Bound.-Layer Meteorol. 2015, 154, 317–331. [Google Scholar] [CrossRef]
- Maphangwaa, K.W.; Musilb, C.F.; Raittc, L.; Zedda, L. Differential interception and evaporation of fog, dew and water vapor and elemental accumulation by lichens explain their relative abundance in a coastal desert. J. Arid. Environ. 2012, 82, 71–80. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, Y.M. Effects of leaf hair points on dew deposition and rainfall evaporation rates in moss crusts dominated by Syntrichia caninervis, Gurbantunggut Desert, northwestern China. Acta Ecol. Sin. 2012, 32, 7–16. [Google Scholar] [CrossRef]
- Temina, M.; Kidron, G.J. The effect of dew on flint and limestone lichen communities in the Negev Desert. Flora-Morphol. Distrib. Funct. Ecol. Plants 2015, 213, 77–84. [Google Scholar] [CrossRef]
- Cheng, L.; Jia, X.H.; Wu, B.; Li, Y.S.; Zhao, X.B.; Zhou, H. Effects of biological soil crusts on the characteristics of hygroscopic and condensate water deposition in alpine sandy lands. Acta Ecol. Sin. 2018, 38, 5037–5046. [Google Scholar]
- Zhuang, Y.L.; Zhao, W.Z. The ecological role of dew in assisting seed germination of the annual desert plant species in a desert environment, northwestern China. J. Arid. Land 2016, 8, 264–271. [Google Scholar] [CrossRef]
- Arabnejad, H.; Mirzaei, F.; Noory, H. Greenhouse cultivation feasibility using condensation irrigation (studied plant: Basil). Agric. Water Manag. 2021, 245, 106526. [Google Scholar] [CrossRef]
- Goebel, T.S.; Lascano, R.J. Time for cotton to uptake water of a known isotopic signature as measured in leaf petioles. Agric. Sci. 2014, 5, 170–177. [Google Scholar] [CrossRef]
- Gong, X.W. A Probe into the Mechanisms of Canopy Dew Uptake by Photosynthetic Organs of Desert Trees: Based on Molecular, Cellular and Physiological Perspectives; Xin Jiang University: Urumqi, China, 2017. [Google Scholar]
- Liu, J.Q. Characiter of Morphology, anatomy and Physiology of water about extreme xerophytes. Acta Ecol. Sin. 1983, 1, 15–20. [Google Scholar]
- Chen, S.H.J.; Hou, P.; Li, W.H. Comprehensive Scientific Investigation of the Ebinur Lake Wetland Nature Reserve; Xinjiang Science and Technology Press: Urumqi, China, 2006. [Google Scholar]
- Ehleringer, J.R.; Roden, J.; Dawson, T.E. Assessing ecosystemlevel water relations through stable isotope ratio analyses. In Methods in Ecosystem Science; Sala, O.E., Ed.; Springer: New York, NY, USA, 2000; pp. 181–198. [Google Scholar]
- Phillips, D.L.; Gregg, J.W. Source partitioning using stable isotopes: Coping with too many sources. Oecologia 2003, 136, 261–269. [Google Scholar] [CrossRef]
- Qin, L. Formation Mechanism of Condensation Water and Its Ecological Effects of Halostachys caspica Community in Different Salinity Habitats. Ph.D. Thesis, Xinjiang University, Urumqi, China, 2014. [Google Scholar]
- Yang, X.D.; Anwar, E.; Zhou, J.; He, D.; Gao, Y.C.; Lv, G.H.; Cao, Y.E. Higher association and integration among functional traits in small tree than shrub in resisting drought stress in an arid desert. Environ. Exp. Bot. 2022, 201, 1–12. [Google Scholar] [CrossRef]
- Yu, W.; Ji, R.P.; Feng, R.; Zhao, X.L.; Zhang, Y.S. Response of water stress on photosynthetic characteristics and water use efficiency of maize leaves in different growth stage. Acta Ecol. Sin. 2015, 35, 2902–2909. [Google Scholar]
- Jia, G.D. Water Movement Mechanism of Plant-Soil System Using Stable Hydrogen and Oxygen Isotope Technology. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2013. [Google Scholar]
- Yang, X.D. Hydraulic redistribution and mechanism of Populus euphratica and Haloxylon persicum in Ebinur Lake Wetland. Master’s Thesis, Xinjiang University, Urumqi, China, 2011. [Google Scholar]
- Bian, J.J.; Sun, Z.Y.; Zhou, A.G.; Yu, S.W. Advances in the D and 18O isotopes of Water Source of Plants in Arid Areas. Geol. Sci. Technol. Inf. 2009, 28, 117–120. [Google Scholar]
- Ehleringer, J.R.; Dawson, T.E. Water uptake by plants: Perspectives from stable isotopes composition. Plant Cell Environ. 1992, 15, 1073–1082. [Google Scholar] [CrossRef]
- Li, W.H.; Zhou, H.H.; Yang, X.M.; Ding, H. Temporal and special distribution characteristics of aboveground biomass of grassland plant communities in an arid area. Acta Prataculturae Sin. 2010, 19, 186–195. [Google Scholar]
- McCole, A.A.; Stern, L.A. Seasonal water use patterns of Juniperus ashei on the Edwards Plateau, Texas, based on stable isotopes in water. J. Hydrol. 2007, 342, 238–248. [Google Scholar] [CrossRef]
- Liu, J.; Wei, W.; Zhang, L.; Wang, Y.; Duan, B.Q.; Liu, F.L. Application on Isotopes D and 18Oof soil water in water movement of unsaturated Zone. Investig. Sci. Technol. 2012, 5, 38–43. [Google Scholar]
- Zhao, L.J.; Xiao, H.L.; Cheng, G.D.; Song, Y.Y.; Zhao, L.; Li, C.Z.; Yang, Q. A Preliminary Study of Water Sources of Riparian Plants in the Lower Reaches of the Heihe Basin. Acta Geosci. Sin. 2008, 26, 709–718. [Google Scholar]
- Zhu, Q.; Jiang, Z. Using stable isotopes to determine dew formation from atmospheric water vapor in soils in semiarid regions. Arab. J. Geosci. 2016, 9, 1–9. [Google Scholar] [CrossRef]
- Li, H.B. It’s Effects on the Moisture Characteristics of Plant in Semi-Arid Area. Ph.D. Thesis, Inner Mongolia Agricultural University, Hohhot, China, 2010. [Google Scholar]
- Li, X.Y. Effects of gravel and sand mulches on dew deposition in the semiarid region of China. J. Hydrol. 2002, 260, 151–160. [Google Scholar] [CrossRef]
- Ramírez, D.A.; Bellot, J.; Domingo, F.; Blasco, A. Can water responses in Stipa tenacissima L. during the summer season be promoted by nonrainfall water gains in soil? Plant Soil 2007, 291, 67–79. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Zhao, W.Z. Experimental Study of effects of Artifical dew on Bassia dasyphylla and Agriophyllum squarrosum. J. Desert Res. 2010, 30, 1068–1074. [Google Scholar]
- Burgess, S.S.O.; Awson, T.E.D. The contribution of fog to the water relations of Sequoia sempervirens (D.Don): Foliar uptake and prevention of dehydration. Plant Cell Environ. 2004, 27, 1023–1034. [Google Scholar] [CrossRef]
- Ellsworth, P.Z.; Williams, D.G. Hydrogen isotope fraction during water uptake by woody xerophytes. Plant Soil 2007, 291, 93–107. [Google Scholar] [CrossRef]
- Vitarelli, N.C.; Riina, R.; Cassino, M.F.; Meira, R.M.S.A. Trichome-like emergences in Croton, of Brazilian highland rock outcrops: Evidences for atmospheric water uptake. Perspect. Plant Ecol. Evol. Syst. 2016, 22, 23–35. [Google Scholar] [CrossRef]
- Yan, X.; Zhou, M.X.; Dong, X.C.; Zou, S.B.; Xiao, H.L.; Ma, X.F. Molecular mechanisms of foliar water uptake in a desert tree. AoB Plants 2015, 7, 129. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.D.; Lv, G.H.; Ali, A.; Ran, Q.Y.; Gong, X.W.; Wang, F.; Liu, Z.D.; Qin, L.; Liu, W.G. Experimental variations in functional and demographic traits of Lappula semiglabra among dew amount treatments in an arid region. Ecohydrology 2017, 10, e1858. [Google Scholar] [CrossRef]
- Cen, Y.; Liu, M.Z. Effects of dew on eco-physiological traits and leaf structures of Leymus chinensis and Agropyron cristatum grown under drought stress. Chin. J. Plant Ecol. 2017, 41, 1199–1207. [Google Scholar]
- Lin, G.H. Stable Isotope Ecology; Higher Education Press: Beijing, China, 2013; p. 131. [Google Scholar]
- Fang, J. Research progress on the ecological hydrological effect of condensation water. J. Desert Res. 2013, 33, 583–589. [Google Scholar]
- Chen, L.; Yang, X.G.; Song, N.P.; Yang, M.X.; Xiao, X.P.; Wang, X. Leaf water uptake strategy of plants in the arid and semi-arid region of Ningxia. J. Zhengjiang Univ. (Argic. Life Sci.) 2013, 39, 565–574. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Contribution of Water Source (%) | ||||
|---|---|---|---|---|---|
| Soil Water | Rain | River Water | Ground Water | Condensation Water | |
| mild salinity | 11.375 ± 2.071 | 8.525 ± 1.280 | 27.025 ± 0.330 | 46.1 ± 4.594 | 7.05 ± 1.100 |
| moderate salinity | 6.775 ± 2.507 | 4.575 ± 1.661 | 18.5 ± 5.750 | 66.4 ± 11.253 | 3.775 ± 1.375 |
| severe salinity | 27.9 ± 9.484 | 11.925 ± 2.258 | 20.1 ± 3.955 | 28.975 ± 10.150 | 11.125 ± 2.478 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, L.; He, X.; Lv, G.; Yang, J. Regulatory Control and the Effects of Condensation Water on Water Migration and Reverse Migration of Halostachys caspica (M.Bieb.) C.A.Mey. in Different Saline Habitats. Forests 2022, 13, 1442. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091442
Qin L, He X, Lv G, Yang J. Regulatory Control and the Effects of Condensation Water on Water Migration and Reverse Migration of Halostachys caspica (M.Bieb.) C.A.Mey. in Different Saline Habitats. Forests. 2022; 13(9):1442. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091442
Chicago/Turabian StyleQin, Lu, Xuemin He, Guanghui Lv, and Jianjun Yang. 2022. "Regulatory Control and the Effects of Condensation Water on Water Migration and Reverse Migration of Halostachys caspica (M.Bieb.) C.A.Mey. in Different Saline Habitats" Forests 13, no. 9: 1442. https://0-doi-org.brum.beds.ac.uk/10.3390/f13091442